











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La anatomía de la madera es una valiosa fuente de información con respecto a las relaciones jerárquicas de las especies y las condiciones ambientales en las que se establecen (Carlquist, 2012). En México, existen trabajos que describen la anatomía de la madera de plantas que crecen en determinados tipos de vegetación. Entre ellos, se encuentran estudios en selva mediana subperennifolia (Rebollar-Domínguez et al., 1997), selva alta perennifolia (Barajas-Morales et al., 1997), matorral xerófilo (Abundiz-Bonilla et al., 2004; Quintanar-Isaías et al., 2005), bosques templados (Pérez-Olvera y Quintanar-Isaías, 1994) y en mayor número en bosques mesófilos de montaña (Aguilar-Rodríguez y Barajas-Morales, 2000; Aguilar-Rodríguez y Castro-Plata., 2006; Aguilar-Alcántara et al., 2014).
La selva baja caducifolia (SBC) ocupa entre 8 y 14% del territorio nacional y cuenta con una riqueza que fluctúa entre 22 y 97 especies de plantas por 0.1 ha (Leopold, 1950; Miranda y Hernández-Xolocotzi, 1963; Rzedowski, 1978; Trejo y Dirzo, 2000; González, 2003; Méndez-Toribio et al., 2014), por lo tanto, tiene una elevada riqueza florística. El estudio de la anatomía de la madera de especies que se distribuyen en este tipo de vegetación aporta evidencia sobre la diversidad e identidad taxonómica (Trejo y Dirzo, 2000), ya que la SBC se caracteriza por presentar temporadas de sequía prolongadas que propician tiempos de floración muy cortos, lo que dificulta la colecta de estructuras reproductivas (Rzedowski, 1978). Además, se puede investigar la respuesta de los caracteres anatómicos de las maderas a las condiciones abióticas de los sitios de estudio (Carlquist, 2012; Beeckman, 2016; Ramírez-Martínez et al., 2017).
A pesar de lo antes mencionado, solo se registran dos estudios anatómicos publicados de plantas leñosas distribuidas en SBC mexicanas. Barajas-Morales y León-Gómez (1989) describieron la madera de 72 especies de Chamela, Jalisco. Las cuales se caracterizan por presentar porosidad difusa, parénquima axial escaso, vasicéntrico, aliforme y confluente, radios uniseriados a multiseriados y fibras septadas, también tienen cristales, gomas y tílides. Posteriormente, Ramírez-Martínez et al. (2017) estudiaron la anatomía de 21 especies en Tamaulipas; señalando que los caracteres cuantitativos de los elementos de vaso de esta región son diferentes a lo reportado por Barajas-Morales y León-Gómez (1989). Ramírez-Martínez et al. (2017) atribuyen estas diferencias a la relación alométrica entre la altura de la planta y las dimensiones de las células del xilema.
En el estado de Puebla la SBC representa 15.68% del territorio estatal, su extensión es superada solamente por las regiones destinadas a actividades agrícolas y forestales, ocupa casi por completo la Mixteca Alta y las regiones que limitan con los estados de Morelos, Guerrero y Oaxaca (Handal-Silva et al., 2011). Por tratarse de una región extensa y diversa, resulta relevante documentar la diversidad anatómica de especies leñosas que ahí ocurren. Los objetivos de este estudio son describir la anatomía de la madera de Berberis moranensis Schult. & Schult. f., Bursera fagaroides (Kunth) Engl., B. morelense Ramírez, Cascabela ovata (Cav.) Lippold, Critoniopsis uniflora (Sch. Bip.) H. Rob., Fouquieria formosa Kunth, Ipomoea murucoides Roem. & Schult., Karwinskia humboldtiana (Schult.) Zucc. y Lycianthes sp., taxones característicos de la Sierra del Tentzo, comparar estadísticamente las maderas; así como identificar si los caracteres son suficientes para diferenciarlas a nivel intra e interespecífico y determinar semejanzas con organismos congéneres estudiados en otras regiones.
Materiales y Métodos
Área de estudio
El material estudiado fue recolectado en el municipio Huehuetlán el Grande, Puebla, México, dentro de la Sierra del Tentzo (Fig. 1). Esta sierra se ubica en la parte central del estado de Puebla, al sur de la presa de Valsequillo, entre los municipios Atlixco, Atoyatempan, Huehuetlán el Grande y Molcaxac; la vegetación se compone de una transición de bosque de pino-encino, matorral y selva baja caducifolia (Pérez-Pérez et al., 2018). La Sierra del Tentzo se encuentra formada por lomeríos con altitudes entre 2000 y 2350 m, con temperaturas medias mensuales que oscilan entre 16 y 21.5 °C; la máxima precipitación mensual no supera los 200 mm, siendo los meses de mayo a septiembre los más lluviosos (Morón et al., 2000).
Trabajo de campo
Se seleccionaron tres individuos adultos de las especies de árboles más comunes en el sitio de estudio; se realizó la colecta de trozos de madera del tronco principal a una distancia de 30 cm del suelo, utilizando un serrucho (Truper®, Estado de México, México). Todas las muestras se fijaron en formaldehído, alcohol, ácido acético glacial y agua en proporción 1:1:1 (FAA; Sass, 1958). Paralelamente, se realizó la medición de la altura de los árboles con un clinómetro (Suunto, Vantaa, Finlandia), el diámetro a la altura del pecho (DAP) con una forcípula (GAFMEX, Cuernavaca, México) y cobertura de los individuos muestreados con una cinta métrica (Truper®, Estado de México, México). La media y desviación estándar se reportan en el Cuadro 1.
Cuadro 1: Media y desviación estándar de altura, Diámetro a la Altura del Pecho (DAP) y cobertura de las especies estudiadas en la Sierra del Tentzo, Puebla, México.
| Especie | Altura (m) | DAP (cm) | Cobertura (m) |
|---|---|---|---|
| Berberis moranensis Schult. & Schult. f. | 2.5 (0.3) | 6.52 (0.8) | 6.52 (1.7) |
| Bursera fagaroides (Kunth) Engl. | 3.7 (1.2) | 7 (0.4) | 7.00 (1.13) |
| Bursera morelense Ramírez | 2.9 (0.2) | 8.59 (1.01) | 8.59 (0.9) |
| Cascabela ovata (Cav.) Lippold | 4.5 (1.1) | 7.32 (0.75) | 7.32 (1.85) |
| Critoniopsis uniflora (Sch. Bip.) H. Rob. | 3.2 (0.5) | 7.63 (0.52) | 7.63 (1.35) |
| Fouquieria formosa Kunth | 4 (0.5) | 6.46 (0.5) | 6.46 (0.25) |
| Ipomoea murucoides Roem. & Schult. | 6.5 (0.8) | 41.38 (3.5) | 41.38 (5.25) |
| Karwinskia humboldtiana (Schult.) Zucc. | 3.2 (0.5) | 5.41 (1.1) | 5.41 (0.7) |
| Lycianthes sp. | 2.7 (0.25) | 6.04 (1.3) | 6.04 (0.5) |
Los individuos fueron identificados en campo utilizando la Flora del Valle de México (Calderón de Rzedowski y Rzedowski, 2005), los fascículos de Burseraceae (Medina-Lemos, 2008) y Fouquieriaceae (Ezcurra y Medina-Lemos, 1997) de la Flora del Valle de Tehuacán-Cuicatlán, y el fascículo de la familia Apocynaceae de la Flora del Bajío y de Regiones Adyacentes (Rzedowski y Calderón de Rzedowski, 1998).
Trabajo de laboratorio
Los trozos de madera fueron cortados en cubos de 3 cm3, se lavaron y ablandaron en una solución de glicerina, alcohol y agua (GAA, 1:1:1; Sass, 1958) durante 20-25 días. Posteriormente se cortaron con un micrótomo de deslizamiento (Leica 2000R, Wetzlar, Alemania), en los planos transversal, tangencial y radial con un grosor de 25-30 μm. Los cortes se deshidrataron en alcoholes graduales desde 50 hasta 100% de concentración, se tiñeron con safranina y verde rápido, mediante la técnica convencional (Sass, 1958); finalmente, se montaron en resina sintética (Johansen, 1940). Se elaboraron laminillas temporales para medir longitud de los elementos de vaso y fibras de cada individuo, las cuales se obtuvieron de material disociado con solución Jeffrey (Berlyn y Miksche, 1976); también se verificó la presencia de traqueidas. Los caracteres cualitativos y cuantitativos se obtuvieron con un analizador de imágenes Image Pro Plus v. 6.1 (Media Cybernetics, Rockville, EUA) adaptado a un microscopio Olympus BX-51 (Olympus Corporation, Tokio, Japón). Las descripciones se realizaron con base en las recomendaciones de la Asociación Internacional de Anatomistas de la Madera (IAWA Committee, 1989). El parénquima radial se clasificó de acuerdo con Kribs (1935).
Análisis estadístico
Se generó una base de datos con las variables cuantitativas de las maderas, para someterlas a pruebas de inferencia. Los análisis estadísticos se realizaron mediante el paquete estadístico R v. 3.6.1 (R Core Team, 2019). Las variables cuantitativas se transformaron con logaritmo natural previo a los análisis, con el objetivo de estandarizar sus valores. Para evaluar si existen diferencias estadísticas entre las variables cuantitativas obtenidas de las maderas estudiadas, se llevó a cabo un modelo lineal generalizado (glm). La selección de esta técnica estadística se basó en el incumplimiento de supuestos de normalidad y homocedasticidad que presentó la base de datos (Seltman, 2018). Asimismo, se llevó a cabo un Análisis Discriminante Lineal (LDA) mediante la librería “MASS” de R, que permite generar funciones discriminantes, con las que se evalúa si las variables analizadas son suficientes para caracterizar a las especies estudiadas (Palacio et al., 2020); también se solicitó la matriz de confusión que indica el número de observaciones clasificadas correcta e incorrectamente.
Con el objetivo de determinar si las especies estudiadas se agrupan con base en sus atributos anatómicos cuantitativos, con especies pertenecientes al mismo género descritas en otras publicaciones (Barajas-Morales y León-Gómez, 1989; Carlquist, 1992b; Rebollar-Domínguez et al., 1993; Barajas-Morales et al., 1997; Abundiz-Bonilla et al., 2004; Ceja-Romero y Pérez-Olvera, 2010; Durán-Guerra et al., 2014; Rodríguez et al., 2016), se realizó un análisis jerárquico aditivo de conglomerado (Zelterman, 2015), mediante una matriz de distancias euclidianas de las medias de cada variable (Crisci y López-Armengol, 1983); los grupos formados se obtuvieron mediante el promedio aritmético no ponderado de cada grupo (UPGMA). Finalmente, tomando como base las descripciones de las maderas y los caracteres cuantitativos significativos en los análisis estadísticos, se construyó una clave de identificación para las especies estudiadas.
Resultados
Los caracteres anatómicos cuantitativos de las nueve especies analizadas se resumen en el Cuadro 2 e ilustran en las Figuras 2, 3 y 4. Los resultados del análisis de conglomerado se muestran en la Figura 5.
Cuadro 2: Caracteres cuantitativos de las especies estudiadas en la Sierra del Tentzo, Puebla, México. LV: Longitud de elementos de vaso, DVMT: Diámetro de los vasos en la madera temprana, DVMTR: Diámetro de los vasos en la madera tardía, PV: Pared de los Vasos, DP: Diámetro de las Punteaduras, LF: Longitud de Fibras, DF: Diámetro de Fibras, PF: Pared de Fibras, ANR: Ancho de Radios, LR: Altura de Radios. Para las especies con porosidad difusa, el diámetro de los vasos se reporta en la columna de vasos de madera temprana. En negritas se señalan los valores mínimos y máximos reportados para las especies.
| Vasos (µm) | Fibras (µm) | Radios (µm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Familia | Especie | LV | DVMT | DVMTR | PV | DP | LF | DF | PF | ANR | LR |
| Apocynaceae | Cascabela ovata (Cav.) | 287.3 | 44.1 | - | 2.68 | 3.92 | 331.65 | 8.56 | 2.01 | 14.99 | 116.28 |
| Lippold | (8) | (3.7) | (0.5) | (0.5) | (26) | (2) | (0.5) | (5) | (10.1) | ||
| Asteraceae | Critoniopsis uniflora (Sch. | 148.4 | 72.43 | 32.61 | 5.36 | 2.64 | 359.58 | 7.31 | 4.39 | 50.64 | 253.29 |
| Bip.) H. Rob. | (13) | (4) | (3.1) | (1.6) | (0.7) | (39) | (2) | (2) | (9) | (13) | |
| Berberidaceae | Berberis moranensis | 132.3 | 52.15 | 24.38 | 3.56 | 3.61 | 414.09 | 6.18 | 3.68 | 171.32 | >1000 |
| Schult. & Schult. f. | (15) | (9.5) | (2.7) | (1) | (0.7) | (2) | (3) | (2) | (21) | ||
| Burseraceae | Bursera fagaroides | 358.3 | 67.6 | 27.67 | 2.27 | 9.06 | 661.19 | 14.15 | 2.07 | 49.22 | 448.11 |
| (Kunth) Engl. | (13) | (14) | (4.3) | (1.1) | (1.5) | (26) | (1.5) | (0.5) | (16) | (74) | |
| Bursera morelense | 335.8 | 87.7 | - | 2.51 | 7.3 | 608 | 18.65 | 2.05 | 58.07 | 456.66 | |
| Ramírez | (10) | (8) | (0.5) | (1) | (45) | (2.2) | (0.7) | (21) | (14) | ||
| Convolvulaceae | Ipomoea murucoides | 161.9 | 125 | 47.8 | 7 | 7.75 | 767 | 17.09 | 4.55 | 250 | 1240 |
| Roem. & Schult. | (7) | (27) | (11.3) | (1.36) | (0.5) | (62.5) | (1.5) | (0.8) | (11) | (85) | |
| Fouquieriaceae | Fouquieria formosa Kunth | 407.8 | 56.6 | - | 2.84 | 6.85 | 334.2 | 10.94 | 4 | 196.67 | 166.16 |
| (32) | (1) | (0.5) | (1.5) | (48) | (3) | (1) | (22) | (12) | |||
| Rhamnaceae | Karwinskia humboldtiana | 104.9 | 33.4 | - | 4.04 | 4.71 | 380.15 | 8.74 | 3.5 | 37.62 | 162.13 |
| (Schult.) Zucc. | (4) | (9) | (1) | (0.8) | (17) | (2) | (1) | (3.5) | (9.2) | ||
| Solanaceae | Lycianthes sp. | 149.5 | 57.5 | 20.16 | 3.45 | 3.78 | 493.79 | 10.63 | 4.03 | 33.27 | 583.36 |
| (6) | (5.4) | (6.1) | (1) | (0.6) | (18) | (3.5) | (1) | (8.1) | (40) | ||
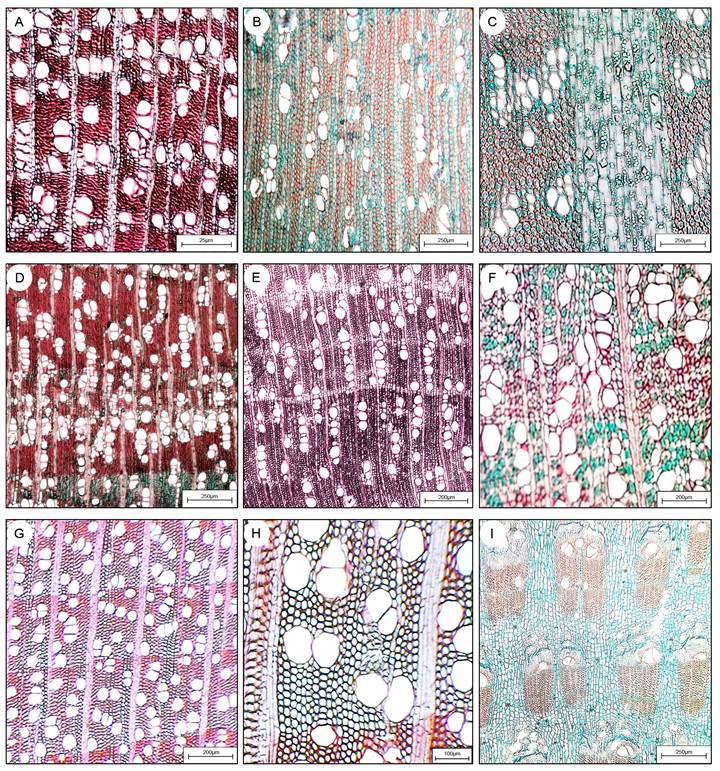
Figura 2: Secciones transversales de las maderas de las especies estudiadas en la Sierra del Tentzo, Puebla, México. A. Critoniopsis uniflora (Sch. Bip.) H. Rob.; B. Lycianthes sp.; C. Berberis moranensis Schult. & Schult. f.; D. Karwinskia humboldtiana (Schult.) Zucc.; E. Cascabela ovata (Cav.) Lippold; F. Fouquieria formosa Kunth; G. Bursera morelense Ramírez; H. Bursera fagaroides (Kunth) Engl.; I. Ipomoea murucoides Roem. & Schult. Escalas: A=25 µm; B-D, G, I=250 µm; E-F=200 µm, H=100 µm. Flechas=límite del anillo.
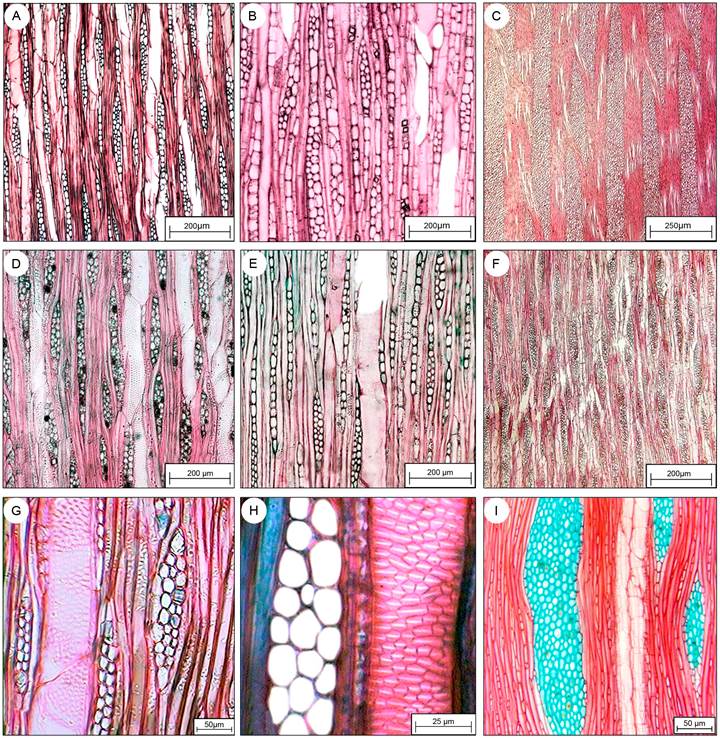
Figura 3: Secciones longitudinales-tangenciales de las especies estudiadas en la Sierra del Tentzo, Puebla, México. A. Criptoniopsis uniflora (Sch. Bip.) H. Rob.; B. Lycianthes sp.; C. Berberis moranensis Schult. & Schult. f.; D. Karwinskia humboldtiana (Schult.) Zucc.; E. Cascabela ovata (Cav.) Lippold, F. Fouquieria formosa Kunth; G. Bursera morelense Ramírez; H. Bursera fagaroides (Kunth) Engl.; I. Ipomoea murucoides Roem. & Schult. Escalas: A-B, D-F=200 µm, C=250 µm, G, I=50 µm, H=25 µm.
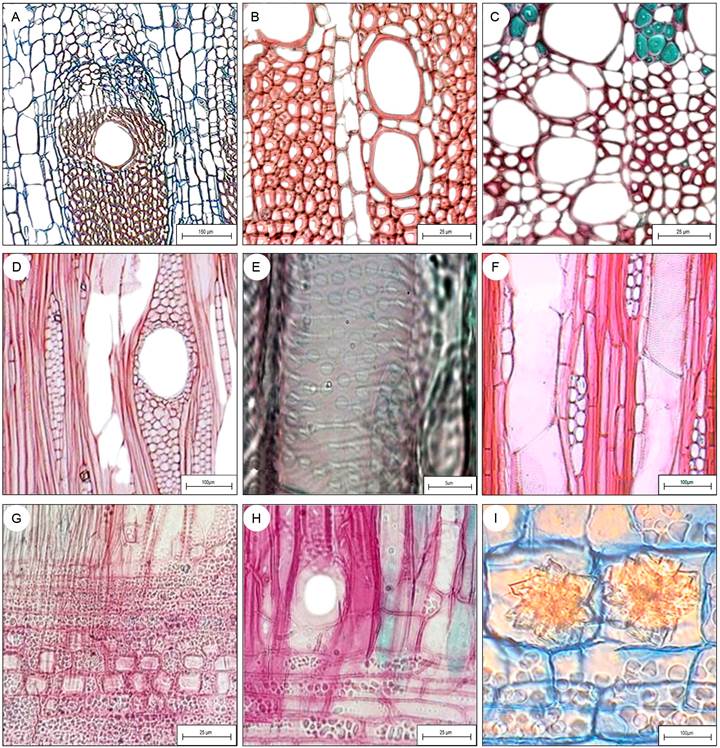
Figura 4: Detalles de diferentes secciones anatómicas de la madera de las especies estudiadas en la Sierra del Tentzo, Puebla, México. A. Floema conductor y no conductor y tejido conjuntivo (Tr), Ipomoea murucoides Roem. & Schult., B. Parénquima vasicéntrico (Tr), Criptoniopsis uniflora (Sch. Bip.) H. Rob., C. Parénquima apotraqueal difuso (Tr), Fouquieria formosa Kunth; D. Canal radial con células epiteliales (Ta), Bursera morelense Ramírez; E. Punteaduras areoladas, coalescentes (Ta), Berberis moranensis Schult. & Schult. f., F. Serie parenquimatosa de tres células (Ta), Critoniposis uniflora; G. Cristales prismáticos en células procumbentes (Ra), Karwinskia humboldtiana (Schult.) Zucc., H. Placa de perforación simple (Ra), Karwinskia humboldtiana; I. Drusas en el tejido conjuntivo (Tr), Ipomoea murucoides. Escalas: A, D, F, I = 100 µm, B, C, G, H=25 µm, E=5 µm. Secciones en las que se encuentra la figura: Tr=transversal, Ta=tangencial, Ra=radial.
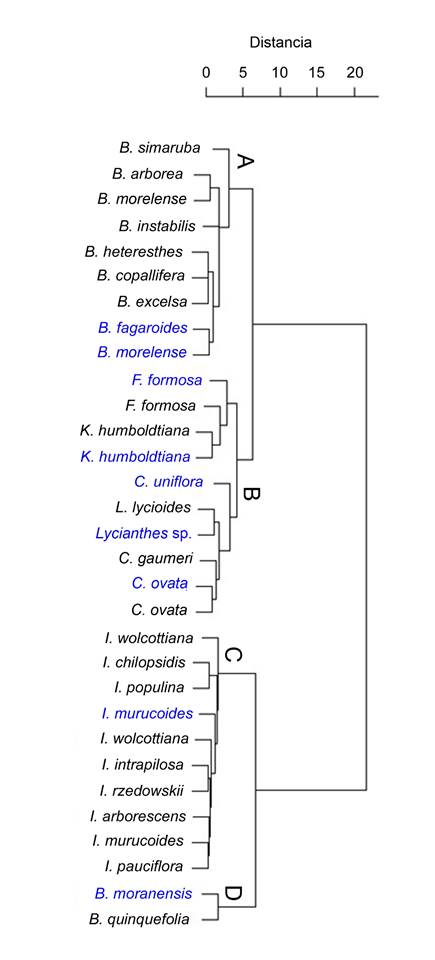
Figura 5: Dendrograma generado a partir de la matriz de distancias de las nueve especies estudiadas en la Sierra del Tentzo, Puebla, México (señaladas en azul) y de especies hermanas descritas en otros estudios. Recuperando cuatro grupos: A. especies del género Bursera Jacq. ex L., B. especies de los géneros Fouquieria Kunth, Karwinskia Zucc., Lycianthes (Dunal) Hassl., Cascabela Raf. y Critoniopsis Sch.Bip., C: Ipomoea L. arborescentes, D: Berberis L.
Marcas de crecimiento
En Berberis moranensis, Bursera fagaroides, B. morelense, Critoniopsis uniflora, Ipomoea murucoides y Lycianthes sp. las marcas fueron definidas por el diámetro de los vasos de la madera tardía y fibras radialmente comprimidas (Fig. 2A, B, C, G, H, I). Karwinskia humboldtiana y Cascabela ovata presentaron marcas de crecimiento delimitadas por 2-5 bandas de parénquima axial (Fig. 2D, E), mientras que Fouquieria formosa careció de ellas (Fig. 2F).
Vasos
La densidad de vasos/mm2 osciló entre 2 (Ipomoea murucoides) y 25 (Critoniopsis uniflora). Ipomoea murucoides presentó porosidad anular, con vasos de mayor diámetro en la madera temprana (Fig. 2I); mientras que B. morelense, F. formosa, K. humboldtiana y C. ovata presentaron porosidad difusa, para el resto de las especies fue semianular. Los vasos se encontraron principalmente formando hileras radiales en Critoniopsis uniflora (Fig. 2A), Lycianthes sp. (Fig. 2B), K. humboldtiana (Fig. 2D) y Cascabela ovata (Fig. 2E); en Berberis moranensis y F. formosa predominan los vasos agrupados (Figs. 2C, F). En las especies de Bursera (Figs. 2G, H) e I. murucoides (Fig. 2I), se encontraron principalmente vasos solitarios o formando grupos de dos o tres.
El diámetro del lumen de los vasos en las especies con porosidad difusa osciló entre 33.4 μm en K. humboldtiana y 87.7 μm en B. morelense. En las especies con porosidad semianular, los vasos de la madera temprana tuvieron diámetros de 52.15 μm (B. moranensis) a 125 μm (I. murucoides) y los de la madera tardía de 20.16 μm (Lycianthes sp.) hasta 47.8 μm (I. murucoides). El espesor de la pared de los vasos no presentó diferencias entre la madera temprana y tardía (P>0.05), observándose la más delgada en B. fagaroides (2.27 μm) y la más gruesa en I. murucoides (7 μm); especie en la que también se observaron vasos fibriformes. Los elementos de vaso registraron longitudes de 104.9 μm en K. humboldtiana a 407.8 μm en F. formosa. Todas las especies tuvieron punteaduras intervasculares alternas y areoladas (Figs. 3H, 4E), así como placas de perforación simple (Fig. 4H). Las punteaduras intervasculares presentaron diámetros de 2.64 μm en C. uniflora a 9.06 μm en B. fagaroides; también hubo punteaduras coalescentes en B. moranensis (Fig. 4E).
Fibras
Critoniopsis uniflora, Cascabela ovata, B. moranensis, I. murucoides, K. humboldtiana y Lycianthes sp. presentaron fibras libriformes no septadas con punteaduras simples (Fig. 3A, E, I), mientras que tres de ellas se observaron con septos (Fig. 3F, G). Las fibras más largas se observaron en I. murucoides (767 μm), mientras que C. ovata tuvo las más cortas (331.65 μm); el diámetro total de las fibras fue de 6.18 μm en B. moranensis hasta 18.65 μm en B. morelense. El grosor de la pared, oscilo entre 2.01 μm en C. ovata y 4.55 μm en I. murucoides. Las paredes de las fibras fueron clasificadas como delgadas a gruesas (Fig. 2H). Solo en B. fagaroides se observaron granos de almidón en las fibras.
Parénquima
Se identificó la presencia de parénquima axial apotraqueal difuso en Berberis moranensis, Bursera morelense, C. uniflora y F. formosa (p. ej., Fig. 4C). Respecto al parénquima axial paratraqueal, fue escaso en C. ovata, B. moranensis, B. morelense, B. fagaroides y K. humboldtiana, y vasicéntrico en C. uniflora, F. formosa, I. murucoides y Lycianthes sp. (Fig. 4B, C). En general, las series parenquimatosas incluyeron de dos a cuatro células (p. ej., Fig. 4F), excepto en Lycianthes sp., que presentó hasta diez células parenquimatosas.
Radios
Berberis moranensis presentó radios homocelulares Tipo I-B, para el resto de las especies se reportan radios heterocelulares Tipo II-B. La densidad fue desde 2 radios/mm en B. moranensis hasta 14 radios/mm en Lycianthes sp. Los radios presentaron alturas de 116.28 μm en C. ovata a >1000 µm en B. moranensis (Fig. 3C). Berberis moranensis también tuvo los radios más anchos (250 μm), mientras que los más delgados fueron los de C. ovata (14.99 μm). Se identificó la presencia de cristales prismáticos en el parénquima radial de C. uniflora, I. murucoides, B. moranensis, Lycianthes sp. (Fig. 3B), B. morelense (Figs. 3G, 4D) y K. humboldtiana (Fig. 4G), también se observaron drusas en I. murucoides (Fig. 4I) y contenidos obscuros en K. humboldtiana (Fig. 3D), así como canales radiales en las especies de Bursera (Fig. 4D).
Variante cambial
Ipomoea murucoides presentó cambia sucesivos (Figs. 2I, 4A) que forman haces concéntricos rodeados por una matriz de tejido conjuntivo no lignificado, cuyas células de parénquima contienen abundantes drusas (Fig. 4I). En el floema se distinguieron dos tipos: conductor y no conductor (Fig. 4A).
Análisis estadístico
El modelo lineal generalizado indicó que no existen diferencias intraespecíficas significativas, pero, sí interespecíficas; para todas las variables cuantitativas de las maderas (P<0.05), por lo que estas fueron incluidas en el análisis discriminante. Bursera morelense y B. fagaroides solamente presentan diferencias en los diámetros de los vasos (P=6.85e-08), punteaduras intervasculares (P<2e-16) y fibras (P=7.82e-09), así como en la longitud de los elementos de vaso (P<2e-16).
Mediante el análisis discriminante lineal, se identificó que las tres primeras funciones explican 88.64% de la traza de la matriz de datos (Cuadro 3). La primera función explicó 55.18% y asignó mayor peso al diámetro de las punteaduras intervasculares. La segunda mostró 24.05% e indicó que las paredes de vasos y fibras tienen los mayores coeficientes. Para la tercera, el ancho de radios y diámetro de las fibras son las variables más importantes, y acumulan 9.41% de la traza (λ-Wilks=0.03). La matriz de confusión (Cuadro 4) indicó que la precisión de la clasificación resultante del análisis LDA fue de 86.93%. Las observaciones de I. murucoides presentaron el mayor porcentaje de clasificaciones correctas con 96%, mientras que la especie con más confusión fue F. formosa, con 72% de sus observaciones clasificadas correctamente.
Cuadro 3: Coeficientes de las tres primeras funciones discriminantes lineales para las variables anatómicas cuantitativas de las nueve especies estudiadas en la Sierra del Tentzo, Puebla, México. Se indica el porcentaje de la traza explicado por cada función entre paréntesis.
| Variable | LD1 | LD2 | LD3 |
|---|---|---|---|
| (55.18%) | (24.05%) | (9.41%) | |
| Diámetro de los vasos de la madera temprana | -0.0054 | -0.009 | 0.0128 |
| Pared de vasos | 0.1828 | -0.289 | 0.0937 |
| Diámetro de punteaduras | -0.70004 | -0.3756 | -0.6415 |
| Diámetro de fibras | -0.0818 | -0.0667 | 0.0941 |
| Pared de fibras | 0.1853 | -0.1849 | -0.0669 |
| Altura de radios | -0.0006 | 0.0004 | 0.0012 |
| Ancho de radios | -0.0119 | 0.0206 | 0.013 |
| Largo de fibras | -0.0001 | -0.0003 | 0.0001 |
| Largo de elementos de vaso | -0.0053 | 0.0023 | 0.0054 |
Cuadro 4: Matriz de confusión resultante del análisis discriminante lineal (LDA). En la diagonal de la matriz se presenta la proporción de correctas clasificaciones. Todas las proporciones están expresadas en forma de porcentajes. 1=Bursera fagaroides (Kunth) Engl., 2=Berberis moranensis Schult. & Schult. f., 3=Bursera morelense Ramírez, 4=Critoniopsis uniflora (Sch. Bip.) H. Rob., 5=Fouquieria formosa Kunth, 6=Ipomoea murucoides Roem. & Schult., 7=Karwinskia humboldtiana (Schult.) Zucc., 8=Lycianthes sp., 9=Cascabela ovata (Cav.) Lippold.
| Especie predicha | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Especie observada | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| 1 | 90.66 | 0 | 2.66 | 0 | 5.33 | 0 | 0 | 0 | 1.33 |
| 2 | 0 | 92 | 0 | 0 | 0 | 0 | 2.66 | 5.33 | 0 |
| 3 | 4.54 | 0 | 95.45 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4 | 0 | 1.33 | 0 | 84 | 0 | 0 | 1.33 | 12 | 1.33 |
| 5 | 10 | 0 | 2 | 0 | 72 | 2 | 0 | 12 | 2 |
| 6 | 0 | 0 | 0 | 0 | 0 | 96 | 1.33 | 2.66 | 0 |
| 7 | 0 | 0 | 0 | 4 | 0 | 0 | 92 | 4 | 0 |
| 8 | 0 | 0 | 1.33 | 4 | 0 | 0 | 12 | 76 | 6.66 |
| 9 | 0 | 0 | 0 | 0 | 0 | 0 | 5.88 | 9.8 | 84.31 |
El análisis de conglomerados, donde también se incluyen los taxones descritos en publicaciones anteriores devolvió la formación de cuatro grupos (Fig. 5). El primero (A), formado por las especies analizadas de Bursera Jacq. ex L. El grupo B con Fouquieria Kunth, Karwinskia Zucc., Lycianthes (Dunal) Hassl., Cascabela Raf. y C. uniflora; el C incluyó a todos los miembros de Ipomoea L. y en el grupo D, a los de Berberis L.
Discusión
Características anatómicas como marcas de crecimiento, agrupamiento de los vasos, placas de perforación simple y elementos de vaso cortos, son típicas de especies que se distribuyen en sitios con estacionalidad marcada (Barajas-Morales y León-Gómez, 1989; Beeckman, 2016). Ya que son una estrategia de las plantas para disminuir eventos de cavitación, haciendo menos eficiente, pero más segura la conducción de agua y otros compuestos (Carlquist, 2012).
Los caracteres de la madera de las especies analizadas permitieron como en otros trabajos formar agrupaciones basadas en similitud, a través de técnicas estadísticas multivariadas (Baas et al., 1983; Terrazas y Wendt, 1995; Ruiz-Valencia et al., 2021). En el dendrograma obtenido (Fig. 5), se observa que los géneros Bursera, Berberis, Lycianthes, Karwinskia e Ipomoea se agruparon con base en las distancias globales, producto de los caracteres anatómicos reportados en el Apéndice. Para grupos que cuentan con pocas descripciones como Lycianthes y Cascabela, futuros estudios de la anatomía de especies hermanas permitirán determinar si estos géneros presentan homogeneidad anatómica como en Bursera (Pace et al., 2022).
En México existen descripciones anatómicas de ocho especies de Bursera (Barajas-Morales y León-Gómez, 1989; Barajas-Morales et al., 1997; Abundiz-Bonilla et al., 2004; Durán-Guerra et al., 2014). Las cuales indican que es un género anatómicamente homogéneo, ya que los estudios corresponden a taxones presentes en distintos tipos de vegetación, en los que caracteres como vasos solitarios, punteaduras alternas y areoladas, placas de perforación simple, porosidad difusa y radios heterocelulares tipo II-B son constantes en todos los hábitats. También se sabe que la presencia de canales de resina radiales son un carácter típico de la madera de Bursera (Durán-Guerra et al., 2014; Pace et al., 2022). La homogeneidad anatómica de las maderas de este género se vio reflejada en el dendrograma resultante del análisis de conglomerados. A pesar de que B. simaruba Sarg. se incluyó en el conglomerado A (Fig. 5), es la especie con menor similitud en este. Esta distancia puede deberse a diferencias ambientales entre los sitios de recolecta, ya que los especímenes analizados se colectaron en un bosque tropical perennifolio (Barajas-Morales et al., 1997), ecosistema en el que las condiciones abióticas son distintas a las del matorral xerófilo o la selva baja caducifolia, asociación vegetal en la que se distribuye la mayor parte de las especies de Bursera estudiadas a la fecha (Barajas-Morales y León-Gómez, 1989; Barajas-Morales et al., 1997; Abundiz-Bonilla et al., 2004; Durán-Guerra et al., 2014).
Se sabe que el diámetro de los vasos de Bursera se encuentra entre 90-168 μm. Sin embargo, tanto B. morelense como B. fagaroides, aquí descritas, presentaron vasos con diámetros menores a los reportados previamente (Cuadro 2). La longitud de las fibras en Bursera estudiadas en México es ≥700 μm (Barajas-Morales y León-Gómez, 1989; Barajas-Morales et al., 1997; Abundiz-Bonilla et al., 2004; Durán-Guerra et al., 2014), mientras que las longitudes aquí reportadas fueron de 608 μm (B. morelense) y 661 μm (B. fagaroides), respectivamente. Sin embargo, no son las más cortas reportadas para el género, ya que en B. simaruba miden 114 μm (Barajas-Morales et al., 1997). En el presente trabajo se obtuvo el primer registro de altura de los radios, siendo de 458.87 μm en B. morelense y 384.4 en B. fagaroides (Apéndice). Así como la presencia de cristales prismáticos en los radios y granos de almidón en las fibras de B. fagaroides. Diversos autores han señalado la importancia de los cristales prismáticos para discriminar entre especies (Cuéllar-Cruz et al., 2020; Ruiz-Valencia et al., 2021), por lo que se deben analizar más especímenes de B. fagaroides y otras especies del género para verificar si los cristales son un carácter relevante en la sistemática de Bursera.
Carlquist (1995) describió la madera de 21 especies de Berberis, señalando que el género se caracteriza por presentar porosidad semianular y vasos agrupados; caracteres también observados en el presente trabajo, específicamente en B. moranensis. Además, Carlquist (1995) indicó que las fibras de Berberis son aproximadamente el doble de largo que los elementos de vaso. Sin embargo, en los resultados de este trabajo encontramos una relación 3:1, en estos tipos celulares. Se sabe que la relación entre la longitud de vasos y fibras depende del tamaño de las células iniciales fusiformes (Honjo et al., 2005), las cuales están correlacionados con la edad de la planta y factores ambientales (Ohshima et al., 2011); posiblemente estas sean las razones de las diferencias reportadas. Carlquist (1995) también señaló que resulta complicado medir la altura de los radios de Berberis, ya que al alcanzar más de 3000 μm, la mayoría no son observables en el microscopio óptico; esto coincide con la descripción hecha aquí para B. moranensis con radios de más de 1000 µm de alto. Al respecto, Nowicke y Skvarla (1981) atribuyen las dimensiones de los radios de Berberis a la ancestría herbácea de Berberidaceae; origen que ha sido corroborado en estudios filogenéticos (Kim et al., 2004), validando así la explicación referente la altura de los radios en este género.
En México, Berberis incluye ca. 40 especies (Zamudio, 2009) y únicamente se había estudiado la madera de dos de ellas: B. quinquefolia (Standl.) Marroq. & Laferr., procedente de un matorral xerófilo (Abundiz-Bonilla et al., 2004), y B. chochoco Schltdl., de una selva baja caducifolia (Rodríguez et al., 2016). Con los datos disponibles, se aprecia que estas especies comparten con B. moranensis el mismo tipo de punteaduras intervasculares y placas de perforación simple. Con respecto a los caracteres cuantitativos, solo se tiene reporte de B. quinquefolia, la cual presenta valores superiores a los de B. moranensis en cuanto a densidad, diámetro y longitud de los elementos de vaso (Abundiz-Bonilla et al., 2004; Apéndice). Los caracteres cuantitativos contribuyeron al agrupamiento de los taxones de Berberis (Fig. 5); aunque se requiere estudiar un mayor número de especies del género para atribuir este agrupamiento a una posible homogeneidad anatómica.
En Ipomoea murucoides se encontraron vasos fibriformes, solitarios, punteaduras alternas, parénquima vasicéntrico, radios heterocelulares Tipo II-B y drusas, corroborando así lo publicado por Carlquist y Hanson (1991), quienes señalan que dichos caracteres definen en parte a la familia Convolvulaceae. Esos mismos autores presentaron la descripción de I. murucoides, con valores menores a los reportados aquí, con excepción del diámetro de los vasos y la altura de los radios (Apéndice). Las características de los cambia sucesivos, observados en I. murucoides ya han sido reportados previamente (Carlquist y Hanson, 1991; Ceja-Romero y Pérez-Olvera, 2010), también para otros miembros del género (Terrazas et al., 2011). Este carácter se puede explicar por el hábito trepador del grupo, debido a que el tejido conjuntivo protege al vascular del daño producido por la torsión de las lianas, además de ser una fuente de almacenamiento de agua (Schenck, 1893). La presencia de vasos fibriformes, indica eficiencia y seguridad en la conducción, ya que estos tipos celulares resisten los embolismos como las traqueidas y al contar con placas de perforación, son más eficientes para conducir agua sin que se produzca una ruptura en la columna de agua (Carlquist, 2012).
En México se reconocen 11 especies arborescentes de Ipomoea (Carranza et al., 1998), de las cuales existen descripciones anatómicas para ocho (Barajas-Morales y León-Gómez, 1989; Ceja-Romero y Pérez-Olvera, 2010). Las dimensiones de elementos de vaso y fibras muestran variaciones con base en las comparaciones con trabajos previos (Apéndice); las cuales pueden estar relacionadas con el tamaño de las plantas y las condiciones ambientales contrastantes de los sitios en los que se han recolectado, como se ha demostrado en otros estudios (Carlquist, 1988; Olson y Rosell, 2012; Beeckman, 2016). A pesar de las diferencias significativas observadas por Ceja-Romero y Pérez-Olvera (2010), en los caracteres cuantitativos entre especies de Ipomoea, el análisis de similitud indica que los miembros de este género analizados hasta el momento forman un grupo (Fig. 5) con alta similitud entre ellos. Con base en lo anterior, se puede considerar que como ocurre en Bursera, las ipomoeas arborescentes mexicanas presentan homogeneidad anatómica.
Con base en las descripciones anatómicas de Cascabela ovata (Barajas-Morales y León-Gómez, 1989) y C. gaumeri (Hemsl.) Lippold (Rebollar-Domínguez et al., 1993), existe similitud entre ellas y C. ovata de la Sierra del Tentzo. Los caracteres compartidos son ocho: porosidad semianular, vasos agrupados en múltiples radiales, elementos de vaso con placas de perforación simple, punteaduras areoladas, fibras libriformes, parénquima paratraqueal escaso, apotraqueal difuso y en bandas marginales, así como radios heterocelulares tipo II-B. De acuerdo con Barajas-Morales y León-Gómez (1989), C. ovata presenta gomorresinas en vasos y radios, además de gomas y cristales en el parénquima axial. En el presente estudio no se observaron dichos contenidos, debido quizá a las diferencias ambientales entre los sitios de colecta (Carlquist, 2012).
Con respecto a los caracteres cuantitativos, los contrastes son notables en las dimensiones de los vasos; C. ovata tiene valores menores a los reportados anteriormente, por Barajas-Morales y León-Gómez (1989) (Cuadro 2). Las diferencias cuantitativas en una misma especie pueden deberse a relaciones alométricas entre el tamaño de la planta y el de los vasos (Carlquist, 2012); para el caso del ejemplar estudiado por Barajas-Morales y León-Gómez (1989), midió entre 6 y 9 m, mientras que el ejemplar de C. ovata aquí analizado no excede de 4 m.
La anatomía de la madera de Fouquieria se caracteriza por presentar placas de perforación simple, fibrotraqueidas, parénquima apotraqueal difuso y radios heterocelulares (Carlquist, 2001a), coincidiendo con lo encontrado aquí para F. formosa. En este estudio, los vasos tuvieron un diámetro de 56.6 μm, similar al intervalo registrado en el género (25-71 μm); lo mismo ocurrió con el espesor de la pared y la longitud de los elementos de vaso. Sin embargo, las fibras y los radios son menores a lo reportado por Carlquist (2001a), quien registró valores variables para estos dos caracteres; mediante análisis correlativos se podría elucidar la causa de dicha variación entre especies (Ruiz-Valencia et al., 2021). Con respecto a F. formosa proveniente de un matorral xerófilo, Abundiz-Bonilla et al. (2004) describieron su madera con mayores dimensiones en los elementos de vaso, fibras y radios que las aquí registradas (Apéndice). Estas diferencias se reflejaron en el dendrograma, ya que a pesar de que ambas están en el grupo B, el espécimen de F. formosa descrito por Abundiz-Bonilla et al. (2004) tuvo mayor similitud con K. humboldtiana. Este ordenamiento puede reflejar la respuesta de las maderas a condiciones ambientales contrastantes entre los sitios de colecta, como se ha mencionado para otras especies (Beeckman, 2016). En este caso, el matorral xerófilo de Tehuacán (sitio de colecta de Abundiz-Bonilla et al. (2004)) capta aproximadamente 478 mm de agua al año, mientras que en la Sierra del Tentzo la precipitación no excede 200 mm al año (Morón et al., 2000).
La anatomía de la madera de Karwinskia humboldtiana de Tehuacán (Abundiz-Bonilla et al., 2004) muestra que los caracteres cualitativos coinciden con los registrados en este estudio para la misma especie y permiten que los especímenes se agrupen (Fig. 5). Sin embargo, las variables cuantitativas muestran diferencias importantes. En Tehuacán se reportan valores mayores a los encontrados aquí para vasos, fibras y radios (Apéndice); como ya se mencionó, puede deberse a las diferencias ambientales entre ambos sitios (Carlquist, 2012; Beeckman, 2016).
Los caracteres registrados en este estudio para Lycianthes sp. coinciden con las descripciones hechas para maderas de la familia Solanaceae, que se caracterizan por presentar placas de perforación simple, vasos ligeramente angulosos, punteaduras vaso-radio similares a las intervasculares, radios heterocelulares tipo II-B, así como cristales prismáticos en parénquima, fibras y radios (Carlquist, 1992a). La descripción de Lycianthes sp. es la primera que se realiza para el género en nuestro país. De Perú se conocen las características de L. lycioides Hassl. (Carlquist, 1992a), que presenta una densidad de vasos considerablemente mayor (433 vasos/mm2) a lo registrado para Lycianthes sp. (9 vasos/mm2). Sin embargo, Lycianthes sp. muestra mayor diámetro en el lumen de los vasos (57.5 μm) (Apéndice). Estos taxones se agruparon mediante el análisis de conglomerados, las diferencias encontradas entre Lycianthes sp. y L. lycioides podrían ser explicadas en futuros estudios, que permitan analizar el efecto de su distribución geográfica y las relaciones filogenéticas. Además, se requiere ampliar las descripciones anatómicas del género para poder elucidar si taxones con distribución en México son homogéneos o presentan variación como respuesta a factores ambientales.
Los caracteres anatómicos de Critoniopsis uniflora coinciden con lo que señala Carlquist (1964, 1992b) para la tribu Vernonieae, en cuanto a la presencia de placas de perforación simple y punteaduras alternas con areolas circulares, parénquima paratraqueal escaso y vasicéntrico, así como radios heterocelulares tipo II-B. Sin embargo, los elementos de vaso de C. uniflora, no exceden de 150 μm de longitud y 60 μm de diámetro, contrastando con los datos de Carlquist (1964) que, para Vernonieae registró vasos con diámetro >70 μm y longitud de elementos de vaso que superan los 160 μm. Critoniopsis uniflora se agrupó con Lycianthes y Cascabela; al ser la primera descripción de la especie, se requieren trabajos posteriores que permitan identificar si C. uniflora se agrupa con otros especímenes que ocurren en ambientes similares o con sus especies hermanas.
Se sabe que conforme se incrementa la sequía, el diámetro de los vasos disminuye, el número de vasos por mm2 aumenta y las longitudes de los elementos de vaso y fibras se acortan (Carlquist, 1995, 2001b, 2012); todo con el fin de minimizar los eventos de embolismo. Esto puede explicar las diferencias encontradas entre las descripciones hechas para las mismas especies, pero colectadas en diferentes sitios; o deberse a relaciones alométricas, las cuales han sido discutidas en trabajos previos (Olson y Rosell, 2012) y deben ser corroboradas mediante modelos lineales en los que se incluyan más especies de los taxones estudiados en este trabajo. Por otro lado, las similitudes anatómicas observadas en las especies con sus respectivos congéneres tienen implicaciones taxonómicas (Carlquist, 2012; Beeckman, 2016; Ruiz-Valencia et al., 2021).
Mediante el análisis LDA se identificó que el diámetro de los vasos, punteaduras intervasculares y fibras, así como la pared de los vasos y el ancho de las fibras, permiten diferenciar especies. Con base en estos caracteres cuantitativos, además de algunos rasgos cualitativos de las maderas, se elaboró una clave dicotómica para identificarlas, la cual se presenta a continuación:
Clave de identificación de las maderas estudiadas en la Sierra del Tentzo
1a. Parénquima paratraqueal escaso, pared de las fibras <4 μm …………………………………………………..……………….…… 2
1b. Parénquima paratraqueal vasicéntrico, pared de las fibras ≥4 μm …………………………………………..………….….... 6
2a. Marcas de crecimiento delimitadas por 2-5 bandas de parénquima axial, vasos en hileras radiales, altura de los radios ≤200 μm, ancho de radios ≤40 μm, diámetro de vasos ≤50 μm ……………………………………....……….…... 3
2b. Marcas de crecimiento delimitadas por diámetro de vasos y fibras de madera tardía, vasos solitarios o agrupados, altura de los radios >200 μm, ancho de los radios >40 μm, diámetro de vasos >50 μm ……...................…. 4
3a. Pared de los vasos ≤3 μm, diámetro de las punteaduras ≤4 μm, longitud de las fibras ≤350 μm, longitud de los elementos de vaso ≥200 μm, sin cristales ni contenidos oscuros ………..…............. Cascabela ovata (Cav.) Lippold
3b. Pared de los vasos >3 μm, diámetro de las punteaduras >4 μm, longitud de las fibras >350 μm, longitud de los elementos de vaso <200 μm, presencia de cristales y contenidos oscuros ................................................................................. Karwinskia humboldtiana (Schult.) Zucc.
4a. Radios tipo II-B, pared de los vasos ≤3 μm, diámetro de las punteaduras ≥4 μm, longitud de las fibras >500 μm, diámetro de las fibras ≥10 μm ………..................……..…. 5
4b. Radios tipo I-B, pared de los vasos >3 μm, diámetro de las punteaduras <4 μm, longitud de las fibras <500 μm, diámetro de las fibras <10 μm …..................................................…………… Berberis moranensis Schult. & Schult. f.
5a. Ancho de radios ≥50 μm, diámetro de vasos ≥80 μm, pared de fibras ≥2.5 μm a ≤3 μm, diámetro de las punteaduras ≥4 μm a ≤8 μm, diámetro de las fibras ≥15 μm, ausencias de canales radiales ................................................................................................ Bursera morelense Ramírez
5b. Ancho de radios <50 μm, diámetro de vasos ≥50 μm a <80 μm, pared de fibras <2.5, diámetro de las punteaduras ≥8 μm, diámetro de las fibras <15 μm, presencia de canales radiales ............ Bursera fagaroides (Kunth) Engl.
6a. Altura de los radios ≤500 μm, longitud de las fibras ≤400 μm …..…..........................................................……………. 7
6b. Altura de los radios >500 μm, longitud de las fibras >400 μm ……………..………………..........……………………......……... 8
7a. Diámetro de vasos ≥60 μm, pared de vasos ≥5 μm, diámetro de fibras <10 μm, ancho de radios ≤100 μm, diámetro de punteaduras ≤5 μm .……….........…............................................... Critoniopsis uniflora (Sch. Bip.) H. Rob.
7b. Diámetro de vasos <60 μm, pared de vasos <5 μm, diámetro de fibras ≥10 μm, ancho de radios >100 μm, diámetro de punteaduras >5 μm …...................................................................................… Fouquieria formosa Kunth
8a. Diámetro de vasos >100 μm, pared de vasos >5 μm, diámetro de fibras ≤15 μm, ancho de radios >100 μm, diámetro de punteaduras >5 μm ..…...........................................................…… Ipomoea murucoides Roem. & Schult.
8b. Diámetro de vasos ≤100 μm, pared de vasos ≤5 μm, diámetro de fibras >15 μm, ancho de radios ≤100 μm, diámetro de punteaduras ≤5 μm ……............... Lycianthes sp.
1a. Parénquima paratraqueal escaso, pared de las fibras <4 μm …………………………………………………..……………….…… 2
1b. Parénquima paratraqueal vasicéntrico, pared de las fibras ≥4 μm …………………………………………..………….….... 6
2a. Marcas de crecimiento delimitadas por 2-5 bandas de parénquima axial, vasos en hileras radiales, altura de los radios ≤200 μm, ancho de radios ≤40 μm, diámetro de vasos ≤50 μm ……………………………………....……….…... 3
2b. Marcas de crecimiento delimitadas por diámetro de vasos y fibras de madera tardía, vasos solitarios o agrupados, altura de los radios >200 μm, ancho de los radios >40 μm, diámetro de vasos >50 μm ……...................…. 4
3a. Pared de los vasos ≤3 μm, diámetro de las punteaduras ≤4 μm, longitud de las fibras ≤350 μm, longitud de los elementos de vaso ≥200 μm, sin cristales ni contenidos oscuros ………..…............. Cascabela ovata (Cav.) Lippold
3b. Pared de los vasos >3 μm, diámetro de las punteaduras >4 μm, longitud de las fibras >350 μm, longitud de los elementos de vaso <200 μm, presencia de cristales y contenidos oscuros ................................................................................. Karwinskia humboldtiana (Schult.) Zucc.
4a. Radios tipo II-B, pared de los vasos ≤3 μm, diámetro de las punteaduras ≥4 μm, longitud de las fibras >500 μm, diámetro de las fibras ≥10 μm ………..................……..…. 5
4b. Radios tipo I-B, pared de los vasos >3 μm, diámetro de las punteaduras <4 μm, longitud de las fibras <500 μm, diámetro de las fibras <10 μm …..................................................…………… Berberis moranensis Schult. & Schult. f.
5a. Ancho de radios ≥50 μm, diámetro de vasos ≥80 μm, pared de fibras ≥2.5 μm a ≤3 μm, diámetro de las punteaduras ≥4 μm a ≤8 μm, diámetro de las fibras ≥15 μm, ausencias de canales radiales ................................................................................................ Bursera morelense Ramírez
5b. Ancho de radios <50 μm, diámetro de vasos ≥50 μm a <80 μm, pared de fibras <2.5, diámetro de las punteaduras ≥8 μm, diámetro de las fibras <15 μm, presencia de canales radiales ............ Bursera fagaroides (Kunth) Engl.
6a. Altura de los radios ≤500 μm, longitud de las fibras ≤400 μm …..…..........................................................……………. 7
6b. Altura de los radios >500 μm, longitud de las fibras >400 μm ……………..………………..........……………………......……... 8
7a. Diámetro de vasos ≥60 μm, pared de vasos ≥5 μm, diámetro de fibras <10 μm, ancho de radios ≤100 μm, diámetro de punteaduras ≤5 μm .……….........…............................................... Critoniopsis uniflora (Sch. Bip.) H. Rob.
7b. Diámetro de vasos <60 μm, pared de vasos <5 μm, diámetro de fibras ≥10 μm, ancho de radios >100 μm, diámetro de punteaduras >5 μm …...................................................................................… Fouquieria formosa Kunth
8a. Diámetro de vasos >100 μm, pared de vasos >5 μm, diámetro de fibras ≤15 μm, ancho de radios >100 μm, diámetro de punteaduras >5 μm ..…...........................................................…… Ipomoea murucoides Roem. & Schult.
8b. Diámetro de vasos ≤100 μm, pared de vasos ≤5 μm, diámetro de fibras >15 μm, ancho de radios ≤100 μm, diámetro de punteaduras ≤5 μm ……............... Lycianthes sp.
Conclusiones
Las especies analizadas presentan caracteres anatómicos cualitativos que corresponden con descripciones previas de los mismos taxones, o en especies filogenéticamente cercanas. Se determinó que los caracteres cuantitativos analizados difieren a nivel intra e interespecífico, corroborando así la importancia de la anatomía de la madera para diferenciar especies. Por otro lado, algunos de los caracteres anatómicos son típicos de organismos que se distribuyen en sitios con estacionalidad marcada como la selva baja caducifolia. Finalmente, el análisis de conglomerados demostró que el conjunto de caracteres estudiados agrupa a los géneros Bursera, Berberis, Karwinskia, Cascabela, Lycianthes, Fouquieria e Ipomoea, con base en caracteres anatómicos. Para corroborar la presencia de homogeneidad anatómica a nivel genérico, se requiere incrementar el número de individuos y sitios de recolecta. También se deberá poner a prueba, mediante análisis correlativos y de correspondencia, una posible interacción entre las condiciones ambientales de la SBC y los caracteres anatómicos de la madera.

