











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los estudios de biología floral en comunidades de plantas tropicales sujetas a clima estacional han revelado algunos patrones respecto a las características de las flores; por ejemplo, dominancia de unidades de polinización individuales, dispuestas en plano horizontal, pero inclinadas alrededor de 45° hacia arriba, con forma de plato a tazón, simetría radial, corolas blancas o amarillas y néctar como recompensa principal (Percival, 1974; Ramírez, 1989; Ramírez et al., 1990; Ramírez y Brito, 1992; Oliveira y Gibbs, 2000; Machado y Lopes, 2004; Barrios y Ramírez, 2016). Asimismo, se ha indicado la prevalencia de antesis diurna y longevidad igual o menor a un día (Percival, 1974; Frankie et al., 1983; Ramírez et al., 1990; Ramírez y Brito, 1992; Oliveira y Gibbs, 2000; Barrios y Ramírez, 2016). Dichos atributos reflejan el papel que tienen los insectos de actividad diurna, específicamente las abejas, moscas, mariposas y escarabajos, en orden decreciente de importancia, en la polinización de las especies de plantas de estas comunidades (Percival, 1974; Heithaus, 1979; Frankie et al., 1983; Ramírez, 1989; Ramírez y Brito, 1992; Oliveira y Gibbs, 2000; Lemus-Jiménez y Ramírez, 2003; Machado y Lopes, 2004; Ramírez, 2004; Barrios, 2017).
Desde otra perspectiva, el predominio de un conjunto de caracteres florales sugiere que existe un nivel alto de solapamiento en el uso de polinizadores; así lo indica, de hecho, la estructura anidada que muestran las interacciones planta-polinizador bajo el enfoque de redes complejas (Bascompte et al., 2003; Jordano et al., 2009). Las redes anidadas presentan un núcleo de especies de plantas y de polinizadores generalistas cuyos patrones de interacción se superponen en gran medida; las especies generalistas interactúan entre ellas, pero a la vez lo hacen con especies más especializadas (Bascompte et al., 2003; Jordano et al., 2009). Esta configuración, altamente cohesiva, mantiene la robustez del sistema frente a perturbaciones extrínsecas (Bascompte et al., 2003; Jordano et al., 2009). No obstante, podría incrementar otros procesos en lo interno de la red, como la pérdida de polen, el bloqueo estigmático con polen heteroespecífico y/o la formación de híbridos estériles o poco vigorosos, factores que afectan negativamente los niveles de adecuación de las especies de plantas (Vidal y Ramírez, 2005; Gómez et al., 2007; Gómez y Perfectti, 2009).
Los efectos negativos del solapamiento en el uso de los polinizadores han sido señalados como las causas supremas del desfase de los períodos de floración entre las especies de plantas que coinciden en sus características florales y/o síndromes de polinización (Stiles, 1978; Waser, 1978, 1983; Rathcke y Lacey, 1985; Rathcke, 1988; Murali y Sukumar, 1994; Henderson et al., 2000; Lobo et al., 2003). Aunque también se han reportado casos de especies con flores aparentemente similares que son polinizadas por distintas clases de agentes, debido a variaciones más o menos sutiles en las señales visuales (colores, guías florales), la composición química de los aromas y el tipo, cantidad y/o calidad de las recompensas ofertadas a los polinizadores (Johnson et al., 2005; Van der Niet et al., 2010; Peter y Johnson, 2014). Esto último, en particular, ha puesto en tela de juicio el poder predictivo de los síndromes de polinización y subrayado la mayor importancia de las observaciones directas de los polinizadores frente a las predicciones basadas en las características de las flores (Ollerton et al., 2009).
El interés particular de este trabajo está centrado en caracterizar la biología floral de un bosque tropical estacionalmente inundable con el propósito de explorar la relación entre atributos florales, examinar la diversidad de síndromes de polinización y evaluar la efectividad de este concepto para predecir las clases de agentes polinizadores. Además, se analizó el solapamiento en la fenología de floración de las especies con síndromes de polinización y/o clases de polinizadores similares en búsqueda de un patrón segregado. Se plantearon las preguntas siguientes: ¿Cuáles son las características florales predominantes en esta comunidad?, ¿Qué caracteres se encuentran preferentemente asociados?, ¿Existe correspondencia entre los polinizadores observados y los predichos por los síndromes de polinización?, ¿Los períodos de floración de las especies con síndromes de polinización y/o clases de polinizadores comunes se encuentran temporalmente desfasados?
Materiales y Métodos
Área de estudio
Este estudio fue realizado en un bosque seco ralo (ca. 81 ha) ubicado en la planicie de inundación del río Mene, costa oriental del lago de Maracaibo, Venezuela (10º26'46''-10°27'28.8''N, 71º26'44.16''-71°27'4''W) (Barrios y Ramírez, 2020). El clima de la región es semiárido (BSh) (Peel et al., 2007), los datos de la estación climatológica más cercana (ca. 1 km del área de estudio) indican que la precipitación media anual es de 570 mm, la evapotranspiración media anual de 2431 mm y la temperatura media mensual de 29 ºC. Las precipitaciones se distribuyen entre abril y noviembre, período en el que el bosque permanece anegado, o al menos con suelo saturado de agua, debido al desborde del río Mene. La estación seca se extiende de diciembre a marzo y durante este período la mayor parte del bosque presenta suelo seco y más de 75% de las especies arbóreas pierden sus hojas (Barrios, 2017; Barrios y Ramírez, 2020).
La vegetación presenta dos estratos: uno superior, discontinuo, de 8-20 m de alto, en el que predominan Coccoloba caracasana Meisn., Copernicia tectorum (Kunth) Mart., Cordia dentata Poir., Enneatypus ramiflorus (C.A. Mey.) Roberty & Vautier, Geoffroea spinosa Jacq., Guazuma ulmifolia Lam., Morisonia hastata (Jacq.) Christenh. & Byng y Pithecellobium lanceolatum (Humb. & Bonpl. ex Willd.) Benth. y uno inferior, de hasta 3 m, dominado por Hecatostemon completus (Jacq.) Sleumer y Rosenbergiodendron formosum (Jacq.) Fagerl. var. formosum. Por otra parte, en áreas anegadas y con baja densidad de árboles prevalecen hidrófitas y helófitas, entre ellas Cyperus articulatus L., Eleocharis mutata (L.) Roem. & Schult., Typha domingensis Pers., Nymphaea spp., Pistia stratiotes L. y varias especies de lemnoides (Barrios, 2017; Barrios y Ramírez, 2020).
Muestreo y clasificación de las especies
Entre los años 2006 y 2013 se realizaron visitas al área de estudio, mensuales (4-5 días de duración) durante los años 2009-2011, y más esporádicas el resto de los años. Las 48 especies de plantas incluidas en este trabajo (Cuadro 1, Fig. 1) fueron observadas al menos una vez en fenofase de floración y representan 41% de las especies de angiospermas que han sido inventariadas en el lugar (Y. Barrios, datos no publicados). Las determinaciones se realizaron con base en literatura especializada (Velásquez, 1994), revisiones en los herbarios HERZU y HMBLUZ (no indizado) de la Universidad del Zulia, así como en VEN de la Universidad Central de Venezuela (acrónimos según Thiers, 2020); también se realizaron consultas a los especialistas locales. Los ejemplares testigos fueron depositados en los herbarios HERZU, HMBLUZ y MERF, este último de la Universidad de Los Andes (acrónimo según Thiers, 2020). La nomenclatura fue actualizada de acuerdo con el sistema Plants of the World Online (POWO, 2019), donde puede consultarse la lista completa de sinónimos.
Cuadro 1 Especies de plantas estudiadas en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Para cada especie se señala la familia, forma de vida y fenología de floración. En el caso de las plantas de humedales, también se indica si son helófitas o hidrófitas. I: enero, II: febrero, III: marzo, IV: abril, V: mayo, VI: junio, VII: julio, VIII: agosto, IX: septiembre, X: octubre, XI: noviembre, XII: diciembre.
| Familia/Especie | Forma de vida | Fenología de floración |
|---|---|---|
| Acanthaceae | ||
| Ruellia paniculata L. | Sufrútice | XII-IV |
| Alismataceae | ||
| Aquarius floribundus (Seub.) Christenh. & Byng | Hierba (helófita) | II, VI, VIII, XI, XII |
| Apocynaceae | ||
| Cryptostegia grandiflora Roxb. ex R.Br. | Trepadora | X, XI |
| Araceae | ||
| Lemna aequinoctialis Welw. | Hierba (hidrófita) | IV |
| Pistia stratiotes L. | Hierba (hidrófita) | II-IV |
| Arecaceae | ||
| Copernicia tectorum (Kunth) Mart. | Árbol | I-XI |
| Asteraceae | ||
| Spilanthes urens Jacq. | Hierba | II-IV, VI-X |
| Bignoniaceae | ||
| Dolichandra unguis-cati (L.) L.G. Lohmann | Trepadora | V |
| Boraginaceae | ||
| Cordia dentata Poir. | Árbol | V-VIII, X |
| Bromeliaceae | ||
| Tillandsia flexuosa Sw. | Epífita | V-VII |
| Capparaceae | ||
| Morisonia americana L. | Árbol | III |
| Morisonia hastata (Jacq.) Christenh. & Byng | Árbol | III-VI, IX-XII |
| Morisonia odoratissima (Jacq.) Christenh. & Byng | Árbol | V |
| Convolvulaceae | ||
| Ipomoea carnea Jacq. | Arbusto | XII-IV |
| Jacquemontia pentanthos (Jacq.) G.Don | Trepadora | XI-I |
| Cyperaceae | ||
| Cyperus articulatus L. | Hierba (helófita) | II, IV-VIII, X-XI |
| Cyperus ligularis L. | Hierba (helófita) | VI |
| Cyperus luzulae (L.) Retz. | Hierba (helófita) | VI, X |
| Cyperus odoratus L. | Hierba (helófita) | II, III |
| Eleocharis mutata (L.) Roem. & Schult. | Hierba (helófita) | V-VIII, X |
| Fabaceae | ||
| Dalbergia ecastaphyllum (L.) Taub. | Arbusto | II, VIII |
| Geoffroea spinosa Jacq. | Árbol | II-VI |
| Libidibia coriaria (Jacq.) Schltdl. | Árbol | VII |
| Neptunia plena (L.) Benth. | Sufrútice | II, VI, X, XI |
| Pithecellobium lanceolatum (Humb. & Bonpl. ex Willd.) Benth. | Árbol | V-XI |
| Prosopis juliflora (Sw.) DC. | Árbol | III |
| Senna bicapsularis (L.) Roxb. | Arbusto | IX-II |
| Sesbania exasperata Kunth | Arbusto (helófito) | II, IX-XI |
| Vachellia macracantha (Humb. & Bonpl. ex Willd.) Seigler & Ebinger | Árbol | V, VII,VIII, IX |
| Hydrocharitaceae | ||
| Hydrocharis laevigata (Humb. & Bonpl. ex Willd.) Byng & Christenh. | Hierba (hidrófita) | II |
| Lamiaceae | ||
| Vitex compressa Turcz. | Árbol | V |
| Lythraceae | ||
| Ammannia auriculata Willd. | Hierba (helófita) | IX-I |
| Malvaceae | ||
| Guazuma ulmifolia Lam. | Árbol | IX-V |
| Waltheria indica L. | Arbusto | I,XI |
| Menispermaceae | ||
| Odontocarya hastata Barneby | Trepadora | VIII |
| Nymphaeaceae | ||
| Nymphaea amazonum Mart. & Zucc. | Hierba (hidrófita) | VII-II |
| Nymphaea pulchella DC. | Hierba (hidrófita) | I-XII |
| Onagraceae | ||
| Ludwigia erecta (L.) Hara | Sufrútice (helófito) | II,V-VII, XI, XII |
| Ludwigia leptocarpa (Nutt.) Hara | Sufrútice (helófito) | III-VII |
| Poaceae | ||
| Sporobolus virginicus (L.) Kunth | Hierba | VI, X |
| Polygonaceae | ||
| Antigonon leptopus Hook. & Arn. | Trepadora | IX |
| Coccoloba caracasana Meisn. | Árbol | II-IV, X |
| Enneatypus ramiflorus (C.A.Mey.) Roberty & Vautier | Árbol | XII |
| Rubiaceae | ||
| Rosenbergiodendron formosum (Jacq.) Fagerl. var. formosum | Arbusto | VII, VIII |
| Salicaceae | ||
| Hecatostemon completus (Jacq.) Sleumer | Arbusto | II-IV, VI, VII, IX-XII |
| Sapindaceae | ||
| Paullinia cururu L. | Trepadora | VI |
| Typhaceae | ||
| Typha domingensis Pers. | Hierba (helófita) | X-VIII |
| Violaceae | ||
| Calyptrion carthagenense (H. Karst.) Paula-Souza | Trepadora | VIII |

Fotografías: Y. Barrios (A, B, C, D, E, I, K), J.R. Grande (F, G, H, L, M); J. Delgado (J).
Figura 1 Algunas de las especies de plantas estudiadas en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela: A. Nymphaea amazonum Mart. & Zucc., B. N. pulchella DC. (Nymphaeaceae); C. Pistia stratiotes L. (Araceae); D. Aquarius floribundus (Seub.) Christenh. & Byng (Alismataceae); E. Cyperus ligularis L. (Cyperaceae); F. Sesbania exasperata Kunth; G. Libidibia coriaria (Jacq.) Schltdl., H. Pithecellobium lanceolatum (Humb. & Bonpl. ex Willd.) Benth. (Fabaceae); I. Morisonia hastata (Jacq.) Christenh. & Byng; J. M. odoratissima (Jacq.) Christenh. & Byng (Capparaceae); K. Jacquemontia pentanthos (Jacq.) G. Don (Convolvulaceae); L. Cordia dentata Poir. (Boraginaceae); M. Antigonon leptopus Hook. & Arn (Polygonaceae).
Las especies fueron clasificadas con base en sus formas de vida como árboles, arbustos, trepadoras, sufrútices, hierbas y epífitas, manteniendo el criterio utilizado por Ramírez (1993). En el caso de las especies de plantas de humedales, se discriminó entre formas hidrófitas y helófitas siguiendo el criterio de Velásquez (1994). Se registró la información fenológica (analizada para 39 spp. en Barrios (2017)) y en cada especie se examinaron las características florales (resumidas para 25 spp. en Barrios y Ramírez (2016) con más detalles en Barrios (2017)) y las clases de agentes polinizadores (descritas para 22 spp. en Barrios (2017)) en condiciones naturales.
Biología floral
Se examinaron las siguientes características florales en un mínimo de diez individuos por especie:
Sistema sexual: se distinguió entre hermafroditismo (flores bisexuales únicamente), andromonoecia (flores bisexuales y masculinas en la misma planta), monoecia (flores unisexuales femeninas y masculinas en la misma planta) y dioecia (flores femeninas o masculinas por individuo).
Dicogamia: en las especies con flores bisexuales (hermafroditas y andromonoicas) e incluso en las especies con flores femeninas y masculinas agrupadas en inflorescencias densas (monoicas), se realizó el seguimiento de al menos diez flores (o inflorescencias), de diferentes individuos, desde el inicio de la antesis hasta la senescencia. De ese modo se precisó si la disponibilidad del polen y el período de receptividad estigmática coinciden (homogamia o adicogamia) o están temporalmente desfasados (dicogamia). En este último caso, las especies fueron clasificadas como protóginas si la fase femenina precede a la masculina y protandras si la fase masculina precede a la femenina (Faegri y van der Pijl, 1979).
Hercogamia: las especies fueron consideradas hercógamas cuando presentaron una separación espacial entre anteras y estigma(s) (Cruden, 1977; Faegri y van der Pijl, 1979). Se distinguieron los casos de hercogamia recíproca como la heterostilia y la enantiostilia. En el primero existen dos o más formas florales que difieren en la longitud de anteras y estigmas: los individuos con flores longistilas presentan estilos largos y estambres cortos; mientras que, en aquellos con flores brevistilas las anteras se encuentran por encima de los estigmas. En el caso de la enantiostilia, las formas florales se diferencian en que el estilo está desviado a la izquierda o a la derecha de los estambres, de manera que cada tipo de flor es la imagen especular de la otra (Endress, 1994).
Tipo de unidad de polinización (UP): las unidades de polinización fueron caracterizadas de acuerdo con la organización de las flores y el comportamiento de los polinizadores (Ramírez et al., 1990). En las especies anemófilas (Faegri y van der Pijl, 1979) solo se consideró la organización de las flores. Cuando las flores formaban grupos densos y presentaban antesis relativamente sincrónica, la inflorescencia fue considerada la UP. En este caso, los polinizadores visitan muchas flores simultáneamente sin moverse a otra inflorescencia (UP colectiva). En el caso opuesto (UP individual), las unidades de polinización fueron las flores solitarias o integrantes de inflorescencias laxas y/o asincrónicas en la antesis de las flores (Ramírez et al., 1990).
Forma, simetría y orientación de las flores: la forma floral fue descrita de acuerdo con los tipos morfológicos de Faegri y van der Pijl (1979), quienes discriminaron entre plato a tazón, cepillo, bandera, campana o embudo, garganta, tubo, trampa y flores inconspicuas. En cuanto a la simetría, se distinguió entre flores actinomorfas (simetría radial), zigomorfas (simetría bilateral) e irregulares (asimétricas). Con relación a la orientación de las flores, se describió el patrón principal distinguiendo entre flores orientadas verticalmente hacia arriba y hacia abajo. Las flores dispuestas horizontalmente se denotaron más precisamente de acuerdo con la variación angular (hacia abajo o hacia arriba) respecto a un plano horizontal (Ramírez et al., 1990).
Dimensiones de flores e inflorescencias: las medidas del diámetro externo y del largo floral fueron registradas en 10 flores de diferentes individuos por especie; en algunos casos este número fue mayor o menor dependiendo de la disponibilidad de individuos. Cuando las inflorescencias constituyeron la unidad de polinización también fueron medidas en sus extremos. El diámetro externo fue el promedio de dos medidas perpendiculares entre ápices opuestos de los pétalos. El largo de las flores tubulares representó la longitud floral. En el caso de las flores no tubulares, el largo floral representó la medida entre la base del receptáculo y la altura de los pétalos, estambres o estigma (Ramírez et al., 1990).
Atrayentes y recompensas florales: el color principal y secundario de las flores (guías florales, estambres llamativos, brácteas) fue establecido de acuerdo con las categorías propuestas por Ramírez et al. (1990), quienes distinguieron entre (1) Blanco, flores blancas y blanco amarillentas; (2) Rojo-rosado, flores rojas, rosadas y fucsia; (3) Pardo, flores marrones; (4) Morado, flores azules, morado claro e intenso; (5) Amarillo, flores amarillas y anaranjadas y (6) Verde. La presencia de aromas fue descrita con dos categorías de intensidad: suave vs intenso. La oferta de recompensas (néctar, polen, partes florales, etc.) fue determinada mediante observación directa en un mínimo de 10 flores de diferentes individuos por especie. La presencia de nectarios o glándulas de otra naturaleza fue examinada con lupa, y al igual que la oferta de polen y de partes florales, también con base en el comportamiento de los visitantes florales.
Antesis y longevidad floral: para determinar el horario de antesis (diurno, nocturno) y el tiempo de vida de la flor se marcaron diez yemas florales de diferentes individuos por especie; estas fueron observadas regularmente desde el inicio de la antesis hasta la senescencia del órgano.
Síndromes de polinización y clases de polinizadores bióticos
Los síndromes de polinización fueron establecidos de acuerdo con el criterio de Faegri y van der Pijl (1979). En este sentido, se distinguió principalmente entre especies anemófilas, cantaridófilas, falenófilas, melitófilas, miófilas, psicófilas y ornitófilas.
Para registrar los polinizadores de las especies zoófilas, se realizaron observaciones directas de los visitantes florales durante toda la antesis y cuando fue posible en diferentes momentos del período de floración. Cuando los visitantes fueron insectos, también se realizaron capturas con redes aéreas para su posterior medición en el laboratorio. Aquellas morfoespecies de visitantes que mostraron más de un registro; es decir, que fueron fieles a la interacción, y cuya forma, tamaño y/o comportamiento les permitieron hacer contacto con las estructuras sexuales de la flor durante la visita, fueron considerados polinizadores.
Dada la naturaleza del ensamble de polinizadores, se siguió el criterio de Ramírez (1989) para separar las clases de agentes por categoría de tamaños. En este sentido, se distinguió entre abejas pequeñas (≤7 mm), medianas (>7≤14 mm) y grandes (>14 mm), avispas pequeñas a medianas (≤20 mm) y grandes (>20 mm), moscas pequeñas (≤10 mm) y grandes (>10 mm), esfíngidos y colibríes. Cabe destacar; sin embargo, que muchas especies de plantas fueron polinizadas por más de un tipo de agente; por esta razón, el número total de especies de plantas por clase de polinizador fue superior al de las evaluadas.
Análisis estadísticos
La relación entre el tipo de unidad de polinización, forma, simetría, orientación y color de las flores, así como entre los síndromes de polinización y las clases de polinizadores, fue explorada mediante la aplicación de la prueba exacta de Fisher a cada par de variables categóricas. La variación de las dimensiones florales entre tipos de unidades de polinización, forma, simetría y color de las flores, fue analizada mediante una prueba t o un ANDEVA según el número de categorías en cada caso. Dado que los datos fueron heterocedásticos, así como heterogéneos en cuanto al tamaño (n) de cada categoría, se utilizaron las versiones de Welch para estas pruebas (Hammer et al., 2001). En ambos casos, los valores fueron previamente transformados por la fórmula √(X + 1) donde X representa el valor de la variable (Sokal y Rohlf, 1969). Las comparaciones múltiples dentro de cada ANDEVA fueron realizadas a posteriori mediante la prueba de Tukey-Kramer. Finalmente, se realizó un análisis de correspondencia (AC) para representar gráficamente los estados de cada atributo floral y las asociaciones entre ellos. Para efecto de todos los análisis, se utilizó el programa PAST v. 3.0 (Hammer et al., 2001) y el nivel de significancia mínimo establecido fue igual a 0.05.
Para examinar si la fenología de floración de las especies con síndromes de polinización y clases de polinizadores comunes es más segregada de lo esperado por efecto del azar, se contrastó el solapamiento promedio de cada clase con un modelo nulo de aleatoriedad. El algoritmo empleado para generar las matrices nulas (n=10000) fue ROSARIO (Castro-Arellano et al., 2010). El solapamiento fue calculado entre todos los pares de especies de cada clase de acuerdo con el índice simétrico de Pianka (Ojk) (Pianka, 1973). La posición del solapamiento promedio observado en la distribución de solapamiento promedio de las matrices nulas arrojó la probabilidad exacta de obtener índices menores de lo esperado por efecto del azar (α=0.05). La permutación de las matrices, la estimación de los índices y de los valores de probabilidad fue realizada con el programa “TimeOverlap” (Castro-Arellano et al., 2010).
Resultados
La forma de vida de las especies estudiadas se encuentra especificada en el Cuadro 1. En su mayoría se trató de hierbas (n=15 spp., 31.25%) y árboles (n=14 spp., 29.16%), seguidos de arbustos (n=7 spp., 14.58%), trepadoras (n=7 spp., 14.58%), sufrútices (n=4 spp., 8.33%) y epífitas (n=1 sp., 2.10%). La mayoría de las especies herbáceas (n=13 spp., 86.66%), dos de las sufruticosas y una de las arbustivas, fueron plantas de humedales (helófitas o hidrófitas).
Biología floral
El sistema sexual más frecuente fue el hermafroditismo (hermafroditas n=39 spp., 81.30%; dioicas n=4 spp., 8.33%; monoicas n=3 spp., 6.25%; andromonoicas n=2 spp., 4.16%). Las especies fueron principalmente adicógamas (adicogamia n=24 spp., 77.41%; protoginia n=6 spp., 19.35%; protandria n=1 sp., 3.22%) y no hercógamas (no hercógamas n=33 spp., 75%; hercógamas n=11 spp., 25%); no obstante, se observó un caso de heterostilia (Cordia dentata Poir.) y otro de enantiostilia (Senna bicapsularis (L.) Roxb.) (Apéndice).
Las flores fueron la unidad de polinización predominante (flor n=34 spp., 70.83%; inflorescencia n=14 spp., 29.16%). La forma floral más frecuente fue plato a tazón (n=14 spp., 28.57%), seguida de los tipos cepillo (n=9 spp., 18.36%) e inconspicuo (n=9 spp., 18.36%), y posteriormente de las formas bandera (n=5 spp., 10.20%), embudo (n=4 spp., 8.16%), tubo (n=4 spp., 8.16%), garganta (n=3 spp., 6.10%) y trampa (n=1 sp., 2%). Las flores fueron principalmente actinomorfas (n=29 spp., 60.41%); la zigomorfía (n=10 spp., 20.83%) y las formas irregulares (n=9 spp., 18.75%) estuvieron menos representadas. La principal orientación de las flores fue horizontal hacia arriba (n=29 spp., 60.41%), seguida de vertical hacia arriba (n=11 spp., 22.91%), horizontal hacia abajo (n=6 spp., 12.50%) y horizontal (n=2 spp., 4.16%) (Apéndice).
Las dimensiones de las flores estuvieron comprendidas entre 0.01-11.5 (
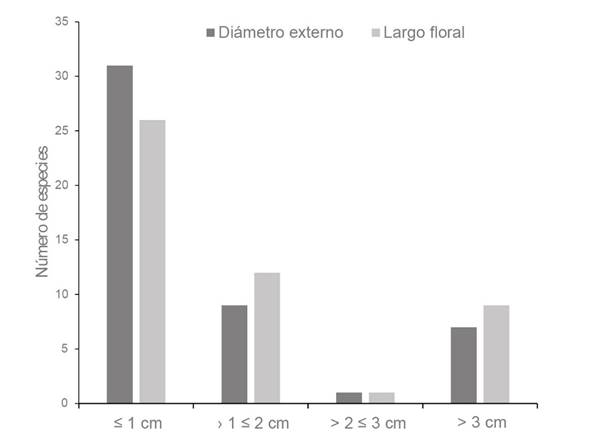
Figura 2 Clases de tamaños florales de las especies de plantas estudiadas en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela.
La mayoría de las especies de plantas ofrecen néctar como recompensa (n=33 spp., 86.84%); no obstante, en muchos de estos casos (n=21 spp., 63.63%), se observó de manera paralela la colecta y/o consumo de polen en las flores, por lo que se consideró a este último un recurso secundario. La oferta exclusiva de polen (n=4 spp., 10.52%) o en combinación con partes florales (n=1 sp., 2.60%) fue un tipo de recompensa poco frecuente. La antesis fue principalmente diurna (antesis diurna n=40 spp., 85.10%; antesis nocturna n=7 spp., 14.90%) y la longevidad floral corta, con una duración de un día (n=25 spp., 69.40%) o una noche (n=4 spp., 11.19%), aunque en Morisonia odoratissima, de antesis nocturna, las flores fueron visitadas hasta el mediodía siguiente a la antesis. De cualquier manera, las flores con longevidad mayor a un día, o a una noche, fueron poco comunes (n=7 spp., 19.40%) (Apéndice).
Síndromes de polinización y clases de polinizadores bióticos
La melitofilia fue el síndrome de polinización predominante (n=28 spp., 62.22%), seguida de anemofilia (n=8 spp., 17.77%), falenofilia (n=6 spp., 13.33%), cantaridofilia (n=1 sp., 2.22%), miofilia (n=1 sp., 2.22%) y ornitofilia (n=1 sp., 2.22%) (Apéndice).
Entre las especies de plantas zoófilas, la mayoría fue polinizada por abejas medianas (n=22 spp., 51.16%), seguidas en orden de importancia por abejas grandes (n=8 spp., 18.60%), avispas pequeñas a medianas (n=4 spp., 9.30%), moscas grandes (n=3 spp., 6.97%), abejas pequeñas (n=2 spp., 4.65%), colibríes (n=2 spp., 4.65%) y esfíngidos (n=2 spp., 4.65%) (Apéndice).
Relación entre el tipo de unidad de polinización (UP), forma, simetría, orientación y color de las flores
El tipo de UP estuvo asociado significativamente con la forma (Fisher p<0.0000), simetría (Fisher p<0.0000), orientación (Fisher p=0.0008) y color de las flores (Fisher p<0.0000). En este sentido, las unidades de polinización individuales estuvieron principalmente representadas por flores de tipo plato a tazón, actinomorfas, dispuestas horizontalmente (hacia arriba o hacia abajo) y de color blanco (Fig. 3). Las unidades de polinización colectivas, en cambio, estuvieron mayormente representadas por inflorescencias con flores inconspicuas, asimétricas o actinomorfas, dispuestas en posición vertical (hacia arriba) y de color pardo o verde (Fig. 3).
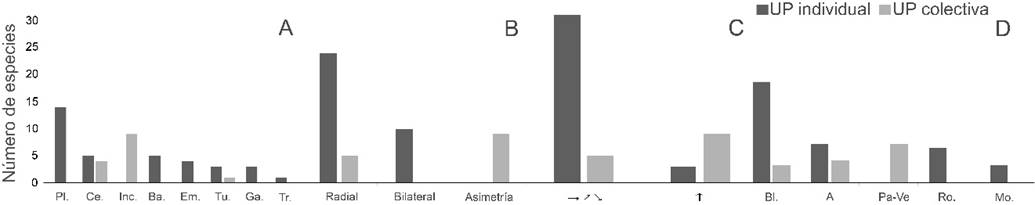
Figura 3 Distribución de frecuencia de los tipos de unidad de polinización (UP), individual y colectiva, entre categorías de forma (A), simetría (B), orientación espacial (C) y color de las flores (D) en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Forma floral (A): Ba.=bandera, Ce.=cepillo, Em.=embudo, Ga.=garganta, Inc.=inconspicua, Pl.=plato a tazón, Tu.=tubo, Tr.=trampa. Orientación espacial (C): horizontal incluyendo hacia arriba y hacia abajo (→, ↗, ↘), vertical hacia arriba (↑). Color (D): A=amarillo, Bl.=blanco, Mo.=morado, Pa-Ve=pardo o verde, Ro.=rojo-rosado.
La forma también estuvo asociada significativamente con la simetría (Fisher p<0.0000), orientación (Fisher p<0.0001) y color de las flores (Fisher p<0.0000). Las flores con formas de plato a tazón, cepillo, embudo y tubo fueron primordialmente actinomorfas; las flores con formas de garganta y bandera, zigomorfas, mientras que las flores inconspicuas, asimétricas (Fig. 4A). En estas últimas, prevaleció la orientación vertical, a diferencia de las formas restantes orientadas principalmente en posición horizontal (Fig. 4B). Por otra parte, las flores con formas de plato, cepillo y tubo fueron mayormente blancas; en las flores inconspicuas prevalecieron el pardo y el verde, mientras que en las formas restantes más de una categoría de color estuvo bien representada, por ejemplo, amarillo y blanco en bandera; rojo-rosado, blanco y morado en embudo; morado y amarillo en garganta (Fig. 4C).
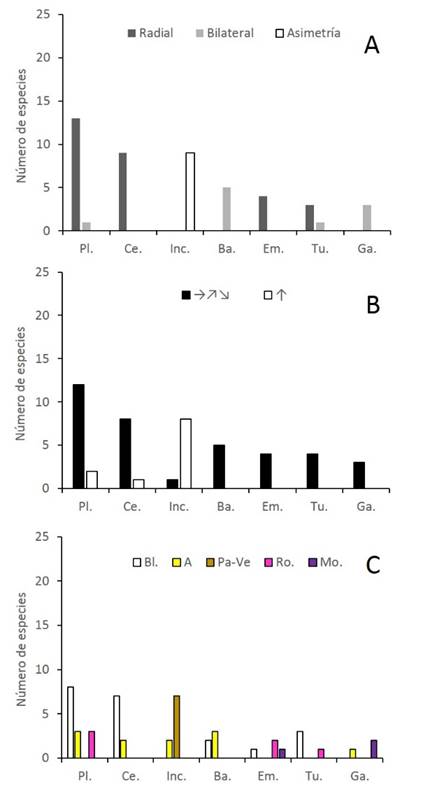
Figura 4 Distribución de frecuencia de los tipos de simetría (A), orientación espacial (B) y color de las flores (C) entre categorías de formas florales: Ba.=bandera, Ce.=cepillo, Em.=embudo, Ga.=garganta, Inc.=inconspicua, Pl.=plato a tazón, Tu.=tubo, en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Orientación espacial (B): horizontal incluyendo hacia arriba y hacia abajo (→, ↗, ↘), vertical hacia arriba (↑). Color (C): A=amarillo, Bl.=blanco, Mo.=morado, Pa-Ve=pardo o verde, Ro.=rojo-rosado.
En cuanto a la relación del color con la simetría (Fisher p<0.0000), el blanco y el rojo-rosado estuvieron principalmente asociados con la actinomorfía, el amarillo fue más frecuente en las flores simétricas (actinomorfas y zigomorfas) que en las asimétricas y el morado exclusivo de las flores simétricas. Los colores pardo y verde, en cambio, fueron exclusivos de flores asimétricas (Fig. 5A). El color y la orientación de las flores también estuvieron relacionados (Fisher p<0.0000); los colores rojo rosado y morado solo se encontraron en flores dispuestas horizontalmente, asimismo, el blanco y el amarillo fueron más frecuentes en este tipo de orientación, mientras que el pardo y el verde fueron exclusivos de flores orientadas verticalmente (Fig. 5B). Respecto a la relación entre la simetría y la orientación (Fisher p<0.0000), las flores actinomorfas y zigomorfas estuvieron principalmente orientadas en posición horizontal mientras que la orientación de las flores asimétricas fue mayormente vertical (Fig. 6).
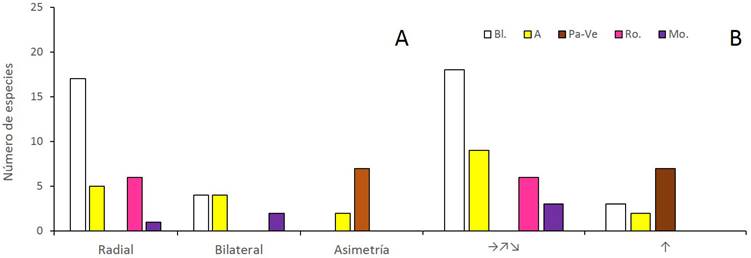
Figura 5 Distribución de frecuencia de los colores de las flores entre categorías de simetría (A) y orientación espacial (B) en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Color: A=amarillo, Bl.=blanco, Mo.=morado, Pa-Ve=pardo o verde, Ro.=rojo-rosado. Orientación espacial: horizontal incluyendo hacia arriba y hacia abajo (→, ↗, ↘), vertical hacia arriba (↑).
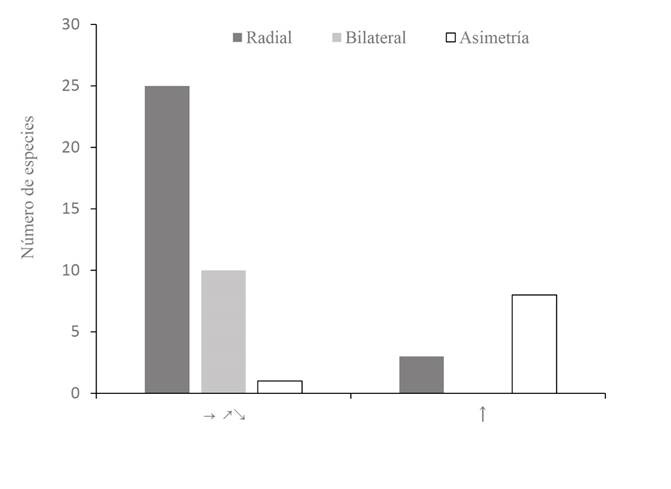
Figura 6 Distribución de frecuencia de los tipos de simetría (radial, bilateral, asimetría) de acuerdo con la orientación espacial de las flores en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Orientación espacial: horizontal incluyendo hacia arriba y hacia abajo (→, ↗, ↘), vertical hacia arriba (↑).
Variabilidad de las dimensiones florales entre tipos de unidad de polinización (UP), forma, simetría y color de las flores
Las dimensiones de las flores variaron entre tipos de UP, forma y simetría de las flores. En este sentido, el diámetro externo y el largo floral fueron menores en unidades de polinización colectivas que en las individuales. El largo de las flores inconspicuas fue menor que el de las flores con forma de embudo, garganta y tubo; asimismo, el largo de flores con forma de plato a tazón y bandera fue menor que el de flores con forma de garganta. Con relación a la simetría, las flores asimétricas mostraron menor diámetro externo que las flores actinomorfas y menor largo floral que las flores actinomorfas y zigomorfas respectivamente (Cuadro 2).
Cuadro 2 Comparación de las dimensiones florales (diámetro externo y largo) entre tipos de unidad de polinización (UP), de forma, simetría y color de las flores en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Para cada análisis se muestra el número de especies en cada categoría (n), la media (
| n |
|
|
|||
|---|---|---|---|---|---|
| Unidad de polinización | |||||
| Flor | 34 | 2.19±2.90 | t=5.12 | 2.22±2.29 | t=5.20 |
| Inflorescencia | 14 | 0.16±0.12 | (p<0.0000) | 0.40±0.31 | (p<0.0000) |
| Forma | |||||
| Plato a tazón | 14 | 2.20±3. | F(9.99 g.l.)=5.45 | 1.30±1.65 a | F(10.56g.l.)=7.49 |
| Cepillo | 9 | 1.43±1.99 | (p=0.0090)* | 1.44±1.45 | (p=0.0025) |
| Inconspicua | 9 | 0.11±0.08 | 0.29±0.16 bcd | ||
| Bandera | 5 | 0.88±0.52 | 1.04±0.28 e | ||
| Embudo | 4 | 3.47±2.71 | 3.32±2.57 b | ||
| Tubo | 4 | 1.7±2.66 | 3.7±3.32 d | ||
| Garganta | 3 | 2.36±1.65 | 4.73±3.51 ace | ||
| Simetría | |||||
| Radial | 29 | 2.19±3.14 a | F (18.63 g.l.) =15.32 | 1.95±2.15a | F(19.07 g.l.) =15.11 |
| Bilateral | 10 | 1.23±1.17 | (p=0.0001) | 2.21±2.46b | (p=0.0001) |
| Asimetría | 9 | 0.11±0.08 a | 0.29±0.16ab | ||
| Color | |||||
| Blanco | 21 | 2.26±3.43 | F(9.50 g.l.)=10.59 | 1.83±2.18 | F(9.57 g.l.)=7.94 |
| Amarillo | 11 | 0.97±1.19 | (p=0.0015)* | 1.51±2.42 | (p=0.0042)* |
| Pardo-verde | 7 | 0.10±0.04 | 0.28±0.10 | ||
| Rojo-rosado | 6 | 2.33±2.72 | 2.92±2.29 | ||
| Morado | 3 | 1.33±0.49 | 2.20±1.21 | ||
Análisis de correspondencia entre los caracteres florales
Los diferentes estados de los caracteres florales se distribuyen normalmente en el espacio multivariado, lo que refleja una elevada redundancia a nivel general. Algunos de estos caracteres; sin embargo, tienden a alejarse en grupo de la nube central de datos y permiten vislumbrar ciertos patrones específicos. Por ejemplo, la relación entre unidad de polinización colectiva (inflorescencia), forma inconspicua, asimetría, orientación vertical y color pardo-verde (anemofilia principalmente); la asociación entre antesis nocturna y aroma intenso (falenofilia y cantaridofilia); y la asociación de las formas bandera y garganta con la zigomorfía, así como con los colores amarillo y morado, respectivamente (melitofilia) (Fig. 7).
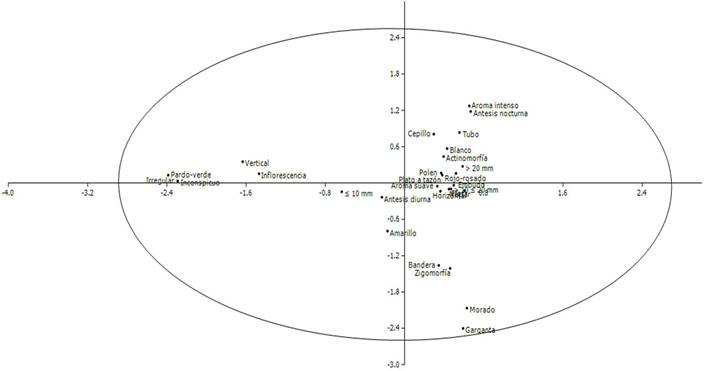
Figura 7 Análisis de correspondencia (AC) entre las características florales de las especies estudiadas en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. Características: tipo de unidad de polinización (flor, inflorescencia), forma (bandera, cepillo, embudo, garganta, inconspicua, plato a tazón, tubo), simetría (actinomorfa, asimetría, zigomorfa), orientación espacial (horizontal, vertical), dimensiones (≤10 mm, >10≤20 mm, >20 mm), color (amarillo, blanco, morado, pardo-verde, rojo-rosado), aroma (intenso, suave), recompensa (néctar, polen) y antesis (diurna, nocturna). Los dos primeros ejes del AC explican 40,70% de las asociaciones observadas.
Relación entre los síndromes de polinización y las clases de polinizadores
Los polinizadores observados estuvieron bastante acordes con lo esperado de acuerdo con los síndromes de polinización de las especies (Fisher p=0.0099). No obstante, en varios casos se observó más de una clase de agente polinizador e incluso clases que no se correspondían con lo esperado según el síndrome de las flores. En este sentido, las especies melitófilas fueron polinizadas principalmente por abejas, seguidas de avispas, moscas y colibríes; las especies falenófilas fueron polinizadas por esfíngidos y abejas, y la especie ornitófila por colibríes. La mayor discrepancia se observó en la especie miofila, cuyos únicos polinizadores registrados fueron avispas (Fig. 8).
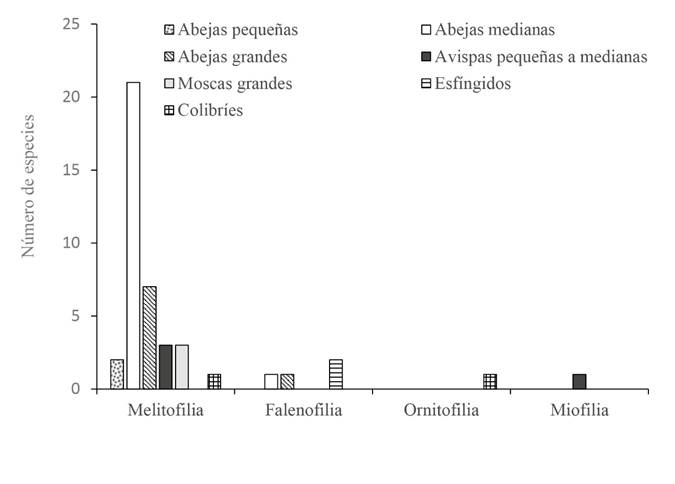
Solapamiento fenológico
La distribución temporal de los eventos de floración de las especies con síndromes de polinización y/o clases de polinizadores en común no difirió significativamente de los resultados del modelo nulo; es decir, la fenología no muestra un patrón más segregado de lo esperado por efecto del azar (Cuadro 3).
Cuadro 3 Solapamiento fenológico de las especies que comparten síndromes de polinización y/o clases de agentes polinizadores en un bosque inundable de la cuenca del lago de Maracaibo, Venezuela. En cada caso se señala el número de especies en cada categoría (n), el promedio del índice simétrico de Pianka (Ojk) y la probabilidad asociada con la hipótesis nula de aleatoriedad (p). * Los síndromes de cantaridofilia, miofilia y ornitofilia no se incluyen en el análisis por presentar n=1.
| Solapamiento fenológico | |||
|---|---|---|---|
| n | Ojk | (p) | |
| Síndrome de polinización* | |||
| Melitofilia | 28 | 0.26 | 0.83 |
| Anemofilia | 8 | 0.35 | 0.97 |
| Falenofilia | 6 | 0.19 | 0.63 |
| Clase de agente polinizador | |||
| Abejas medianas | 22 | 0.27 | 0.45 |
| Abejas grandes | 8 | 0.34 | 0.82 |
| Avispas pequeñas a medianas | 4 | 0.38 | 0.39 |
| Moscas grandes | 3 | 0.27 | 0.72 |
| Abejas pequeñas | 2 | 0.69 | 0.90 |
| Colibríes | 2 | 0.44 | 0.82 |
| Esfíngidos | 2 | 0.02 | 0.15 |
Discusión
Los atributos florales predominantes en este estudio coinciden con los registrados en otras comunidades tropicales con clima estacional en las que los himenópteros constituyen los polinizadores principales (Percival, 1974; Frankie et al., 1983; Ramírez, 1989; Ramírez et al., 1990; Ramírez y Brito, 1992; Oliveira y Gibbs, 2000; Machado y Lopes, 2004). Estos insectos, sobre todo las abejas de menor tamaño (≤1 cm) y las avispas, son visitantes asiduos de flores pequeñas (<1 cm), blancas, actinomorfas y de fácil acceso al néctar (formas de plato a tazón, cepillo, embudo y tubo corto), por lo que sus visitas generalmente se solapan con las de otros grupos de polinizadores poco especializados (p. ej., moscas) (Faegri y van der Pijl, 1979; Endress, 1994). Puede decirse entonces que este patrón floral, dado el carácter oportunista de las interacciones que promueve, representa más bien un síndrome dirigido a la polinización por insectos pequeños generalistas, tal como lo sugirieron Ramírez et al. (1990) y Ramírez y Brito (1992).
Las asociaciones observadas entre tipos de unidad de polinización (UP), forma, simetría y color de las flores fueron más o menos similares a lo que se ha señalado en la literatura. Por ejemplo, en un arbustal de la Guayana venezolana, Ramírez et al. (1990) encontraron que las flores organizadas individualmente están representadas en todos los colores excepto el verde; mientras que, en la organización colectiva, así como en formas intermedias, son más frecuentes los colores blanco-cremoso y verde. Por otra parte, estos autores refirieron que las flores radialmente simétricas son predominantemente blanco-cremosas y las flores sin plano simétrico son de color verde. En otro estudio, Ramírez (2003) encontró asociaciones entre el tipo actinomorfo (Leppik, 1969) y la forma de plato a tazón (Faegri y van der Pijl, 1979), así como entre el tipo zigomorfo (Leppik, 1969) y las formas de bandera y garganta (Faegri y van der Pijl, 1979), respectivamente. Estas asociaciones son congruentes, por un lado, con el síndrome alofílico (Faegri y van der Pijl, 1979) dominante en la comunidad (forma plato a tazón - simetría radial - color blanco) y por el otro, con dos síndromes que reflejan una mayor especialización frente al vector de polen. Estos son la combinación de UP colectiva, asimetría y color pardo-verde en especies predominantemente anemófilas, y la combinación de zigomorfía con las formas de bandera y garganta en especies estrictamente melitófilas (Faegri y van der Pijl, 1979).
La variabilidad de las dimensiones florales entre tipos de unidad de polinización (UP), forma, simetría y color de las flores también fue coincidente con los resultados de Ramírez et al. (1990). Estos autores refirieron que las flores organizadas en unidades de polinización colectiva e intermedia son más pequeñas que las flores solitarias, y que el largo de las flores, siguiendo el esquema de tipos florales de Leppik (1969), tiende a incrementarse desde formas haplomorfas-actinomorfas hasta las estereomorfas. En el sistema de Faegri y van der Pijl (1979), esto último equivale a un aumento del largo floral desde el tipo plato a tazón hasta los tipos embudo-tubo, respectivamente (Ramírez, 2003). En cuanto a la simetría y el color, Ramírez et al. (1990) señalaron que las flores asimétricas son más pequeñas que las radiales y las bilaterales. Asimismo, hicieron énfasis en el menor tamaño de las flores verdes con respecto a flores en otras categorías de colores. En el contexto de este trabajo, el menor tamaño de las flores organizadas en UP colectiva, con forma inconspicua, simetría irregular y/o color pardo-verde (si bien las diferencias en cuanto a color no fueron significativas), parece estar principalmente determinado por la ausencia de estructuras de atracción en las especies con síndrome anemófilo, y en el caso particular de Lemna aequinoctialis Welw. y Pistia stratiotes, por las características de su modo de reproducción. En este sentido, la baja relación polen/óvulo de L. aequinoctialis y P. stratiotes (Barrios y Ramírez, 2020) sugiere autogamia, sistema que también implica una reducción en el tamaño de las flores (Cruden, 1977).
La proporción de cada síndrome de polinización fue similar a la registrada por los modos de polinización correspondientes en otras comunidades tropicales con clima estacional. En este sentido, la proporción de especies melitófilas se asemejó a la de especies polinizadas por abejas en el Cerrado de Brasil (Silberbauer-Gottsberger y Gottsberger, 1988), en un morichal (pantano de palmeras) de los Llanos centrales (Ramírez y Brito, 1992) y en un arbustal de la Guayana (Ramírez, 1989) en Venezuela. La proporción de especies anemófilas y falenófilas, en cambio, fue alta en comparación con la compilación que hacen Machado y Lopes (2004) de la polinización por el viento y por esfíngidos en varias comunidades tropicales, e incluso mayor que los datos del morichal estudiado por Ramírez y Brito (1992). En el caso de la anemofilia, esta discrepancia puede deberse a la alta riqueza de ciperáceas en esta comunidad (Barrios et al., 2007), lo que eleva la incidencia del síndrome de polinización. Respecto a la cantaridofilia, su representación fue similar a la proporción de especies polinizadas por escarabajos en la Caatinga (Machado y Lopes, 2004) y el Cerrado (Silberbauer-Gottsberger y Gottsberger, 1988; Oliveira y Gibbs, 2000), así como en el morichal de los Llanos (Ramírez y Brito, 1992) y en el arbustal de la Guayana (Ramírez, 1989). Del mismo modo, la proporción de especies ornitófilas fue semejante a la de especies polinizadas por colibríes en el Cerrado (Silberbauer-Gottsberger y Gottsberger, 1988; Oliveira y Gibbs, 2000), mientras que la miofilia estuvo poco representada si se considera la importancia de las moscas como polinizadoras en el Cerrado (Silberbauer-Gottsberger y Gottsberger, 1988) y el arbustal guayanés (Ramírez, 1989).
Por otra parte, si se analizan directamente las clases de polinizadores observados en la comunidad, la polinización por abejas se mantiene por encima de la proporción que muestra este modo de polinización en el morichal estudiado por Ramírez y Brito (1992), al igual que en las diferentes comunidades revisadas por Machado y Lopes (2004). Además, al hacer este análisis resalta el papel de las avispas que pasa desapercibido si se intentan predecir los modos de polinización con base en los síndromes de polinización de las especies (p.ej. Guazuma ulmifolia Lam.). Del mismo modo, las moscas y los colibríes desempeñan un papel más importante como polinizadores que lo sugerido por las características de las flores. Esta discrepancia posiblemente esté determinada por el patrón de interacción de algunas especies melitófilas que, dada su facilidad de acceso a las recompensas florales (forma plato a tazón, cepillo, embudo, tubo corto, bandera) logran ser polinizadas, más allá de las abejas, por otras clases de agentes como avispas, moscas y colibríes. Este sistema polifílico (Faegri y van der Pijl, 1979) puede ser más ventajoso para las especies de plantas, ya que les permitiría mantener el servicio de polinización ante variaciones espacio-temporales en la disponibilidad de polinizadores e incluso frente a la pérdida definitiva de algunas interacciones específicas (Waser et al. 1996).
En cuanto a la fenología, cabe destacar que la distribución al azar de los períodos de floración de las especies que comparten vectores bióticos de polinización ha sido reportada con anterioridad, incluso más comúnmente que el esquema segregado (Stiles, 1978; Poole y Rathcke, 1979; Rabinowitz et al., 1981; Rathcke, 1988; Díaz et al., 1994; Brody, 1997; Boulter et al., 2006). Entre las causas de este patrón se han señalado la estocasticidad ambiental, las restricciones ecofisiológicas de cada una de las especies y las impuestas por efecto de la filogenia que impiden el desplazamiento de los eventos de floración (Kochmer y Handel, 1986). Las diferentes historias de vida de las especies también pueden incorporar restricciones determinantes (van Schaik et al., 1993). Por ejemplo, la evasión al parasitismo en la fase de yemas florales, el tiempo necesario para la maduración de los frutos, el escape a depredadores y/o patógenos de semillas, las condiciones para la germinación y supervivencia de las plántulas (Rathcke y Lacey, 1985; Brody, 1997; Elzinga et al., 2007). Lo anterior sugiere que las presiones de selección relacionadas con el uso compartido de los polinizadores deben causar más frecuentemente diferencias en caracteres como el color y/aroma de las flores (Johnson et al., 2005; Van der Niet et al., 2010; Peter y Johnson, 2014), la cantidad, calidad y/o forma de presentación de las recompensas ofertadas a los polinizadores (Bosh et al., 1997), el horario de antesis (Stone et al., 1998), entre otros. No obstante, algunos autores argumentan que la propia aleatoriedad de los períodos de floración es suficiente para minimizar la transferencia de polen entre especies (Boulter et al., 2006), lo cual puede ser apoyado por los altos niveles de especificidad de carga y transporte de polen que han sido registrados en los polinizadores de varias de las especies estudiadas (Barrios, 2017).
Los resultados de esta investigación permiten concluir que muchas de las especies presentan más bien un síndrome alofílico que puede ser resumido en las siguientes características de las flores: forma abierta o poco restrictiva, simetría radial y color blanco o blanco-amarillento. Es posible que este síndrome, lejos de ser perjudicial para la aptitud biológica de las especies, redunde en un beneficio colectivo de atracción de polinizadores oportunistas. Los efectos negativos derivados de la promiscuidad de las interacciones podrían estar minimizados, al menos en parte, por la distribución al azar de los períodos de floración de las especies que coinciden en sus características florales.

