











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Pseudognaphalium Kirp. es un género de la tribu Gnaphalieae, familia Asteraceae, propuesto por Kirpicznikov (1950). La especie tipo es P. oxyphyllum (DC.) Kirp., la cual fue segregada de Gnaphalium L. (Kirpicznikov y Kuprijanova, 1950). Años más tarde, Hilliard y Burtt (1981) y Anderberg (1991) transfirieron diez, y aproximadamente 70 especies más de Gnaphalium a Pseudognaphalium, respectivamente. Recientemente se han realizado más cambios en otros Pseudognaphalium (Kartesz, 1999; Hinojosa-Espinosa y Villaseñor, 2014; Dillon y Luebert, 2015), y también se describió una nueva especie (Pruski, 2012). Una comparación completa entre Pseudognaphalium y Gnaphalium, así como la relación con otros géneros, se presenta en la publicación de Hinojosa-Espinosa y Villaseñor (2014). En Gnaphalium las plantas son anuales, hasta 30 cm de alto con cabezuelas agrupadas en inflorescencias espiciformes o glomérulos, vilano dimórfico y estereoma dividido; mientras que en Pseudognaphalium son plantas de hasta 220 cm de alto con cabezuelas agrupadas en inflorescencias corimbiformes, vilano monomórfico y estereoma dividido o fenestrado.
En 1972, McVaugh describió Gnaphalium vulcanicum I.M. Johnst. var. monticola McVaugh (= Pseudognaphalium liebmannii (Sch. Bip. ex Klatt) Anderb. var. monticola (McVaugh) Hinojosa & Villaseñor) a la que diferenció de la variedad típica (= P. liebmannii var. liebmannii) por la altura de la planta (50-100 vs. <30 cm), longitud de las hojas (50-75 vs. <50 mm), pubescencia foliar (con glándulas vs. sin glándulas), forma del ápice (acuminado vs. agudo), longitud del involucro (4-7 vs. 6.5-9 mm de alto) y número de flores femeninas (60-80 vs. 125-200) y hermafroditas (10-20 vs. 5-7), por cabezuela.
Pseudognaphalium oxyphyllum es una especie con un intervalo de distribución geográfica y altitudinal amplio; por ello se han segregado tres taxones: P. oxyphyllum var. oxyphyllum, P. semilanatum (DC.) Anderb. y P. oxyphyllum var. nataliae (F.J. Espinosa) Hinojosa & Villaseñor. Este último se distingue de la variedad típica por ser plantas de mayor porte (≤2.2 m alto vs. <0.7 m), ápice de la hoja agudo a acuminado (vs. acuminado a apiculado), tallo alado (vs. sin alas o las alas no abarcan todo el entrenudo), pubescencia lanoso-araneosa (vs. araneosa densa o escasa), base de las hojas decurrente a lo largo del entrenudo (vs. no decurrente a lo largo del entrenudo) e involucro grisáceo (vs. blanco, pajizo a rojizo). Espinosa-García (1983) propuso que la variedad nataliae tiene afinidad con P. liebmannii var. monticola, por las características del involucro y el número de flores; también consideró la posibilidad de que sea resultado de hibridación entre P. oxyphyllum var. oxyphyllum y P. liebmannii var. monticola.
Una especie se conceptualiza de muchas formas; en este trabajo se utiliza el concepto biológico que la define como un conjunto de individuos con características morfológicas similares, con distribución geográfica definida y aislamiento reproductivo (Crisci, 1994). El concepto de variedad debe aplicarse cuando existen patrones de variación morfo-geográfica dentro de las poblaciones de una especie; es decir, que se presenten una o algunas diferencias morfológicas notorias y se tenga distribución alopátrica con superposición entre las poblaciones (Stuessy, 2009). Dada la complejidad natural de la variación infraespecífica, en algunos casos tanto el empleo de métodos fenéticos, como la aplicación de análisis estadísticos multivariados ayudan a resolver dudas sobre los niveles de jerarquización (Stuessy, 2009).
Aunque prácticamente todas las especies conocidas para México con las características de Pseudognaphalium han sido reubicadas en este género, aquí se analizan cuatro de sus variedades (P. liebmannii var. liebmannii, P. liebmannii var. monticola, P. oxyphyllum var. oxyphyllum y P. oxyphyllum var. nataliae) con el objetivo de evaluar si existe soporte para reconocer cada una como especie.
Materiales y Métodos
Con el propósito de comparar y analizar la variación morfológica de las cuatro variedades de Pseudognaphalium, se revisó el material disponible en los herbarios ENCB (Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional), MEXU (Instituto de Biología, Universidad Nacional Autónoma de México) y XAL (Instituto de Ecología, A.C.) (acrónimos según Thiers, 2019). Estas colecciones tienen una buena representación de las especies analizadas. Además, se examinaron las imágenes digitales de los ejemplares tipo (JSTOR, 2019), localizados en los herbarios de la Universidad de Copenhague (C, 2019), Conservatorio y Jardín Botánico de la Ciudad de Génova (G, 2019), Universidad de Michigan (MICH, 2019) y Universidad Nacional Autónoma de México (MEXU, 2019) (acrónimos según Thiers, 2019).
Del material examinado se seleccionó una serie de características morfológicas y ecológicas contrastantes para diferenciar las variedades elegidas (Cuadro 1) y se realizó un análisis de similitud para ver la relación entre las muestras estudiadas.
Cuadro 1: Caracteres usados en el Análisis de Correspondencia Corregido (DCA).
| 1. Longevidad: 0=hierbas anuales o bianuales, 1=hierbas perennes. |
| 2. Altura de la planta (cm). |
| 3. Tallo: 0=sin alas, 1=entrenudos parcialmente alados, 2=entrenudos totalmente alados. |
| 4. Pubescencia (tallo): 0=lanosa, 1=araneosa. |
| 5. Pubescencia glandular: 0=ausente, 1=presente. |
| 6. Longitud de las hojas en inflorescencia: 0=menor que las hojas del tallo 1=igual que las hojas del tallo. |
| 7. Forma de la hoja (relación largo/ancho): 0=oblongas (4-5), 1=lineares (7-15). |
| 8. Presencia de aurículas de las hojas: 0=ausentes, 1=presentes. |
| 9. Forma del ápice de la hoja: 0=agudo, 1=acuminado. |
| 10. Margen de la hoja: 0=plano, 1=revoluto. |
| 11. Coloración de la hoja: 0=concolora, 1=discolora. |
| 12. Pubescencia del envés de la hoja: 0=lanosa, 1=lanoso-araneosa, 2=araneosa. |
| 13. Longitud del pedúnculo en milímetros: 0=2-6, 1=6-12. |
| 14. Forma del involucro: 0=campanulado, 1=subgloboso. |
| 15. Longitud del involucro (mm). |
| 16. Color de las brácteas involucrales: 0=blancas a amarillentas. 1=pajizas, 2=rosadas. |
| 17. Número de flores femeninas por cabezuela. |
| 18. Número de flores hermafroditas por cabezuela |
| 19. Tipo de vegetación donde crece la planta: 0=bosque mesófilo, 1=bosque de pino, pino-encino, 2=vegetación subalpina. |
| 20. Altitud donde crece la planta (m): 0=1000-2000, 1=2000-3000, 2=3000-4600. |
Se evaluaron 77 ejemplares de herbario que representan la diversidad morfológica de las variedades a lo largo de su distribución: 20 de P. leibmannii var. liebmannii, 15 de P. liebmannii var. monticola, 15 de P. oxyphyllum var. nataliae y 27 de P. oxyphyllum var. oxyphyllum. Se analizaron 20 caracteres para cada espécimen (Cuadro 1) y se creó una matriz de datos (Apéndice), la cual se evaluó con un Análisis de Correspondencia Corregido (Detrended Canonical Analysis DCA, por sus siglas en inglés), que es una técnica estadística de análisis multivariado (Hill y Gauch, 1980), realizada con el programa CANOCO versión 4.5 (Braak y Šmilauer, 2002). Este método muestra la proximidad (similitud) entre las muestras utilizando una representación gráfica.
Resultados
La revisión de material de herbario y de campo muestra diferencias notorias entre Pseudognaphalium oxyphyllum var. nataliae y P. liebmanii var. monticola. Algunas de estas características contrastantes son la longevidad, altura de la planta, ápice de la hoja, presencia o ausencia de alas en el tallo y número de flores femeninas por cabezuela (Cuadro 2).
Cuadro 2: Comparación de caracteres en cuatro variedades de Pseudognaphalium Kirp. (Rzedowski et al., 2005; Pruski, 2018 y este trabajo).
| Carácter | Pseudognaphalium liebmannii (Sch. Bip. ex Klatt) Anderb. var. liebmannii | Pseudognaphalium liebmannii (Sch. Bip. ex Klatt) Anderb. var. monticola (McVaugh) Hinojosa & Villaseñor | Pseudognaphalium oxyphyllum (DC.) Kirp. var. nataliae (F.J. Espinosa) Hinojosa & Villaseñor | Pseudognaphalium oxyphyllum (DC.) Kirp var. oxyphyllum |
| Longevidad | perenne | anual o bianual | perenne | anual |
| Altura (cm) | 10-30(-40) | 40-140 | 40-220 | 20-70(-120) |
| Tallo | sin alas | sin alas | alado | sin alas o las alas no abarcan todo el entrenudo |
| Pubescencia (tallo) | lanosa-aracnosa | lanosa-araneosa | lanosa-araneosa | araneosa densa o escasa |
| Tricomas glandulares | ausentes o poco evidentes | presentes | presentes | presentes |
| Hojas en inflorescencia | no reducidas | no reducidas | menor tamaño | menor tamaño |
| Forma de hoja | linear a lanceolada | linear a lanceolada | oblanceolada | oblanceolada |
| Aurículas en las hojas | ausentes | ausentes/presentes | presentes | presentes |
| Ápice de la hoja | agudo o obtuso | acuminado | agudo a acuminado | acuminado a apiculado |
| Margen de la hoja | revoluto | plano/revoluto | plano | plano |
| Coloración de la hoja | concoloras | discoloras | discoloras | concoloras |
| Pubescencia envés | lanosa | lanosa | lanosa-araneosa | araneosa densa a escasa |
| Tamaño de pedúnculo (mm) | 2-5 | 2-10 | 2-5 | 2-8 |
| Forma de involucro | subgloboso | subgloboso | campanulado | campanulado |
| Tamaño de involucro (mm) | 5-9 | 4-7 | (3-)4-5 | 4-5 |
| Color de brácteas involucrales | pardo-amarillentas | amarillo-parduzcas | blanco-amarillentas, verdosas | blancas, amarillentas a rojizas |
| Flores femeninas | 90-100 | 50-70 | 20-50 | 30-50 |
| Flores hermafroditas | 5-10 | 12-24 | 2-9 | 3-10 |
| Altitud (m) | 2400-4300 | 1800-3800 | 2500-3800 | 1000-3600 |
| Distribución | Centro y Sur de México y Guatemala | Centro y Sur de México | Centro de México | México y Guatemala |
En la Figura 1, se representa gráficamente el eje I (porcentaje acumulado de varianza de 81.4%) y el eje II (porcentaje acumulado de varianza de 88.6%). El diagrama de ordenación muestra que la mayoría de los especímenes están positivamente correlacionados con el eje I. La misma figura presenta todos los elementos formando cuatro agrupaciones (L, M, N, O). Los ejemplares de P. liebmannii var. liebmannii correspondientes al grupo L 1-20, se localizan separados de los otros grupos. Las muestras de P. liebmannii var. monticola, que forman el conjunto M 21-35, son los más cercanos a los de P. liebmannii var. liebmannii (grupo L), pero con un poco de separación. A su vez es el más próximo a P. oxyphyllum var. nataliae, cuyos ejemplares forman la agrupación N 36-50, que están cercanos a P. liebmannii var. monticola y muy próximos a P. oxyphyllum var. oxyphyllum, cuyos representantes constituyen el conglomerado O 51-77. Dado que los grupos representados en la gráfica no muestran continuidad entre ellos, se propone el cambio de las variedades monticola y nataliae a la categoría de especie. A continuación, se presenta una clave de identificación para los taxones estudiados.
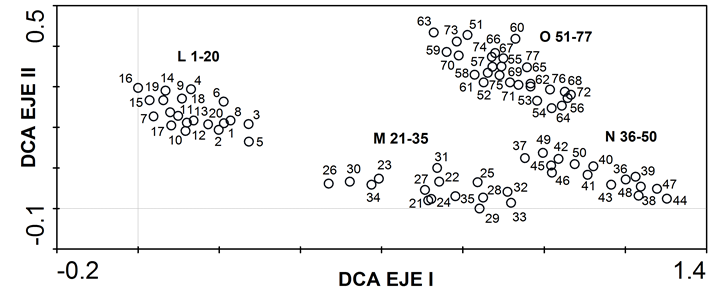
Figura 1: Análisis de Correspondencia Corregido (DCA) donde se muestra los resultados de agrupamiento de acuerdo al análisis de similitud. Pseudognaphalium liebmanii (Sch. Bip. ex Klatt) Anderb. var. liebmannii (L 1-20); P. liebmannii (Sch. Bip. ex Klatt) Anderb. var. monticola (McVaugh) Hinojosa & Villaseñor (M 21-35); P. oxyphyllum (DC.) Kirp. var. nataliae (F.J. Espinosa) Hinojosa & Villaseñor (N 36-50); P. oxyphyllum (DC.) Kirp. var. oxyphyllum (O 51-77).
Clave de identificación para los taxones estudiados
1a. Hojas oblanceoladas, auriculadas; involucros campanulados, blancos a amarillentos, verdosos o rojizos; hojas cercanas a la inflorescencia, de menor longitud que las del tallo …………….….……………………………………………………………………. 2
1b. Hojas lineares o lanceoladas, usualmente sin aurículas; involucros subglobosos, pardo-amarillentos a parduzcos; hojas cercanas a la inflorescencia de igual longitud que las del tallo ……………………………………………………………………………………. 3
2a. Plantas perennes; tallos alados; hojas discoloras ….………………………………. P. nataliae (F.J. Espinosa) Villarreal, A.E. Estrada & Encina
2b. Plantas anuales; tallos parcialmente alados; hojas concoloras …………………………………………………………...……. P. oxyphyllum (DC.) Kirp.
3a. Plantas 10-30(-40) cm de alto; hojas concoloras; flores periféricas 90-100 ..………………………………………………..... P. liebmannii (Sch. Bip. ex Klatt) Anderb.
3b. Plantas 40-140 cm de alto; hojas discoloras; flores periféricas 50-70 ..………………………….……. P. monticola (McVaugh) Villarreal, A.E. Estrada & Encina
Pseudognaphalium monticola (McVaugh) Villarreal, A.E. Estrada & Encina, stat. nov.
≡ Gnaphalium vulcanicum I.M. Johnst. var. monticolaMcVaugh, Contr. Univ. Michigan Herb. 9: 466. 1972. TIPO: MÉXICO. Jalisco. Autlán de Navarro, 25-30 km SE of Autlán, between El Chante and Cuzalapa, R. McVaugh 23126 (holotipo: MICH 1107419!, isotipos: CAS 2859!, ENCB 3706!, NY 169500!).
≡ G. liebmannii Sch. Bip. ex Klatt var. monticola (McVaugh) D.L. Nash, Fieldiana Bot. 36: 74. 1974.
≡ P. liebmannii (Sch. Bip. ex Klatt) Anderb. var. monticola (McVaugh) Hinojosa & Villaseñor, Botanical Sciences 92(4): 490. 2014.
Esta especie ha sido tratada como una variedad de P. liebmannii; en el Cuadro 2 se resumen las características contrastantes encontradas en este trabajo y en la revisión de la literatura (Espinosa-García, 1983; McVaugh, 1984; Pruski, 2018).
Ejemplares examinados de P. liebmannii var monticola: MÉXICO. Ciudad de México, alcaldía Cuajimalpa, Desierto de Los Leones, 19°18'30''N, 99°18'30''O, 9.IV.1967, O. Aguirre 624 (ENCB); SW de la Ciudad de México, 19°13'51''N, 99°13'43.9''O, prado, 2960 m, 9.VIII.2014, M. Galbany 2422 (MEXU). Estado de México, municipio Amecameca, 2 km al SE de San Pedro Nexapa, 19°04'06''N, 98°43'07''O, 2.VIII.1964, M. Villegas 804 (ENCB). Municipio Jilotzingo, 3 km al NO de San Luís Ayucan, 19°30'46''N, 99°22'54''O, bosque de Abies, Quercus y Pinus, 29.X.1978, M. Gómez 228 (ENCB). Hidalgo, municipio Tepeapulco, parte alta del cerro Xihuingo, 19°49'N, 98°35'O, 7.X.1973, J. Rzedowski 12047 (ENCB). Jalisco, municipio Autlán, Sierra de Manantlán, along lumber-roads east of the road crossing La Cumbre, between El Chante and Cuzalapa, 19.583°N, 104.192ºO, pine forest, 2750 m, 20.III.1965, R. McVaugh 23126 (MICH). Municipio Magdalena, La Joya, cima del volcán Nevado de Colima, bosque de Pinus hartwegii y Cupressus, 3280 m, 30.IX.1987, A. García 3517 (MEXU). Municipio Tequila, volcán de Tequila, 20°47'12''N, 103°50'54''O, 25.I.1985, L. M. Villarreal 1025 (XAL). Municipio Zapotitlán El Grande, faldas del Nevado de Colima, 19°33'N, 103°36'O, 12.IX.1986, R. Ornelas 433 (XAL). Michoacán, municipio Morelia, lado E del cerro El Águila, subiendo por el poblado de San Nicolás Obispo, 19.62472°N, 101.34583°O, 2385 m, 19.I.2008, E. Sánchez 531 (MEXU). Municipio Paracho, parte alta del cerro El Águila, 19°35'22''N, 102°02'17''O, 24.VIII.1990, E. García 743 (XAL). Morelos, municipio Tepoztlán, San Juan Tlacotenco, 7 km al N, bosque de pino-encino, 2850 m, 1.VIII.1987, M. L. Espín 114 (MEXU). Veracruz, municipio Las Vigas, rancho Llano Grande, 19°40'19''N, 97°03'33''O, F. Ventura 666 (ENCB). Municipio Perote, Cofre de Perote, falda O, 19°29'48''N, 97°09'48''O, 4000 m, 25.XI.1975, J. Dorantes 5305 (XAL). Municipio Xico, 1 km al SE de Tembladeras, 7.V.1986, R. Arriaga 288 (XAL).
Pseudognaphalium nataliae (F.J. Espinosa) Villarreal, A.E. Estrada & Encina, stat. nov.
≡ Gnaphalium oxyphyllum DC. var. nataliae F.J. Espinosa, Bol. Soc. Bot. México 45: 17. 1983. TIPO: MÉXICO. Estado de México, Amecameca, alrededores de San Pedro Nexcapa, F. J. Espinosa 766 (holotipo: MEXU 343482!, isotipo: ENCB 3705!).
≡ P. oxyphyllum (DC.) Kirp. var. nataliae (F.J. Espinosa) Hinojosa & Villaseñor, Botanical Sciences 92(4): 491. 2014.
Originalmente esta especie fue propuesta como una variedad de G. oxyphyllum (Espinosa-García, 1983), con la cual se contrastan las características en el Cuadro 2.
Ejemplares examinados de P. oxyphyllum var. nataliae: MÉXICO. Ciudad de México, alcaldía Magdalena Contreras, dínamo Contreras, Rastrojera, 2600 m, X.1929, E. Matuda 18704 (MEXU). Alcaldía Milpa Alta, faldas del volcán Acopiaxco, 3200 m, 15.XI.1992, R. Giles 18 (MEXU). Estado de México, municipio Amecameca, alrededores de San Pedro Nexcapa, bosque de pino-oyamel, 3000 m, 21.IX.1979, F. J. Espinosa 766 (MEXU). Municipio Iturbide, alrededores de la presa Iturbide, bosque de Abies, 3250 m, 21.XI.1982, J. Rzedowski 38024 (MEXU). Municipio México, sierra Las Cruces, 21.X.1892, C. G. Pringle 4312 (MEXU). Municipio San José del Rincón, 19.59431°N, 100.251°O, 3620 m, 24.XI.2014, D. Álvarez 14208 (ENCB). Municipio Zoquiapan, Ixtapaluca, bosque de Pinus, Alnus, Ulmus, 3280 m, 1.XI.1978, L. Rico 23 (MEXU). Michoacán, municipio Angangueo, El Rosario, 7 km SE de Angangueo, 3050 m, 1.XII.1984, J. C. Soto 6956 (MEXU). Municipio Morelia, lado O del cerro El Águila, aproximadamente 4 km SE de Iratzio, 19.62944°N, 101.375°O, 2800 m, 15.XI.2010, M. Flores 147 (MEXU). Morelos, municipio Huitzilac, Parque Nacional Lagunas de Zempoala, 8.XII.1983, V. Peñaloza 188 (MEXU). Puebla, municipio Santa Cruz Texmalaquilla, Pico de Orizaba, 27.IX.1973, J. Rodríguez s.n. (MEXU). Veracruz, municipio Calcahualco, 9.5 km by road W of Escola along Coscomatepec - Escola-Jacal road, 15.XI.1981, M. Nee 23167 (XAL). Municipio Las Vigas, Las Lachas, carretera Perote - Jalapa, 19.IX.1972, L. W. Boege 2477 (MEXU). Municipio Mariano Escobedo, Agua de San Pablo, 4.5 km de Pilancón, 11.VIII.1988, J. L. Ramírez 103 (XAL). Municipio Rafael Ramírez, rancho San Isidro El Chico, a 2 km de Los Pescados, 17.X.1984, L. Tapia 485 (XAL).
Ejemplares examinados de las variedades que conservan su jerarquía taxonómica.
Ejemplares examinados de P. liebmannii var. liebmannii: MÉXICO. Chiapas, municipio San Cristóbal de las Casas, San Juan Chamula, 10.XI.1972, D. E. Breedlove 29338 (MEXU). Ciudad de México, alcaldía Magdalena Contreras, Las Palmas, cañada de Contreras, IX.1932, E. Lyonnet 789 (MEXU). Alcaldía Tlalpan, El Ajusco, 19°30'10''N, 99°12'08''O, 14.VII.1958, J. H. Beaman 2254 (ENCB). Colima, municipio Comala, rancho El Jabalí, 22 km NNW of city of Colima at Jalisco state line, near hacienda San Antonio, 19°27'N, 103°42'O, 23.VIII.1988, A. C. Sanders 448 (XAL). Estado de México, municipio Amecameca, cañada del cerro Venacho, 6 km al E de Amecameca, 19°07'02''N, 98°42'17''O, 3.II.1978, J. García 1748 (ENCB); Joya de Alcalican, extremo S del Iztaccíhuatl, 19°08'33''N, 98°38'55''O, límite de vegetación arbórea y zacatonal, 14.XI.1976, P.M. Rubio 1021 (XAL). Municipio Tlamacas, Atlautla, 19°03'30''N, 98°38'18''O, 24.X.1982, I. Díaz 321 (XAL). Jalisco, municipio Tuxpan, Nevado de Colima, 3850 m, 05.VII.1975, C. L. Díaz 6026 (MEXU). Municipio Zapotitlán de Vadillo, upper slopes of the peak of the Nevado de Colima, near timberline, 3300-4340 m, 13.IX.1952, R. McVaugh 12905 (MEXU). Michoacán, municipio Angangueo, Santuario Sierra de Chincua, 3250 m, 12.XI.2005, M. A. Salinas 784 (MEXU). Municipio Contepec, 3 km del camino de Santa María de los Ángeles a Solís, 19°58'26''N, 100°07'41.02''O, 25.IX.2004, J. Martínez 1067 (MEXU); Cerro Altamirano, Reserva de la Biosfera de la Mariposa Monarca, 19°58'36.01''N, 100°08'15''O, 3130 m, 29.X.2005, M. A. Salinas 541 (MEXU). Morelos, municipio Tepoztlán, 5 km al W de San Juan Tlacotenco, 2775 m, 16.VIII.1987, M. L. Espín 132 (MEXU). Puebla, municipio Santa Cruz Texmalaquilla, Pico de Orizaba, cerca de Cueva del Muerto, 2800 m, 30.III.1940, F. Miranda 370 (MEXU); vertiente del volcán Pico de Orizaba, en los límites de Puebla - Veracruz, 3400 m, 27.VII.1971, L. J. Nevling 2049 (MEXU); Pico de Orizaba, vereda que lleva al pastizal alpino, partiendo de Texmalaquilla, 3600-4000 m, 16.X.1985, J. L. Villaseñor 850bis (MEXU). San Luís Potosí, municipio Guadalcázar, San José de las Flores, La Borrega hacia Pozas, 22°48'N, 100°23'O, 14.X.2000, R. Torres 15954 (MEXU). Veracruz, municipio Calcahualco, Pico de Orizaba, border of Puebla and Veracruz, F. M. Liebmann 310 (C, fototipo MEXU); 5 km NW of Jacal, NE slopes of Pico de Orizaba, 6.VII.1982, M. Nee 24820 (XAL). Municipio Perote, por la vereda a Los Altos de la Laguna Tilapa, parque nacional Cofre de Perote, 6.VII.1983, H. Narave 783 (ENCB).
Ejemplares examinados de P. oxyphyllum var oxyphyllum: MÉXICO. Chiapas, municipio Unión Juárez, volcán Tacaná, side of the summit, 15°07'50''N, 92°06'37''O, 10.XII.1972, D. E. Breedlove 44128 (TEX). Ciudad de México, alcaldía Coyoacán, Ciudad Universitaria, Centro Cultural Universitario, terreno baldío, 19°18'51''N, 99°11'11''O, vegetación ruderal, 2326 m, 10.XI.2008, O. Hinojosa 437 (MEXU). Alcaldía Cuajimalpa, Desierto de Los Leones, 19°18'30''N, 99°18'30''O, 00.XI.1953, L. Paray s.n. (ENCB). Alcaldía Tlalpan, km 10 de la carretera Picaho - Ajusco, cerca del parque El Tepozán, 19°15'36''N, 99°14'18''O, bosque de Quercus, 2859 m, 9.XI.2008, O. Hinojosa 462 (MEXU). Coahuila, municipio Arteaga, Los Lirios, La Jacinta, pine wood, 2390 m, 15.IX.1995, G. Hinton 25551 (TEX); La Jacinta a El Tunal, mixed forest, 2470 m, 15.IX.1995, G. Hinton 25596 (TEX). Estado de México, municipio México, sierra Las Cruces, 21.X.1892, C. G. Pringle 4312 (COLO). Municipio San Martín de las Pirámides, ladera SE del Cerro Gordo, 19°44'37''N, 98°50'00''O, 29.VII.1981. M. E. Castilla 547 (ENCB). Guanajuato, municipio Guanajuato, Villalpando au sud est de Guanajuato, s.f., J. Méndez s.n. (G). Hidalgo, municipio Mineral del Monte, barranca below La Trinidad iron works, 1676 m, 22.VII.1902, C. G. Pringle 8668 (MEXU). Municipio Pachuca de Soto, Cerro Gordo, 5 km al E de Pachuca, 20°07'N, 98°47'O, 14.IX.1975, J. Rzedowski 36578 (XAL). Municipio Tepeapulco, cerro Santa Ana, 19°45'45''N, 98°31'39''O, 8.IX.1976, A. Ventura 182 (ENCB). Jalisco, municipio Magdalena, N slope of Nevado de Colima, road to Zapotlán, 16.X.1952, R. McVaugh 13552 (MEXU). Michoacán, municipio Salvador Escalante, Cerro Burro, 19°25'48''N, 101°30'48''O, 26.XI.1990, E. Pérez 339 (XAL). Municipio Zitácuaro, San Miguel Chichimequillas, volcán El Molcajete, 19°23'54''N, 100°22'05''O, vegetación secundaria, 2230 m, 13.VII.2007, C. Ledesma 355 (MEXU). Morelos, municipio Huitzilac, volcán Las Palomas, bosque de pino, 2500-3000 m, 22.XI.2007, T. Amezcua 43 (MEXU). Municipio Tepoztlán, Parque Nacional El Tepozteco, 19°01'45.15''N, 99°05'2.53''O, encinar, 2650 m, 22.XI.2010, B. Munguía 541 (MEXU). Nuevo León, municipio Galeana, Las Placetas, 24°55'10''N, 100°13'01''O, 30.IX.1989, G. Hinton 19828 (TEX). Oaxaca, municipio Miahuatlán, Neverías a San Miguel Amazotepec, pine forest, 2225 m, 5.VIII.1996, G. Hinton 26740 (TEX). Municipio San Pedro y San Pablo Ayutla, 3 km by road to center Ayutla, 17°01'N, 96°05'O, 16.XII.1985, M. Nee 24354 (XAL). Puebla, municipio Ahuazotepec, Beristain, 3.VII.2007, O. del Rosario 122 (MEXU). Sonora, municipio Álamos, Sierra de Álamos, ca. 5.5 km SW of Álamos, along the trail La Huerta to Pico de Águila, 26°58'45''N, 108°59'O, pine-oak woodland, 1600 m, 24.XI.1997, V. W. Steinmann 1330 (MEXU). Veracruz, municipio Altotonga, Fesahuapa, cerca de Ermita, 24.XII.1972, F. Ventura 18 (ENCB). Municipio Perote, 3 km de Tenextepec, camino al Escobillo, 19°29'N, 97°15'O, 27.X.1997, J. Becerra 249 (XAL); Caja de Agua, F. Ventura 7467 (ENCB). Municipio Xalapa, 1 mi N of La Joya, pine-oak forest, 2040 m, 7.XII.1974, T. F. Stuessy 3664 (MEXU). Zacatecas, municipio Jalpa, 38 km al W por la carretera Tlaltenango de Sánchez Román, 30 km del entronque de la carretera Jalpa - Juchipila, 23.X.1973, J. Rzedowski 955 (TEX).
Discusión
En la Figura 1, el grupo de P. liebmannii var. liebmannii se presenta aislado de los otros; de acuerdo con esto, es la variedad que mejor se caracteriza morfológicamente y se corresponde con las plantas perennes de menor tamaño, con tricomas glandulares escasos o poco visibles en el indumento, cabezuelas con 90-100 flores; su distribución es frecuente en las partes más altas de las montañas (Cuadro 2). Pseudognaphalium liebmannii var. monticola se muestra como un grupo un poco extendido horizontalmente con algunos elementos ligeramente separados (Fig. 1), lo que indica variación morfológica, sobre todo en el tamaño de las plantas, forma de las hojas, presencia de aurículas en las hojas y número de flores por cabezuela (50-70). Presenta mayor afinidad morfológica con P. oxyphyllum var. nataliae, por ello está más cerca de él que con P. liebmannii var. liebmannii.
Pseudognaphalium oxyphyllum var. nataliae muestra extensión corta horizontal que indica variación morfológica como es el caso del tamaño de la planta, ápice de la hoja, tamaño del involucro de 3-5 mm de alto, y número de flores por cabezuela (20-50). Se localiza en una posición intermedia entre los grupos de P. liebmannii var. monticola y P. oxyphyllum var. oxyphyllum, mostrando mayor afinidad con ambos. En la Figura 1 no se muestran ejemplares mezclados de P. liebmannii var. monticola y P. oxyphyllum var. oxyphyllum, como para validar la hipótesis de contacto genético entre ellos propuesta por Espinosa-García (1983), quien también postuló que P. oxyphyllum var. nataliae es resultado de la hibridación entre P. liebmanni var. monticola y P. oxyphyllum var. oxyphyllum, lo cual puede quedar evidenciado por su posición intermedia, pero tendría que ser validado genéticamente.
El grupo de P. oxyphyllum var. oxyphyllum es bastante variable, posiblemente por tener un rango de distribución amplio; es cercano a P. oxyphyllum var. nataliae existiendo proximidad entre ellos y, por lo tanto, mayor similitud; aunque hay una separación sin continuidad o mezcla de sus integrantes que revela independencia entre las dos agrupaciones.
Pseudognaphalium liebmannii var. monticola también presenta cercanía con P. oxyphyllum var. nataliae, mostrando mayor afinidad morfológica. Pruski (2018) considera que podría ser un taxón intermedio entre P. liebmannii y P. oxyphyllum por tener una mezcla de características de ambos. Debido a que los elementos de los grupos no se encuentran mezclados ni hay continuidad entre ellos, se descarta la posibilidad de que uno represente una variedad de otro; por lo tanto, cada agrupamiento apoya la propuesta de que sus elementos corresponden a especies diferentes. En los ejemplares examinados tampoco se observaron individuos con características que permitan suponer que sean representantes híbridos.
Con base en el análisis anterior, se propone que Pseudognaphalium liebmannii var. monticola y P. oxyphyllum var. nataliae deben ser tratadas como especies. Pruski (2018) menciona que es posible que P. liebmannii var. monticola merezca reconocimiento a nivel específico. Asimismo, algunos de los ejemplares de P. oxyphyllum var. nataliae revisados en MEXU tienen anotaciones de Guy L. Nesom con el nombre P. nataliae; sin embargo, en ninguno de los casos, hasta ahora, se formalizaron las jerarquías taxonómicas.
Conclusión
Mediante el uso de análisis multivariados se establece la formación de agrupaciones definidas y aisladas que permiten considerar Pseudognaphalium monticola y P. nataliae en nuevas jerarquías taxonómicas. Ambas especies se pueden reconocer por características de las hojas, presencia o ausencia de aurículas y alas en el tallo, longevidad, forma y color del involucro, así como diferencias en el número de flores por cabezuela.

