











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El amaranto pertenece a la familia Amaranthaceae, subfamilia Amaranthoideae, género Amaranthus L. (Carmona y Orsini-Velázquez, 2010). Este género es predominantemente tropical, se distribuye en todos los climas desde el nivel del mar hasta 3600 m (Mazón et al., 2003). Borsch et al. (2015) reportan que la familia Amaranthaceae se conforma por 82 géneros y 840 especies. Costea et al. (2001) indican que aproximadamente 40 especies de Amaranthus son nativas de América.
El género Amaranthus se caracteriza por incluir plantas herbáceas anuales o arbustivas con flores de diversos colores, de verde a morado o púrpura con distintas coloraciones intermedias; unisexuales, con flores masculinas y femeninas en la misma planta (monoicas) o en plantas diferentes (dioicas) (Mosyakin y Robertson, 1996). Además de su importancia botánica y taxonómica, el género ha acaparado gran interés a nivel mundial, debido al valor nutricional que presentan algunas de sus especies, mismas que son fuente importante de alimento como verdura y grano (Wesche-Ebeling et al., 1995). Dentro del género se encuentran plantas silvestres, semidomesticadas y domesticadas productoras de grano, que han contribuido en los procesos de diversificación debido al flujo génico entre ellas (Boege, 2009).
Para la identificación de especies dentro del género Amaranthus se han establecido caracteres reproductivos y vegetativos (Sauer, 1967; Walton, 1968). Entre los rasgos reproductivos de valor taxonómico se encuentran la inflorescencia, flores estaminadas, flores pistiladas e incluso el fruto. En términos generales, la identificación de sus especies se basa en características de las flores pistiladas debido a que presentan la mayoría de los caracteres diagnósticos (Sauer, 1955). Tucker y Sauer (1958) propusieron como caracteres los relacionados con tépalos (tamaño, forma del ápice y dirección), brácteas (tamaño y longitud de la vena media), frutos (base de las ramas del estilo y dirección de los estigmas) e inflorescencias (grosor, hábito, tamaño y densidad en flores). Iamonico (2012) señala qué caracteres morfológicos y cualitativos, entre ellos la estructura de la sinflorescencia, la longitud de la sinflorescencia terminal, y la relación entre bráctea-tépalo, permiten reconocer las distintas subunidades reconocibles como taxa. Respecto a la descripción morfológica de estructuras de importancia taxonómica para la identificación de A. hybridus L. y A. cruentus L. en el sureste de México (desde el estado de Chiapas, Tabasco, la Península de Yucatán), Belice, y Guatemala (zona centro y norte), Solís-Fernández et al. (2020) señalan para A. hybridus un largo de bráctea de 2.42-4.75 mm, largo del sépalo externo de 1.65-3.52 mm, y largo del sépalo interno de 1.19-2.58 mm; y en A. cruentus brácteas con largo de 1.69-3.39 mm, sépalos externos de 1.89-2.86 mm de largo, y sépalos internos de 1.16-1.91 mm de largo.
Respecto al origen de especies con valor alimenticio en el continente americano, evidencia antropológica señala que Mesoamérica fue una de las principales regiones de domesticación y comienzo de la agricultura (Kantún-Balam et al., 2013). Una de las primeras civilizaciones que se desarrollaron en esta área fue la Maya, la cual abarcó una gran extensión territorial que comprendió los países de Guatemala, Belice, El Salvador, la porción occidental de Honduras y, en México, los estados de Campeche, Chiapas, Tabasco, Quintana Roo y Yucatán (Ruz-Lhuillier, 1981; Drew, 2002). Esta civilización se caracterizó por el manejo de los recursos naturales, desarrollando diversos sistemas de producción agroforestal, ejemplo de ellos la milpa y el huerto familiar (Méndez-Ramón, 2007). Resultado de lo anterior, se desarrolló una rica y compleja agricultura con varias especies importantes para su alimentación, basada principalmente en maíz (Zea mays L.), frijol (Phaseolus sp.), calabaza (Cucurbita sp.), amaranto (Amaranthus sp.) y otras especies complementarias como cacao (Theobroma cacao L.), vainilla (Vanilla sp.) y chiles (Capsicum sp.), además de árboles frutales, hierbas y hongos (FAO, 2009).
Con respecto a la domesticación del amaranto, se cuenta con evidencia arqueológica que señala a México como uno de sus centros de origen y domesticación (Sauer, 1950). Por otro lado, Peralta (2009) sugiere que este proceso se pudo llevar a cabo en tres sitios: América del Norte (México), América Central (Guatemala) y América del Sur (Ecuador, Perú y Bolivia). En América tres especies son cultivadas como especie de grano, Amaranthus caudatus L. en los Andes de Perú, A. hypochondriacus L. en la región central de México (Peralta, 2009), y A. cruentus en el sur de México y Guatemala (Sauer, 1967), en la región Maya. Incluso en la zona centro de México su grano es usado como materia prima para la industria de dulces tradicionales como “la alegría” (Ayala et al., 2014). A pesar de ello, los reportes de la distribución de A. cruentus en el sureste de México son escasos, tan solo se ha reportado su presencia en el estado de Chiapas (Espitia-Rangel et. al, 2010b).
Sobre el origen de las especies cultivadas de amaranto, Sauer (1967) propuso dos hipótesis: la primera presenta a Amaranthus hybridus como ancestro común de todas ellas, mientras en la segunda, considera que cada especie procede de una especie silvestre donde A. caudatus deriva de A. quitensis Kunth, A. cruentus de A. hybridus y A. hypochondriacus de A. powellii S. Watson. Estudios a nivel molecular del ADN de A. hybridus y especies de amaranto productoras de grano usando marcadores RAPD e isoenzimas (Chan y Sun, 1997), microsatélites (Kietlinski et al., 2014), y SNP o Polimorfismo de Nucleotidos Unicos (Clouse et al., 2016) respaldan las afirmaciones anteriores, al señalar a A. hybridus como el ancestro común de las especies de grano de amaranto. Las especies cultivadas empleadas para la producción de grano son fácilmente reconocidas por presentar brácteas cortas y frágiles, inflorescencias de gran tamaño y semillas blancas, resultado de la selección artificial; a diferencia de las silvestres que presentan brácteas espinosas, tallas bajas de plantas e inflorescencias, reducida producción de semillas y semillas oscuras (Sauer, 1967); aunque el color oscuro puede observarse también en organismos semidomesticados o en proceso de domesticación (Stetter et al., 2019).
Sobre la diferenciación entre las especies cultivadas de grano, la morfología de la bractéola es uno de los pocos caracteres que ayudan en su identificación, especialmente entre A. caudatus y A. cruentus, e incluso entre sus parientes silvestres como A. hybridus, A. quitensis y A. powellii. De esta forma las brácteas en los amarantos de grano son menos espinosas y más cortas y delgadas que las de sus parientes silvestres (Sauer, 1950; Espitia-Rangel et al., 2010c).
Partiendo de la evidencia del origen de A. cruentus, su cercanía con culturas originarias de América, su importancia con estas culturas y las hipótesis que señalan a A. hybridus como su ancestro durante el proceso de domesticación y su pariente silvestre más cercano, el presente trabajo tiene como objetivo presentar evidencia sobre la distribución actualizada y la frecuencia de A. cruentus y A. hybridus, así como evaluar la diversidad morfológica de ambas especies tomando como base caracteres de flores pistiladas y del fruto.
Materiales y Métodos
Especímenes evaluados y reporte de distribución
Se utilizaron para el estudio un total de 147 ejemplares procedentes de los herbarios CAS, CICY, ENCB, F, MEXU, MO y NHM (acrónimos con base en Thiers, 2020+ continuamente actualizado), señalados con colectas en Guatemala, Honduras y México, para las especies Amaranthus cruentus y A. hybridus. Estos ejemplares fueron reportados como individuos de las especies A. cruentus y A. hybridus, identificados usando métodos de botánica tradicional, análisis de varianza permutacional multivariada PERMANOVA (por sus siglas en inglés, permutational multivariate variance analysis) y la prueba de porcentaje de similaridad SIMPER (por sus siglas en inglés; similarity percentage) mediante el uso de la paquetería estadística PRIMER v. 7 y PERMANOVA por Solís-Fernández et al. (2020).
Se actualizó la información de distribución y frecuencia de A. cruentus y A. hybridus en Guatemala, Honduras y el sureste de México; el reporte de su distribución extendida se realizó con la información recabada de las etiquetas de los ejemplares de herbario.
Estudio morfológico
El estudio de diversidad morfológica realizado en A. cruentus y A. hybridus se basó en la terminología y los 22 caracteres morfológicos (Cuadros 1, 2), reportados por Solís-Fernández et al. (2020), usando para ello imágenes digitales obtenidas a través de un microscopio estereoscopico Nikon (modelo SMZ745T, Tokio, Japón), cámara digital INFINITY1 (modelo 3C, Ontario, Canadá) y el programa INFINITY ANALYZE v. 5.0.3 software (Lumenera Corp., 2010).
Cuadro 1: Estadística descriptiva entre caracteres morfológicos relacionados a flores pistiladas de Amaranthus cruentus L. procedentes de Guatemala y México y el valor general de ambas procedencias usados comúnmente en la determinación taxonómica de la familia Amaranthaceae. Coeficiente de variación=CV, Largo de la Flor=LF, Largo de la Bráctea=LB, Ancho de la Bráctea=AB, Largo del Sépalo Externo=LSE, Ancho del Sépalo Externo=ASE, Largo del Ápice del Sépalo Externo=LASE, Largo de la Lámina del Sépalo Externo=LLSE, Índice del Sépalo Externo=ISE, Largo del Sépalo Interno=LSI, Ancho del Sépalo Interno=ASI, Largo del Ápice del Sépalo Interno=LASI, Largo de la Lámina del Sépalo Interno=LLSI, Índice del Sépalo Interno=ISI, Largo del Estilo=LE, Largo del Utrículo=LU, Ancho del Utrículo=AU, Relación entre el Largo con respecto a el Ancho del Utrículo=RLAU, Largo de la Semilla=LS, Ancho de la Semilla=AS, Relación entre el Largo de la Bráctea con respecto al largo del Sépalo Externo=RLBSE, Relación entre el Largo del Utrículo con respecto a el largo del Sépalo Externo=RLUSE, Relación entre el Largo de la Bráctea con respecto a el largo del Utrículo=RLBU.
| Carácter | Guatemala | México | General | |||||||||
| Media (mm) | Desviación estándar | Varianza | CV (%) | Media (mm) | Desviación estándar | Varianza | CV (%) | Media (mm) | Desviación estándar | Varianza | CV (%) | |
| LF | 1.97 | 0.46 | 0.21 | 23.30 | 1.49 | 0.35 | 0.12 | 23.33 | 1.90 | 0.47 | 0.22 | 24.52 |
| LB | 2.37 | 0.45 | 0.20 | 19.06 | 2.21 | 0.56 | 0.32 | 25.41 | 2.35 | 0.45 | 0.20 | 19.07 |
| AB | 0.49 | 0.22 | 0.05 | 43.77 | 0.43 | 0.09 | 0.01 | 21.38 | 0.48 | 0.20 | 0.04 | 41.78 |
| LSE | 2.22 | 0.35 | 0.12 | 15.58 | 2.07 | 0.22 | 0.05 | 10.62 | 2.20 | 0.33 | 0.11 | 15.01 |
| ASE | 0.58 | 0.14 | 0.02 | 23.54 | 0.68 | 0.13 | 0.02 | 18.72 | 0.59 | 0.14 | 0.02 | 22.89 |
| LASE | 0.25 | 0.12 | 0.01 | 47.43 | 0.13 | 0.06 | 0.00 | 43.51 | 0.23 | 0.12 | 0.01 | 50.49 |
| LLSE | 1.98 | 0.27 | 0.07 | 13.64 | 1.94 | 0.16 | 0.03 | 8.40 | 1.97 | 0.25 | 0.06 | 12.87 |
| ISE | 4.14 | 1.60 | 2.57 | 38.80 | 3.06 | 0.25 | 0.06 | 8.18 | 3.99 | 1.53 | 2.35 | 38.44 |
| LSI | 1.55 | 0.24 | 0.06 | 15.22 | 1.66 | 0.22 | 0.05 | 13.24 | 1.56 | 0.23 | 0.05 | 14.67 |
| ASI | 0.52 | 0.12 | 0.01 | 22.76 | 0.39 | 0.06 | 0.00 | 14.50 | 0.51 | 0.12 | 0.01 | 23.90 |
| LASI | 0.10 | 0.07 | 0.00 | 68.93 | 0.09 | 0.03 | 0.00 | 31.43 | 0.10 | 0.06 | 0.00 | 64.89 |
| LLSI | 1.45 | 0.27 | 0.07 | 18.59 | 1.57 | 0.19 | 0.04 | 12.20 | 1.46 | 0.26 | 0.07 | 17.60 |
| ISI | 3.12 | 1.01 | 1.02 | 32.30 | 4.33 | 1.19 | 1.42 | 27.49 | 3.28 | 1.07 | 1.15 | 32.71 |
| LE | 0.58 | 0.24 | 0.06 | 40.46 | 0.67 | 0.11 | 0.01 | 15.95 | 0.59 | 0.22 | 0.05 | 37.40 |
| LU | 1.63 | 0.24 | 0.06 | 14.81 | 1.64 | 0.03 | 0.00 | 1.72 | 1.63 | 0.22 | 0.05 | 13.71 |
| AU | 1.35 | 0.25 | 0.06 | 18.65 | 1.24 | 0.27 | 0.07 | 21.67 | 1.33 | 0.25 | 0.06 | 18.47 |
| RLAU | 1.23 | 0.19 | 0.03 | 15.06 | 1.36 | 0.32 | 0.10 | 23.35 | 1.25 | 0.20 | 0.04 | 15.72 |
| LS | 1.17 | 0.17 | 0.03 | 14.53 | 1.16 | 0.10 | 0.01 | 8.53 | 1.17 | 0.16 | 0.03 | 13.66 |
| AS | 1.07 | 0.18 | 0.03 | 16.77 | 1.14 | 0.21 | 0.04 | 18.07 | 1.08 | 0.18 | 0.03 | 16.37 |
| RLBSE | 1.30 | 0.22 | 0.05 | 16.90 | 1.30 | 0.20 | 0.04 | 15.39 | 1.30 | 0.21 | 0.04 | 16.17 |
| RLUSE | 1.07 | 0.20 | 0.04 | 18.59 | 1.21 | 0.01 | 0.00 | 0.65 | 1.09 | 0.19 | 0.04 | 17.49 |
| RLBU | 1.26 | 0.40 | 0.16 | 32.01 | 1.08 | 0.17 | 0.03 | 16.03 | 1.24 | 0.38 | 0.15 | 30.93 |
Cuadro 2: Estadística descriptiva entre caracteres morfológicos relacionados a flores pistiladas de Amaranthus hybridus L. procedentes de Guatemala y México y el valor general de ambas procedencias usados comúnmente en la determinación taxonómica de la familia Amaranthaceae. Coeficiente de variación=CV, Largo de la Flor=LF, Largo de la Bráctea=LB, Ancho de la Bráctea=AB, Largo del Sépalo Externo=LSE, Ancho del Sépalo Externo=ASE, Largo del Ápice del Sépalo Externo=LASE, Largo de la Lámina del Sépalo Externo=LLSE, Índice del Sépalo Externo=ISE, Largo del Sépalo Interno=LSI, Ancho del Sépalo Interno=ASI, Largo del Ápice del Sépalo Interno=LASI, Largo de la Lámina del Sépalo Interno=LLSI, Índice del Sépalo Interno=ISI, Largo del Estilo=LE, Largo del Utrículo=LU, Ancho del Utrículo=AU, Relación entre el Largo con respecto al Ancho del Utrículo=RLAU, Largo de la Semilla=LS, Ancho de la Semilla=AS, Relación entre el Largo de la Bráctea con respecto al largo del Sépalo Externo=RLBSE, Relación entre el Largo del Utrículo con respecto a el largo del Sépalo Externo=RLUSE, Relación entre el Largo de la Bráctea con respecto a el largo del Utrículo=RLBU.
| Carácter | Guatemala | México | General | |||||||||
| Media (mm) | Desviación estandar | Varianza | CV (%) | Media (mm) | Desviación estandar | Varianza | CV (%) | Media (mm) | Desviación estandar | Varianza | CV (%) | |
| LF | 2.28 | 0.48 | 0.23 | 21.27 | 1.94 | 0.25 | 0.06 | 12.62 | 2.23 | 0.47 | 0.22 | 21.11 |
| LB | 3.70 | 0.64 | 0.41 | 17.22 | 3.06 | 0.45 | 0.20 | 14.65 | 3.61 | 0.65 | 0.42 | 17.91 |
| AB | 0.65 | 0.15 | 0.02 | 22.75 | 0.61 | 0.10 | 0.01 | 16.73 | 0.65 | 0.14 | 0.02 | 22.09 |
| LSE | 2.61 | 0.52 | 0.27 | 19.81 | 2.11 | 0.54 | 0.29 | 25.55 | 2.54 | 0.54 | 0.29 | 21.13 |
| ASE | 0.65 | 0.15 | 0.02 | 22.71 | 0.53 | 0.17 | 0.03 | 32.57 | 0.63 | 0.15 | 0.02 | 24.15 |
| LASE | 0.31 | 0.19 | 0.04 | 62.06 | 0.14 | 0.08 | 0.01 | 53.86 | 0.29 | 0.19 | 0.04 | 65.79 |
| LLSE | 2.29 | 0.39 | 0.16 | 17.18 | 1.97 | 0.48 | 0.23 | 24.41 | 2.25 | 0.41 | 0.17 | 18.31 |
| ISE | 4.24 | 1.29 | 1.66 | 30.39 | 4.06 | 0.60 | 0.36 | 14.70 | 4.21 | 1.22 | 1.48 | 28.84 |
| LSI | 1.79 | 0.39 | 0.15 | 21.71 | 1.77 | 0.56 | 0.32 | 31.70 | 1.79 | 0.40 | 0.16 | 22.55 |
| ASI | 0.45 | 0.11 | 0.01 | 24.63 | 0.49 | 0.11 | 0.01 | 22.04 | 0.46 | 0.11 | 0.01 | 24.04 |
| LASI | 0.11 | 0.07 | 0.01 | 63.37 | 0.10 | 0.02 | 0.00 | 21.91 | 0.11 | 0.07 | 0.00 | 60.86 |
| LLSI | 1.67 | 0.35 | 0.12 | 20.92 | 1.68 | 0.55 | 0.30 | 32.90 | 1.68 | 0.37 | 0.14 | 22.09 |
| ISI | 4.20 | 1.52 | 2.31 | 36.25 | 3.58 | 0.48 | 0.23 | 13.50 | 4.12 | 1.44 | 2.07 | 34.98 |
| LE | 0.82 | 0.34 | 0.12 | 41.45 | 0.34 | 0.31 | 0.10 | 92.54 | 0.76 | 0.37 | 0.14 | 48.80 |
| LU | 1.51 | 0.27 | 0.07 | 17.94 | 1.51 | 0.36 | 0.13 | 23.86 | 1.51 | 0.28 | 0.08 | 18.33 |
| AU | 1.25 | 0.17 | 0.03 | 13.66 | 1.32 | 0.17 | 0.03 | 12.78 | 1.26 | 0.17 | 0.03 | 13.45 |
| RLAU | 1.21 | 0.21 | 0.04 | 17.40 | 1.14 | 0.24 | 0.06 | 20.69 | 1.21 | 0.21 | 0.04 | 17.58 |
| LS | 1.13 | 0.15 | 0.02 | 12.97 | 1.12 | 0.18 | 0.03 | 15.89 | 1.13 | 0.15 | 0.02 | 13.08 |
| AS | 1.09 | 0.16 | 0.03 | 14.68 | 1.00 | 0.17 | 0.03 | 16.78 | 1.08 | 0.16 | 0.03 | 14.94 |
| RLBSE | 1.45 | 0.39 | 0.15 | 26.54 | 1.37 | 0.81 | 0.65 | 59.06 | 1.44 | 0.44 | 0.20 | 30.62 |
| RLUSE | 0.83 | 0.17 | 0.03 | 20.05 | 0.95 | 0.11 | 0.01 | 11.51 | 0.84 | 0.16 | 0.03 | 19.36 |
| RLBU | 1.76 | 0.31 | 0.10 | 17.74 | 1.39 | 0.70 | 0.49 | 50.34 | 1.71 | 0.39 | 0.15 | 22.58 |
Análisis de datos
La diversidad morfológica se determinó mediante la morfometría de las estructuras florales pistiladas y el fruto, realizando análisis de estadística descriptiva y la determinación del coeficiente de variación. Se aplicó a esta misma información un análisis multivariado de componentes principales (CP). Para obtener la diversidad morfológica, los caracteres fueron normalizados (matriz de varianza) y se aplicó el coeficiente de distancia euclideana para su representación gráfica. El análisis multivariado se desarrolló con el paquete estadístico PRIMER v. 6 (Clarke y Gorley, 2006).
Resultados
Distribución y frecuencia de A. cruentus y A. hybridus
El Cuadro 3 presenta la distribución y frecuencia de ejemplares de A. cruentus y A. hybridus por país. Amaranthus cruentus (34 ejemplares en total) se distribuyó en Guatemala (27 ejemplares, 79%) en siete departamentos, la mayor cantidad en Baja Verapaz y Sacatepéquez, con nueve y ocho ejemplares respectivamente, así como en el sureste de México (siete ejemplares, 21%), de los cuales tres se ubican en Chiapas, tres en Yucatán y uno en Tabasco.
Cuadro 3: Distribución de registros de colecta de ejemplares de Amaranthus cruentus L. y A. hybridus L. en partes de la región del área Maya (suereste de México, Guatemala y Honduras) (acrónimos con base en Thiers, 2020+ continuamente actualizado).
| PÁIS | Amaranthus cruentus L. | Amaranthus hybridus L. | ||
| Depto. / Estado | Cant. | Colector, Núm. de colecta (Herbario) | Cant. | Colector, Núm. de colecta (Herbario) |
| GUATEMALA | 27 | 43 | ||
| Alta Verapaz | 4 | von Tüerckheim 8567 (F), Standley 90924(F); Sauer 1265-D (F); Sauer 1265-E (MO) | 5 | Standley 40806, 69273, 90147 y 91312 (F); Williams et al. 25336 (F) |
| Baja Verapaz | 9 | Sánchez-del Pino et al. 520, 521, 523, 524, 526, 526-1, 526-2, 527 y 530 (CICY) | 1 | Sánchez-del Pino et al. 528 (CICY) |
| Chimaltenango | 2 | Saur 1168-K (F); Sauer 1240-B (MO) | 5 | Standley 59043, 59046, 59076, 79743 y 80832 (F) |
| El Progreso | 1 | Juvenal 2304 (F) | 0 | -- |
| Guatemala | 1 | Sauer 1285-D (MO) | 2 | Sánchez-del Pino 518 (CICY); Brenckle 47428 (CAS) |
| Huehuetenango | 0 | -- | 4 | Williams et al. 41273 (F); Standley 82495, 82994 y 82995 (F) |
| Jalapa | 0 | -- | 4 | Standley 76360, 77139 y 77291 (F); Kellerman 7693 (F) |
| Jutiapa | 0 | -- | 2 | Standley 75135 y 75806 (F) |
| El Petén | 0 | -- | 1 | Landell 15787 (MEXU) |
| Quezaltenango | 0 | -- | 4 | Steyermark 34431 (F); Standley 83166, 83500 y 85879 (F) |
| Sacatepéquez | 8 | Standley 60352, 63056 y 64288 (F); Sauer 1175-B, 1176-L, 1177-A, 1177-D, 1176 (MO) | 11 | Sánchez-del Pino et al. 517, 517-A y 517-B (CICY); Standley 63760, 58041, 59853, 59944, 60327, 63260 y 64619 (F); Molina y Molina 27054 (F) |
| Santa Rosa | 0 | -- | 2 | Standley 77864 y 78252 (F) |
| Sololá | 0 | -- | 1 | Williams et al. 25336 (F) |
| Suchitepéquez | 0 | -- | 1 | Standley 88905 (F) |
| Zacapa | 2 | Standley 3955 y 73955 (F) | 0 | -- |
| HONDURAS | 0 | 15 | ||
| Comayagua | 0 | -- | 1 | Standley y J. Chacón 5666 (CAS) |
| El Paraíso | 0 | -- | 1 | Standley 16735 (CAS) |
| Francisco Morazán | 0 | -- | 12 | Standley 1660, 5199, 28485 y 29505-A (CAS); Standley y Williams 1400 (CAS); Rodríguez 416 y 1057 (F); de Belibasis 53 (MO); Nichols 2028 (MO); Rodríguez 117 y 159 (MO); Trochez 217 (MO) |
| Intibucá | 0 | -- | 1 | Standley 25600 (CAS) |
| MÉXICO | 7 | 55 | ||
| Campeche | 0 | -- | 16 | Sánchez-del Pino et al. 537 (CICY); Álvarez y Sánchez-del Pino et al. 4067, 4260 y 5192 (MEXU); Alvaro 744 (MEXU); Cabrera 11703, 14320, 14411 y 15977 (MEXU); Martínez 28326, 28690-A y 29285 (MEXU); Palmillas 999, 999-A y 999-B (MEXU); Martínez 2973 (MO) |
| Chiapas | 3 | Sánchez-del Pino et al. 532 (CICY); Calzada 2869 (ENCB); Breedlove 14649 (F) | 28 | Sánchez-del Pino et al. 535 y 535-C (CICY); Breedlove 6091 (F); Breedlove 7423 y 12785 (MEXU); Calzada 2768 (MEXU); Durán 546 (MEXU); García 643 (MEXU); Gómez 566 (MEXU); Martínez 9078 (MEXU); Martínez y Lambera 26150 (MEXU); Méndez 4763 y 6818 (MEXU); Sant 460; Shilom 419, 1570, 1737, 1738, 4497 y 7468 (MEXU); Sinaca 2258, 2414, 2427, 2414-A y 2414-B (MEXU); Breedlove 14732 y 41524 (MO); Zuil 758 (MO) |
| Quintana Roo | 0 | -- | 4 | Trejo 118 (CICY); Balam 451 (CICY); Ucan 4036 (CICY); Álvarez 10530 (MEXU) |
| Tabasco | 1 | Magaña 1468 (ENCB) | 2 | Sol 1125 (MEXU); Magaña 2456 (ENCB) |
| Yucatán | 3 | Simá y Sánchez-del Pino 06 (CICY); Simá 07 (CICY); Espejel 97 (F) | 5 | Simá y Sánchez-del Pino 04 (CICY); Darwin y Sundell 2052 (MO); Gaumer 2447 (F); Rivera 220 (MEXU); Gaumer 1081 (USCH) |
| Total | 34 | 113 |
Por otro lado, Amaranthus hybridus (113 ejemplares en total) se encontró en el sureste de México (55 ejemplares, 49%); en Guatemala (43 ejemplares, 38%) donde la mayor cantidad de ejemplares fueron reportados en el departamento de Sacatepéquez (11) y los 32 restantes se distribuyeron en otros 12 departamentos; y Honduras (15 ejemplares, 13%), principalmente en el departamento de Francisco Morazán, con un total de 12 ejemplares, mientras que los restantes fueron colectados en Comayagua, El Paraíso e Intibucá (Cuadro 3). En México, la mayor cantidad de ejemplares corresponde a los estados de Chiapas (28 ejemplares) y Campeche (16), en tanto que los estados de Quintana Roo, Tabasco y Yucatán presentaron los 11 ejemplares restantes.
Diversidad morfológica en A. cruentus y A. hybridus
El análisis exploratorio aplicado en estructuras morfológicas de flores pistiladas y frutos en A. cruentus reveló que las flores de mayor longitud fueron las provenientes de Guatemala (LF: 1.97 mm). De igual manera, las brácteas fueron más largas y anchas (LB: 2.37 mm y AB: 0.49 mm) (Cuadro 1).
En Amaranthus cruentus, el sépalo externo fue de mayor longitud en ejemplares colectados en Guatemala (LSE, LASE, LLSE e ISE), mientras que su ancho (ASE), fue mayor en ejemplares colectados en México (Cuadro 1). El sépalo interno (LSI, LLSI e ISI) fue de mayor tamaño en ejemplares colectados en México (Cuadro 1). El ancho del sépalo interno (ASI) y el largo del ápice del sépalo interno (LASI) fueron de mayor tamaño en ejemplares colectados en Guatemala (Cuadro 1). Los estigmas y utrículos más largos se observaron en ejemplares de México; solamente el ancho del utrículo fue mayor en ejemplares colectados en Guatemala (Cuadro 1). Las semillas colectadas en Guatemala fueron más largas, pero menos anchas que las procedentes de México (Cuadro 1). Tres relaciones fueron consideradas con base en la longitud de bráctea, sépalo externo y utrículo: (1) bráctea:sépalo externo (RLBSE), (2) utrículo:sépalo externo (RLUSE) y (3) bráctea:utrículo (RLBU). Los valores de RLBSE fueron similares en los ejemplares colectados en Guatemala y México, mientras que RLUSE fue mayor en México y RLBU fue mayor en Guatemala (Cuadro 1).
En Amaranthus cruentus el coeficiente de variación (CV) osciló entre 12.87% (LLSE) y 64.89% (LASI) (Cuadro 1). En ejemplares de Guatemala la variación del CV más alta se observó en LASI (68.93%) y la menor en LLSE (13.64%); para México los valores del CV fluctuaron entre 43.51% (LASE) y 0.65% (RLUSE) (Cuadro 1).
En Amaranthus hybridus se observó mayor tamaño en las estructuras de flores pistiladas de ejemplares colectados de Guatemala (Cuadro 2), referente a el largo de la flor (LF: 2.28 mm), largo de las brácteas (LB: 3.70 mm y AB: 0.65 mm) y de los sépalos externos (LSE: 2.61 mm, ASE: 0.65 mm, LASE: 0.31 mm, LLSE: 2.29 mm y ISE: 4.24) (Cuadro 2). Los sépalos internos fueron de menor tamaño en muestras de México (LSI: 1.77 mm y LASI 0.10 mm). En contraste, el LLSI presentó un mayor largo en ejemplares colectados en México (1.68 mm); el ISI fue mayor en ejemplares de Guatemala (4.20) (Cuadro 2). El estigma fue más largo en ejemplares de Guatemala (0.82 mm); el valor del largo del utrículo fue similar en ejemplares colectados en Guatemala y México, mientras que el ancho fue mayor en los colectados en México (1.32 mm), la RLAU (1.21) fue mayor en ejemplares colectados en Guatemala (Cuadro 2). Las semillas de Guatemala fueron de mayor tamaño (LS: 1.13 mm y AS: 1.09), con respecto a las colectadas en México Cuadro 2). Los caracteres generados en relación con el largo bráctea: sépalo externo (RLBSE: 1.45) y bráctea: utrículo (RLBU: 1.76) fueron mayores en ejemplares procedentes de Guatemala y los ejemplares de México presentaron una mayor relación entre el utrículo: sépalo externo (RLUSE: 0.95) (Cuadro 2).
En Amaranthus hybridus, el coeficiente de variación señaló altas diferencias entre los ejemplares de ambas procedencias. La menor variación fue de 13.08% para el LS, y la mayor de 65.79% para LASE. Entre ejemplares de Guatemala el caracter LASI fue el de mayor variación (63.37%), el menos variante fue el LS (12.97%) (Cuadro 2). Para México la mayor variación se observó en LE (92.54%) y la menor en RLUSE (11.51%) (Cuadro 2).
La variación revelada en A. cruentus en los tres primeros componentes principales (CP) fue de 58%. En el CP1 (28%) fueron asociados a caracteres del sépalo externo (LSE, LASE, ISE y LLSE), sépalo interno (LSI), y la relación del largo de la bráctea:utrículo (RLBU); al CP2 (17%) se asociaron caracteres relacionados a la semilla (LS y AS), utrículo (AU) y largo del estigma (LE); y al CP3 (13%) caracteres relacionados al sépalo interno (ISI e ASI) y bráctea (AB) (Cuadro 4).
Cuadro 4: Detalles del análisis de componentes principales (CP) realizados en Amaranthus cruentus L. y A. hybridus L., señalando para los tres primeros componentes, su variación y los caracteres morfológicos que intervienen en ella.
| Descripción | A. cruentus L. | Descripción | A. hybridus L. | ||||
| CP1 | CP2 | CP3 | CP1 | CP2 | CP3 | ||
| Valores propios | 6 | 4 | 3 | Valores propios | 7 | 3 | 2 |
| Variación individual (%) | 28 | 17 | 13 | Variación individual (%) | 33 | 15 | 10 |
| Variación acumulada (%) | 28 | 45 | 58 | Variación acumulada (%) | 33 | 48 | 58 |
| Largo sépalo externo (LSE) | -0.777 | 0.393 | -0.263 | Largo sépalo interno (LSI) | 0.913 | -0.189 | -0.012 |
| Largo ápice sépalo externo (LASE) | -0.770 | 0.330 | -0.109 | Largo sépalo externo (LSE) | 0.883 | 0.204 | 0.153 |
| Índice sépalo externo (ISE) | -0.740 | 0.119 | -0.086 | Largo lámina del sépalo interno (LLSI) | 0.883 | -0.254 | 0.029 |
| Relación largo bráctea utrículo (RLBU) | -0.705 | 0.021 | 0.165 | Largo lámina del sépalo externo (LLSE) | 0.865 | 0.078 | 0.182 |
| Largo sépalo interno (LSI) | -0.683 | 0.254 | 0.242 | Relación largo bráctea sépalo externo (RLBSE) | -0.687 | 0.098 | -0.439 |
| Largo lámina del sépalo externo (LLSE) | -0.659 | 0.361 | -0.292 | Índice sépalo interno (ISI) | 0.627 | 0.435 | -0.324 |
| Largo semilla (LS) | 0.340 | 0.845 | 0.221 | Largo semilla (LS) | 0.624 | -0.370 | -0.341 |
| Ancho semilla (AS) | 0.360 | 0.745 | 0.192 | Índice sépalo externo (ISE) | 0.378 | 0.768 | 0.249 |
| Ancho utrículo (AU) | 0.644 | 0.616 | 0.266 | Ancho sépalo externo (ASE) | 0.307 | -0.709 | -0.199 |
| Largo estigma (LE) | 0.010 | 0.527 | -0.088 | Ancho sépalo interno (ASI) | 0.002 | -0.699 | 0.368 |
| Índice sépalo interno (ISI) | -0.185 | -0.318 | 0.786 | Ancho utrículo (AU) | 0.302 | -0.163 | -0.722 |
| Ancho sépalo interno (ASI) | -0.099 | 0.485 | -0.726 | Relación largo ancho del utrículo (RLAU) | 0.143 | -0.297 | 0.498 |
| Ancho de bráctea (AB) | 0.460 | 0.026 | -0.633 | ||||
La variación revelada en A. hybridus en los tres primeros CP fue de 58%. El CP1 (33%) fue asociado a caracteres del sépalo externo (LSE y LLSE), sépalo interno (LSI, LLSI e ISI), semilla (LS) y la relación entre el largo de bráctea:sépalo externo (RLBSE); el CP2 (15%) se asoció al sépalo externo (ASE e ISE), y sépalo interno (ASI); el CP3 (10%) al utrículo (AU) y la relación del largo:ancho del utrículo (RLAU) (Cuadro 4).
Tanto en A. cruentus como en A. hybridus algunas estructuras florales resultaron relevantes, ya que ambas especies compartieron estructuras florales que revelarón alta variación como ASI, AU, ISE, ISI, LLSE, LS, LSE y LSI (Cuadro 4).
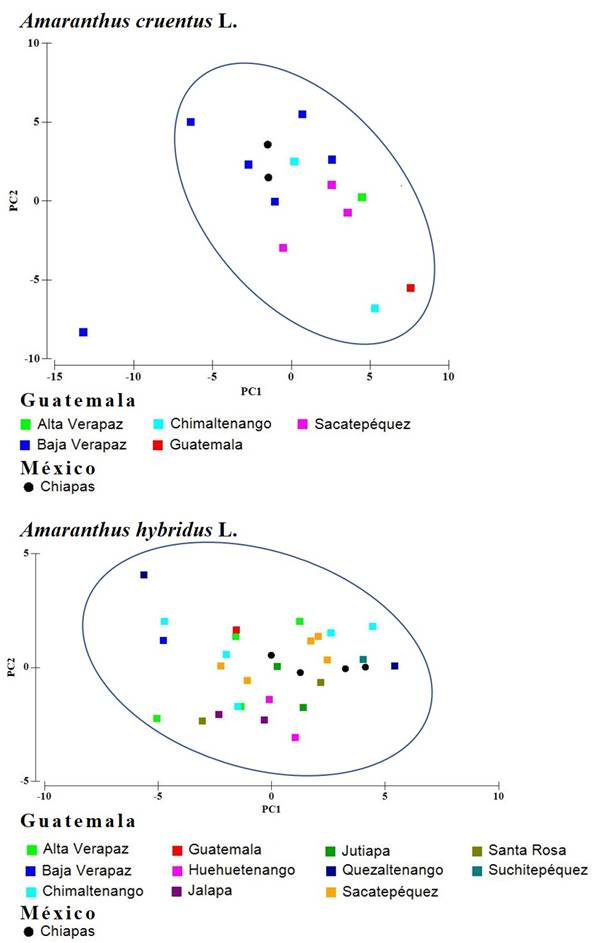
El análisis de ordenamiento para los CP 1 y 2, realizados en A. cruentus y A. hybridus, mostraron un solo grupo en ambas especies sin que la procedencia de colecta fuera un factor de agrupamiento. Entre los ejemplares de A. cruentus se separó del grupo uno de los colectados en Baja Verapaz (Fig. 1).
Discusión
Actualización de la distribución y frecuencia de A. cruentus y A. hybridus en la zona de estudio
Basado en los ejemplares de A. cruentus procedentes de la República Méxicana y en los resultados de este estudio, se puede afirmar que esta especie posee una mayor distribución con respecto a lo previamente reportado para el sureste del país, donde fue señalada su baja o limitada presencia, ya que básicamente se reportó solo de Chiapas (Espitia-Rangel et al., 2010a; Fig. 2). En este trabajo se evidencia una mayor distribución, se amplia su extensión territorial por el sureste mexicano, específicamente por los estados de Tabasco, Campeche y Yucatán (Fig. 2), pertenecientes a la Provincia Biótica Península de Yucatán (PBPY), misma que abarca la sección sureste de México (Campeche, Quintana Roo, Yucatán y porciones de Chiapas y Tabasco), el norte de Belice y el departamento guatemalteco de El Petén.
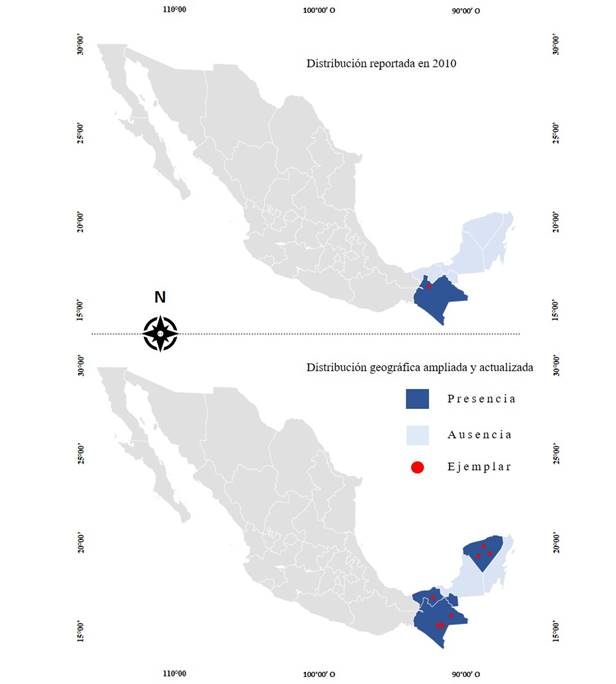
Figura 2: Nuevo reporte sobre la distribución de Amaranthus cruentus L. respecto a su distribución en 2010 para el sureste de México (Campeche, Chiapas, Tabasco, Quintana Roo y Yucatán).
No existen registros previos del cultivo comercial de especies de grano de amaranto, partiendo de los reportes del Sistema de Inovación Agroalimentaria y Pesquera (SIAP, 2018) de la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). El estudio etnobotánico realizado por Solís-Fernández (2015) señaló un reducido uso tradicional de A. cruentus en la Península de Yucatán, lo que podría ser uno de los factores limitantes de su presencia.
Los ejemplares de herbario incluidos en este estudio son la base para el reporte de la distribución y diversidad morfológica de A. cruentus en el área Maya. Estos contienen ejemplares silvestres, que históricamente se originaron por procesos de domesticado, y durante la prohibición de su cultivo en la colonia española se volvieron ferales, escaparon de cultivo o han sido conservados por núcleos familiares como herencia cultural. Por lo tanto, los resultados sobre su distribución nos llevan a suponer un posible vínculo derivado de procesos de domesticación, mismo que podría estar asociado a una limitada área de distribución por depender de circunstancias antropogénicas, en comparación con la distribución geográfica más amplia observada en su pariente silvestre, A. hybridus. Si bien su distribución es más amplia, al comparar ésta con reportes de A. cruentus en Chiapas, es posible señalar una abundancia mayor de A. cruentus en Yucatán y Chiapas, estados que presentan un área de dispersión y abundancia importante para esta especie en el sureste de México.
Espitia-Rangel et al. (2010a, b, c) han señalado la diversidad de A. cruentus en la República mexicana. Para América, Adhikary y Pratt (2015) reportaron el análisis de caracteres morfológicos asociados a flores pistiladas y estaminadas mediante técnicas multivariadas usando componentes principales en especies del complejo Amaranthus hybridus y ejemplares de A. cruentus procedentes de México (Sonora y Veracruz), y de A. hybridus de Estados Unidos de América (Indiana y Ohio), México (Puebla y Oaxaca), Guatemala (Huehuetenango y Sacatepéquez), Colombia (Cundinamarca), Perú (Ancash y Apurímac) y Brasil (Goiás). Este último estudio reveló, en A. cruentus, que todos los ejemplares se ordenaron en un mismo grupo, observando una variación morfológica continua, una situación similar a lo revelado en el presente trabajo.
Diversidad morfológica entre especies en la zona de estudio
El presente estudio proporciona nueva evidencia respecto a la diversidad en las especies A. cruentus y A. hybridus, al menos dentro de la región Maya, en la cual podemos observar diferencias morfológicas entre algunos de los ejemplares de A. cruentus estudiados, independientemente de su procedencia (Fig. 1).
Las diferencias son significativas en dimensiones del perianto con respecto al fruto y estigma de flores femeninas de A. cruentus procedentes de Guatemala con respecto a las presentes en México. El perianto tiene dimensiones mayores en las flores provenientes de Guatemala (p. ej. LB=2.37, LSE=2.22, LLSE=1.98; Cuadro 1), mientras que las partes reproductivas son mayores en las de México (p. ej. LE=0.67; Cuadro 1). Adicionalmente, este trabajo resalta como uno de los caracteres diagnósticos de relevancia para la identificación de taxa en el género a la relación entre el largo del sépalo externo y el fruto, la cual presenta muy poca variación para los ejemplares de México (p. ej. RLUSE CV=18.59, Cuadro 1). Este conjunto de rasgos permite considerar que probablemente las flores de Guatemala han sido menos manejadas en cultivo o domesticación a diferencia de las de México, debido a que los efectos más importantes en los síndromes de domesticación del género es la reducción en la talla de flores y brácteas (Sauer, 1967), así como el incremento en el tamaño del fruto. Además, resulta probable que el tamaño del estigma esté relacionado como una adaptación para la mayor captación de polen (Niklas y Buchmann, 1987; Swanson et al., 2004). Con base en esta posibilidad y sustentando la hipótesis del origen de A. cruentus en Guatemala, el gradiente de distribución observado (Fig. 1) puede ser resultado de la mezcla de formas silvestres, semidomesticadas o ferales que se encuentran en Guatemala, llevándose un intercambio genético entre estas formas. Esta suposición se basa en el reporte de Stetter et al. (2019), quienes reportaron, al analizar especies cultivadas de amarantos (A. caudatus, A. cruentus y A. hypocondriacus), sus ancestros silvestres (A. hybridus y A. quitensis) y organismos semidomesticados, la reducción de talla en caracteres morfológicos asociada al proceso de domesticación. La reducción de talla en este trabajo se observó principalmente en los ejemplares colectados en México, por lo que los procesos de domesticación podrían ser más notorios en está región.
En lo que respecta a A. hybridus que es la forma silvestre y maleza, se observaron gradientes de diferenciación morfológica (Cuadro 2) en tamaño de la mayoría de las estructuras florales, siendo más evidentes en los especímenes provenientes de Guatemala con respecto a los de México, mientras que el fruto de ambos es similar en tamaño; lo que permite observar un gradiente de diferenciación en la especie (Fig. 1). En este sentido, tan solo se reportan los rangos de variación morfológica para la especie contemplando material de la región Maya.
La alta variación morfológica revelada en A. cruentus y A. hybridus puede ser asociada a una alta diferenciación intraespecífica. Este efecto es reportado en centros de origen y diversificación de las principales especies cultivadas como arroz, maíz, papa y trigo (Boege, 2009). El reporte realizado por Solís-Fernández et al. (2020), sobre la presencia de morfotipos (Amaranthus aff. hybridus-Morphotype 1 y Amaranthus aff. hybridus-Morphotype 2) e híbridos (Amaranthus × wallichii y Amaranthus hybridus × Amaranthus cruentus) en la región de colecta, es también una evidencia indirecta de variación morfológica intraespecífica.
Con respecto a la diversidad de A. cruentus, Das (2012) reportó un estudio realizado a tres especies de grano del género Amaranthus: dos accesiones de A. hypochondriacus, tres de A. caudatus y cuatro de A. cruentus, así como a especies arvenses del mismo género (A. dubius Mart. ex Thell., A. graecizans L., A. hybridus, A. retroflexus L., A. spinosus L. y A. viridis L.), analizando la morfología (hojas, flores masculinas y femeninas, semillas y frutos) y bioquímica (compuestos fenólicos y la variación de isoenzimas de la fosfatasa ácida) de las especies incluidas. Como resultado, Das (2012) determinó que el conjunto de caracteres morfológicos y bioquímicos analizados permitieron observar diferencias entre las especies de grano y los arvenses, además de presentar la capacidad de distinguir a A. cruentus de las otras dos especies de grano por las características de la inflorescencia (estructura apical, color, grado de ramificación), la semilla (color y ornamentación) y a nivel isoenzimático (perfil). Lo encontrado por Das (2012) se complementa con los resultados obtenidos en el presente estudio sobre la amplia diversidad de A. cruentus con respecto a caracteres morfológicos asociados a estructuras florales y esta podría expresarse en características de importancia como la concentración de β-lainas y proteínas, las cuales son actualmente muy apreciadas en el ramo industrial y de gran valor en la alimentación humana y animal.
Nuestros resultados nos permiten sentar precedentes en la región sureste de México y Guatemala sobre la alta variación morfológica de A. cruentus, la cual podría ser una fuente genética de importancia. Espitia-Rangel et al. (2010 b) también señalan como importante la región biogeográfica de la Península de Yucatán, aunque discrepamos sobre la observación realizada para esta zona al ser señalada como centro secundario de colecta. Consideramos que se evidencia gran diferenciación morfológica entre ejemplares de esta región, los cuales por distancia geográfica podrían incluir caracteres o alelos diferentes a los encontrados en las áreas con mayor reporte de abundancia como lo ha sido el centro de México, incrementando el acervo genético de A. cruentus.
Conclusiones
Las flores en el género Amaranthus son estructuras básicas para la identificación taxonómica de las especies del grupo. Por lo tanto, la variación morfológica de estas estructuras debe ser baja, para permitir que se conserve la identidad de cada taxon, y en el caso de presentarse gran variación, como hemos observado en este trabajo, suponemos entonces que debe ser un indicador de alta diversidad genética.
Los resultados aquí obtenidos brindan nueva información relacionada a la diversidad genética y distribución geográfica de A. cruentus y A. hybridus en América, además de presentar evidencia que contribuye a apoyar la hipótesis sobre el centro de domesticación de A. cruentus en Guatemala y su área de diversificación en México. Con base en la caracterización morfológica de flores pistiladas, podemos afirmar alta diversidad genética en A. cruentus, incluso mayor a lo observado en A. hybridus, situación poco común ya que esta última especie es considerada silvestre. Tomando en cuenta que A. cruentus presentó procesos de domesticación relacionados con su importancia para culturas prehispánicas, es posible que aún se presente flujo de genes entre especies, misma que se refleja en la variación morfológica observada. Además, los resultados observados en este trabajo permiten hipotetizar que el germoplasma de A. cruentus distribuido en el área Maya podría ser un nuevo acervo genético, al menos diferente al estudiado en la zona centro y pacífico de México, información que contribuye con bases para estudios posteriores de ambas especies para esta región geográfica. Esta hipótesis tambien se sustenta con base en los reportes de distribución y frecuencia presentados en este estudio, que proporcionan evidencia respecto a una distribución extendida de A. cruentus dentro de México, misma que se amplía hacia del sureste de México y sobre la cual existen escasos reportes. El reporte de distribución extendida evidencia la importancia de contribuir con el incremento del acervo botánico y la conservación de los herbarios como centros de sistematización, representación y conservación de la biodiversidad, al permitir accesar y analizar información histórica y regional. La representatividad de la distribución y frecuencia de una especie con base en registros de herbario está supeditada a la cantidad de ejemplares y la representatividad de su acervo. Los herbarios son depositarios históricos de la biodiversidad de una región o país (Martínez-Castillo y Yánez-Espinosa, 2011), y, por ello, pueden ser tomados como una herramienta básica y fiable en la determinación de distribución y abundancia, tomando mayor valor al estudiar especies representativas del área biogeográfica donde estos se localizan, como es el caso de ambas especies de Amaranthus incluidas en el presente estudio. Partiendo de la idea anterior, este trabajo documenta el acervo botánico de A. cruentus en el herbario CICY, aumentando el número de ejemplares a nivel nacional y señalando la presencia de esta especie en la Península de Yucatán, complementando estudios como el realizado por Espitia-Rangel et al. (2010a). En comparación, A. hybridus se distribuye prácticamente en todo el territorio nacional, aunque el Eje Volcánico Transversal, el Golfo de México y el Altiplano mexicano son las zonas de mayor distribución.
Con base en lo anterior se sugiere ampliar estudios sobre la diversidad y abundancia de A. cruentus en la región sureste de México, extendiendo estos estudios hacia la Península de Yucatán, realizando análisis a nivel molecular, químico, bromatológico y agronómico de A. cruentus con la finalidad de revelar su potencial genético, agronómico e industrial.
Se reporta en A. hybridus una distribución y frecuencia más amplia que las de A. cruentus, quizá debido a que A. hybridus es una especie arvense colonizadora, invasiva, tolerante a ambientes adversos y de amplia distribución, asociada a diferentes cultivos a lo largo del mundo (Das, 2016a, b). En contraste, A. cruentus es una especie domesticada que, a pesar de haber iniciado este proceso en épocas prehispánicas (Hernández y Herrerías, 1998; Becerra, 2000), su uso como cereal de grano y como verdura fue limitado durante la colonia por razones religiosas de los españoles, al menos en México.

