











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introduction
Gymnopodium Rolfe is one of the smallest genera of Polygonaceae, including only two species: G. floribundum Rolfe and G. toledense Ancona & Ortiz-Díaz (Ortiz-Díaz, 1994; Ancona et al., 2018). Its distribution occurs with disjunct populations in the Yucatán Peninsula, Oaxaca, Chiapas, Belize and Guatemala (Ortíz-Díaz, 1994; Burke and Sanchez, 2011), all localities in the biogeographic Mesoamerican Domain (Morrone, 2014). It is an important arboreal component of the dry forests of Mexico and the savannas of northern Belize, forming pure populations from sea level up to 1350 m elevation (Miranda, 1952; Flores and Espejel, 1994; Goodwin et al., 2013).
The species Gymnopodium floribundum, locally known in the Yucatán Peninsula as “tsi tsil che”, is of great economic importance for beekeeping of the Yucatán Peninsula, and it is highly valued on the international honey market (Alfaro-Bates et al., 2010). Moreover, trees of G. floribundum in Chiapas are associated with the production of the edible macromycet Tremelloscypha gelatinosa (Murrill) Oberw. & K. Wells (Bandala et al., 2014; Bandala and Montoya, 2015). The shrubs or small trees of G. floribundum are characterized by fissured bark, alternate or fasciculate leaves, short petiole, originating in deciduous ochreas when mature; its inflorescences are terminal racemes, whereas the flowers are hermaphrodite with six perianth segments, the three outer ones larger than the inner ones; the fruit is an achene covered by all the accrescent perianth segments (Ortiz-Díaz, 1994).
Phylogenetic studies in the subfamily Eriogonoideae confirm the monophyly of the genus Gymnopodium and place it as a sister group of the Eriogoneae tribe (Burke et al., 2010; Burke and Sanchez, 2011). In previous taxonomic studies of this genus, up to three species and two varieties have been recognized based on the presence or absence of indument, the shape of the leaf blades, and the perianth shape: G. antigonoides (B.L. Rob. ex Millsp. & Loes.) S.F. Blake, G. floribundum and G. ovatifolium (B.L. Rob. ex Millsp. & Loes) S.F. Blake, G. floribundum var. floribundum, and G. floribundum var. antigonoides (B.L. Rob. ex Millsp. & Loes) Standl. & Steyerm. However, in the latest taxonomic treatment of the genus, Ortiz-Díaz (1994) recognized G. floribundum as the only highly polymorphic species.
The taxonomic treatments of G. floribundum have been carried out without analyses of morphological variation and without covering the entire geographical and ecological distribution. Therefore, the goal of this study was to quantitatively evaluate the morphological variation with multivariate statistical and geometric morphometric tech niques to propose an infraspecific taxonomy of G. floribundum.
Materials and Methods
Selection of representative populations
The preliminary review of the morphological variation and geographical distribution assessed from herbarium specimens (BM, CICY, MEXU, MO and UADY) allowed us to organize nine representative populations: five from the Yucatán Peninsula, three from the Pacific Coastal Plain, and one from Belize (Table 1). Additionaly, we carried out field work in Mexico to gather representative samples of the eight populations. These specimens were deposited in the herbarium UADY. Specimens collected in Belize were obtained from the herbaria BM and MO.
Table 1: Geographic region, coordinates, elevation and number of specimens of the nine populations of Gymnopodium floribundum Rolfe analyzed. n=number of analyzed specimens. In bold face specimen nomenclature used for the dendrogram.
| Region | Locality | Geographic location | Elevation (m) | n |
| Yucatán Peninsula | 1.- Carretera Dzemul-Zona arqueológica de Xcambo, municipio Dzemul, Yucatán, Mexico. | 21°17'53.9''N 89°19'25.5''W | 8 | 30 |
| 2.- Carretera Huhi-Tixcacal, municipio Huhi, Yucatán, Mexico. | 20°42'11.8''N 89°08'54.8''W | 12 | 30 | |
| 3.- Carretera Vigia Chico-Punta Alem, municipio Felipe Carrilo Puerto, Quintana Roo, Mexico. | 19°55'54.89''N 87°47'3.60''W | 20 | 15 | |
| 4.- Camino hacia Dos Lagunas, municipio Calakmul, Campeche, Mexico. | 18°52'31.59''N 89°23'42.61''W | 216 | 18 | |
| 5.- Sitio arqueológico Santa Rosa Xtampak, municipio Hopelchén, Campeche, Mexico. | 19°47'26.22''N 89°35'9.16''W | 120 | 30 | |
| Pacific Coastal Plain | 6.- El Chorreadero, municipio Tuxtla Gutiérrez, Chiapas, Mexico. | 16°45'4.57''N 92°58'12.62''W | 650 | 30 |
| 7.- Entronque aeropuerto - Ocozocuautla carretera 190, municipio Ocozocuautla, Chiapas, Mexico. | 16°44'33.9''N 93°20'49.3''W | 1010 | 30 | |
| 8.- Las Anonas, municipio Juchitán de Zaragoza, Oaxaca, Mexico. | 16°39'50.08''N 94°47'27.47''W | 60 | 30 | |
| Belize | 9.- Autopista oeste Belmopan - Cd. Belice, Distrito El Cayo, Belize. | 17°16'35.34''N 88°37'20.87''W | 12 | 19 |
Grouping of specimens
The data matrix consisted of 224 specimens and 32 characters, of which 21 were quantitative and 11 qualitative (Appendix). Due to the mixed type of morphological descriptors, the Gower’s coefficient was elected as a distance measure and unweighted pair-group method (UPGMA) as linkage measure. Calculations were made with PAST3 software (Hammer et al., 2001) and the results were summarized in a dendrogram. The Gower is similar to the Manhattan distance but with the normalization of the range. It allows simultaneous manipulation of quantitative and qualitative variables in a database; through the application of this coefficient it is possible to find the similarity between individuals (Gower, 1971).
Distance data
Four vegetative measurements of mature leaves and brachyblasts were taken from the outermost whorl of leaves: length of the brachyblast, which was measured when it was lignified; length of the petiole; length and width of the leaf blade. Moreover, the reproductive characters were taken from inflorescences, flowers and fruits. We defined mature flowers when the perianth segments were pale brown in color and this indicated a well-developed achene. The seventeen measurements that we registered were the following: length of the peduncle and inflorescence; length from the pedicel base to the articulation and length from the pedicel articulation to the base of the perianth; distances between first and second floral fascicle, distances between second and third floral fascicle, and distances between third and fourth floral fascicle; length and width of the ovary, outer perianth segments, inner perianth segments and achenes; style length in ovary and the fruit (Fig. 1). For each specimen, a single measurement was taken of each character state. A list of definitions of the 21 distances is presented in the Appendix.

Figure 1: Characters of Gymnopodium floribundum Rolfe analyzed, abbreviations according to what is described in Table 2.
Shape data
For the geometric morphometric study we carried out separate analyses of the shape of the leaf blades and the perianth segments. The templates to register the outlines were built using the MakeFan8 program of the Integrated Morphometrics Package (IMP) series (Sheets, 2014a). MakeFan provides several options for drawing rays at equal angular intervals or combs perpendicular from a reference line that can be used as guides for digitizing semi-landmarks (Zelditch et al., 2004). For the leaves, a 15-ray comb was used, placing as reference points the insertion of the petiole with the blade and the apex. For the leaf outline, a configuration of 32 points was digitized: two landmarks and 30 semi-landmarks. For the outer segments of the perianth, the template consisted of a circle with 30 rays, taking as reference points the apex and the point of union of the main nerve with the primary veins, having a total of 31 points: two landmarks and 29 semi-landmarks. Landmarks are discrete anatomical loci that can be recognized as the same loci in all specimens in the study. The collection of points that describe an outline are called semi-landmarks, they are points located at concave or convex ends of processes and curvatures.
The Cartesian coordinates of each point were recorded in 224 specimens using the program tpsDig2 version 2.18 (Rohlf, 2015). In the Cartesian coordinates of marks an object is a collection of vectors (a tensor) and the collection of objects configures a cloud of points (tensors) in a Riemannian space. The objects or samples are points positioned in a multidimensional space defined by the basis vectors. The difference between objects or the sample is measured with Procrustes distances. The calculation of the distances between points is based on the Pythagorean theorem for the estimation of the hypotenuse of some implicit right triangle. For this, orthogonal axes that work as legs in that triangle are required. Landmark configurations for each shape separately were superimposed with the Procrustes model in the CoordGen8 software and the semilandmarks were aligned by using the Semiland8 tool (Sheets, 2014b). Superimposition methods eliminate non-shape variation in configurations of landmarks by overlaying them according to some optimization criterion. The Procrustes Analyses superimposes landmark configurations using least-squares estimates for translation and rotation parameters (Adams et al., 2004).
Distance morphometric analyses
The 21 quantitative characters were subjected to multivariate exploratory analyses (n=224), through the principal component analyses technique (PCA). The components were extracted from the correlation matrix, and the significant ones were retained by comparing observed eigenvalues with the broken stick distribution. The calculations were made with the FactoMineR program (Husson et al., 2007), implemented in the R programming environment (R Core Team, 2015), through the R-Commander program (Fox, 2005). With the three main groups identified in the previous step, we conducted non-parametric multivariate analyses of variance with permutations (PERMANOVA) to test the null hypothesis of equality in terms of the position of the multivariate centroids, using the Bray-Curtis distance as a metric of differences. Moreover, the null hypothesis of equality in multivariate dispersions was also tested for the three groups. Calculations were made with the “adonis” and “betadisper” routines, both contained in the “vegan” program (Oksanen et al., 2014), and implemented in R (R Core Team, 2015).
Geometric morphometric analyses
The superimposed and aligned Procrustes coordinates were used to explore patterns of shape variance among the three groups detected in the UPGMA analyses, as explained above. Multivariate analyses of variance of leaf shape separately from perianth segment shape were executed with all specimens in three “a priori” groups with a Canonical Variate Analyses in CVAGen8 (Sheets, 2014c). Partial shape deformations explained by each significant canonical axes were visualized using the interpolation technique Thin Plate Spline (TPS) in CVAGen8. The CVAGen8 software is used to calculate partial deformation scores and uniform components, extracting the canonical variations of such scores to generate a plot of distribution of points. It also uses a discriminant function to classify specimens into morphologically similar groups through an assignment test which determines the probability that specimens from a sample are closer to the group average to which it was assigned a priori than that from another group (Zelditch et al., 2004).
Distribution map
The distribution map of species and subspecies of Gymnopodium was made using the SimpleMappr program (Shorthouse, 2010) and geographic coordinates from the electronic database of the Missouri Botanical Garden (TROPICOS, 2019) and Open Data Portal of the UNAM (UNAM, 2019).
Results
The dendogram based on the qualitative and quantitative characters employed showed three main clusters: the first represented by the Belize specimens (black cluster), the second those of the Yucatán Peninsula (red cluster), and the third cluster (green) the specimens of Chiapas and Oaxaca (Pacific Coastal Plain) (Fig. 2).
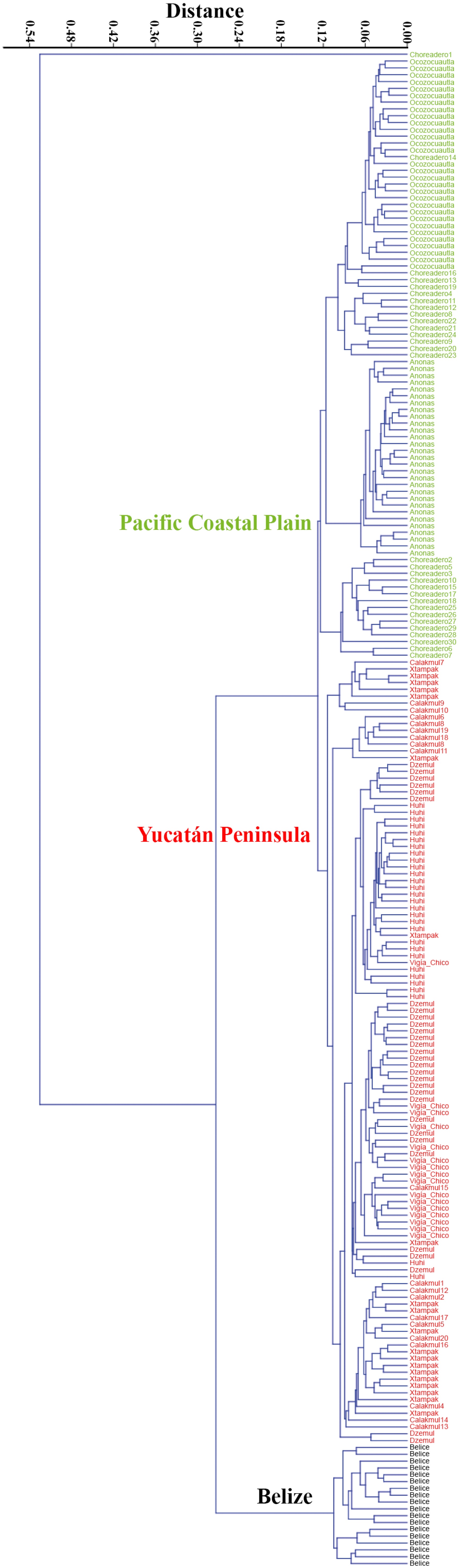
Figure 2 Dendrogram of 224 specimens of Gymnopodium floribundum Rolfe, based on a pairwise distance matrix from quantitative and qualitative characters and clustered with the UPGMA grouping method. Colors and labels show the geographic pattern of the three main clusters used in multivariate analyses of variance.
The PCA ordination analyses of the generalized variance in the distance data set showed that the first three components explain 52.9% of variance of the data (Fig. 3). These are those that turned out to be morphologically relevant according to the rule of the broken bar. The variables correlated with the first component were the distances between the floral fascicles (DF1, DF2, DF3), length and width of the outer perianth segments (OPL, OPW), the length of the achene (ACL) and the length of the accrescent style in the fruit (STLF). For the second component these were the length of the racemes (RAL) and length of the peduncle (PEDL) and for the third component the length of the petiole (PEL), as well as the length and width of the leaf blade (LBL, LBW) (Table 2).
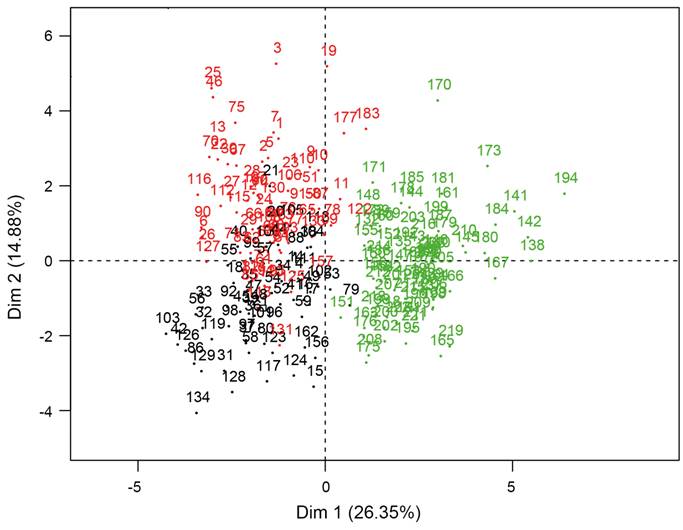
Figure 3: Multivariate morphological variation in the set of four measurements from vegetative characters and seventeen distances from reproductive characters of Gymnopodium floribundum Rolfe. Ordination diagram based on the scores of 224 specimens in the first two principal components showing the three clusters found in the UPGMA analyses: Cluster 1=Belize (black numbers), Cluster 2=Yucatán Peninsula (red numbers) and Cluster 3=Pacific Coastal Plain (green numbers).
Table 2: Correlation values obtained of the most important morphological characters (bold) of the first three principal components (PC) for Gymnopodium floribundum Rolfe.
| Morphological characters (mm) | PC 1 | PC 2 | PC3 |
| 1. Brachyblast length (BRL) | 0.04 | 0.18 | 0.51 |
| 2. Petiole length (PETL) | -0.03 | 0.35 | 0.6 |
| 3. Leaf blade length (LBL) | 0.34 | 0.34 | 0.68 |
| 4. Leaf blade width (LBW) | 0.23 | 0.38 | 0.76 |
| 5. Raceme length (RAL) | -0.2 | 0.74 | -0.31 |
| 6. Peduncle length (PEDL) | -0.47 | 0.6 | -0.02 |
| 7. Distance between the first and second flowering fascicle (1-2) (DF1) | -0.63 | 0.48 | -0.14 |
| 8. Distance between the second and third flowering fascicle (2-3) (DF2) | -0.56 | 0.54 | -0.11 |
| 9. Distance between the third and fourth flowering fascicle (3-4) (DF3) | -0.59 | 0.49 | -0.17 |
| 10. Length from the pedicel base to the articulation (PBA) | 0.55 | 0.16 | -0.35 |
| 11. Length from the pedicel articulation to the base of the perianth (PDI) | 0.34 | 0.22 | -0.4 |
| 12. Outer perianth segments length (OPL) | 0.74 | 0.37 | -0.12 |
| 13. Outer perianth segments width (OPW) | 0.72 | 0.36 | -0.18 |
| 14. Inner perianth segments length (IPL) | 0.66 | 0.28 | 0.03 |
| 15. Inner perianth segments width (IPW) | 0.42 | 0.25 | -0.15 |
| 16. Ovary length (OVL) | -0.001 | -0.002 | -0.01 |
| 17. Ovary width (OVW) | 0 | -0.001 | 0 |
| 18. Style length (STL) | 0 | 0 | -0.005 |
| 19. Achene length (ACL) | 0.72 | 0.18 | -0.04 |
| 20. Achene width (ACW) | 0.61 | 0.24 | -0.08 |
| 21. Style length in fruit (STLF) | 0.73 | -0.19 | -0.1 |
Figure 4 shows the graphs of the means and confidence intervals at 95% of the 15 morphological characters that showed the highest correlation with the first three principal components. The morphological groups characters, such as the length of the leaf blade, width of the inner perianth segment, length of the brachyblast, length of the accrescent style in the fruit, length and width of the fruit ,have means and intervals that do not overlap among the three morphological groups from Belize, Yucatán Peninsula, and the southern Pacific.
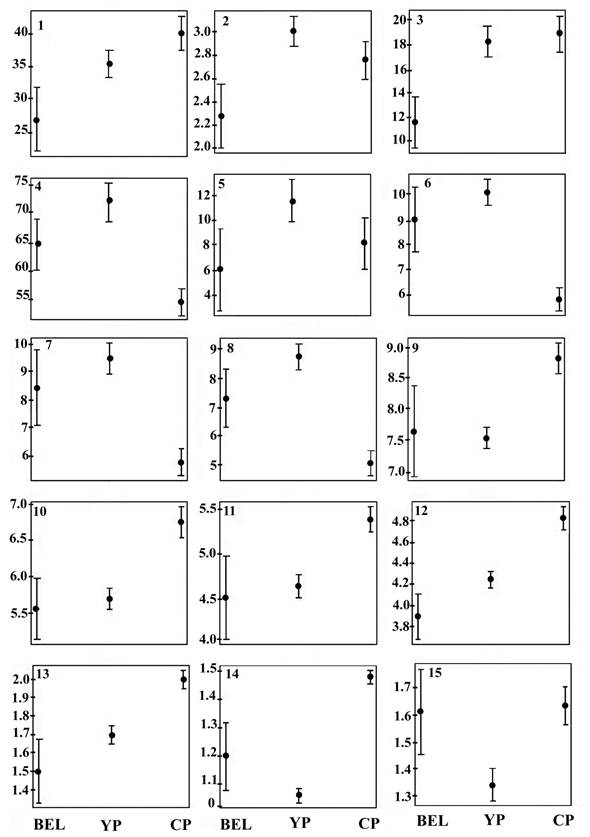
Figure 4: Contribution of morphological characters to the variation and differentiation of three geographical groups within Gymnopodium floribundum Rolfe. Mean graphs and confidence intervals at 95% for the fifteen highly correlated characters: 1=LBL, 2=PETL, 3=LBW, 4=RAL, 5=PEDL, 6=DF1, 7=DF2, 8=DF3, 9=OPL, 10=OPW, 11=IPL, 12=ACL, 13=ACW, 14=STLF, 15=PBA. See Table 2 for character description. X axis: Cluster 1=BEL, Cluster 2=YP and Cluster 3=CP. Y axis: distance in mm.
The multivariate analyses of variance (PERMANOVA) performed on the distance data set comparing the centroids of the three a priori groups (Yucatán Peninsula, Belize and the Pacific Coastal Plain), found significant differences between them (F=21.27, p<0.001 with 2, 220 g.l. and 9999 permutations). In terms of the multivariate dispersions of the data, there were no significant differences between the groups, so the hypothesis test was not significant (F=0.1079, p=0.8978 with 2, 220 g.l.).
Both multivariate analyses of leaf shape separately from perianth segment shape found significant differences among the three geographic “a priori” groups (Figs. 5A-B). In the geometric morphometric analyses of tepal shape of the outer perianth segment there were two significant canonical axes (Fig. 5A). Regarding the analyses of leaf blade shape, there were also two different statistically significant axes (Fig. 5B). Partial shape deformations explained by each significant canonical axes help to detect regions in the contours where there are more differences among the three groups. In the specimens from Belize, the shape of the leaf blade varies from oblong, elliptical to obovate showing a reduction of the apex and the base to an obtuse form. This group contained the smallest leaf blades from 20 to 32 mm long and from 8 to 13 mm wide. The Yucatán Peninsula specimens tend to widen at the leaf base and the middle part, while the apex varies from rounded to slightly emarginated. The leaf blades of the individuals of the Pacific Coastal Plain tend to be reduced both at the base and in the middle portion while the apex varies from acute to acuminate. The segments of the outer perianth present a similar pattern of differentiation, so in the Belizean specimens the perianth lobe tends to widen to the right, while in the specimens of the Yucatán Peninsula the perianth lobe does it towards the left. Finally, Pacific specimens have symmetrical lobes showing a circular shape at the base of the floral segment.
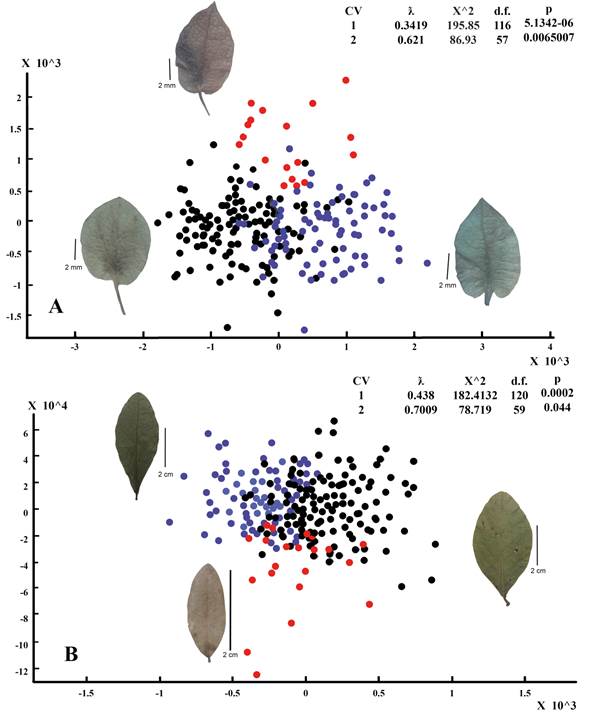
Figure 5: Multivariate analyses of variance of two shapes in three geographical groups within Gymnopodium floribundum Rolfe. These are the CVA ordinations from the partial warp scores estimated with IMP: CVAGen8. In the two geometric morphometric analyses, the three groups were significantly different. The statistics for the two canonical funtions are given in the table above each ordination. A. variation in the landmark coordinates of the shape of the outer perianth segments (Scale line 2 mm); B. variation in the landmark coordinates of the shape of the leaf blade (Scale line 2 cm). Belize (red dots), Pacific Coastal Plain (blue dots) and Yucatán Peninsula (black dots). CV: Canonical variates; ƛ: Wilk's Lambdas; X^2: Bartlett's test; d.f.: Degrees of freedom; p: p-valueless than 0.05.
Discussion
The treatment of quantitative morphological character data with a multivariate approach has been very useful to differentiate and to identify taxa in complexes of species in plants with apparently minimal morphological variation (de Luna and Gómez-Velasco, 2008; Pinzón et al., 2011). In the case of G. floribundum, morphometric characters have not been considered previously. The qualitative characteristics such as the presence or absence of indument beneath the leaf blade, as well as its shape, have been the characters used to delimit the species and varieties (Blake, 1921; Standley and Steyermark, 1946). The multivariate statistical exploration of distance data and the geometric morphometric analyses of the shape of the leaf blade and the outer perianth segment allow us to incorporate new characters in the taxonomic treatment of G. floribundum.
The grouping methods based on Gower´s coefficient and a UPGMA dendrogram produced groups of specimens congruent with a geographic arrangement in three major areas: Belize, Yucatán Peninsula, and the Pacific Coastal Plain (Fig. 6). The multivariate analyses of variance from the distance data showed significant differences between three morphological groups. This distinctive geographic pattern in morphological variation suggest a degree of infraspecific variation or incipient speciation (Mallet, 2007; Remsen, 2010). There are several vegetative and reproductive characters that reveal such differentiation among populations. For example, measurements of the floral fascicles, the length and width of the outer perianth segment, and the length of the inner perianth segment are characters that are different among the three geographic groups in Belize, the Pacific (Chiapas and Oaxaca), and the Yucatán Peninsula. Geometric morphometric analyses of the shape of the leaf and the perianth segment also showed differences between the three morphological groups. In summary, our morphometric analyses of distances and landmarks concur in the hypothesis of the existence of three well-defined morphological groups and geographical groups. This pattern of geographical structure suggest the hypothesis of three taxonomic groups at the level of subspecies.
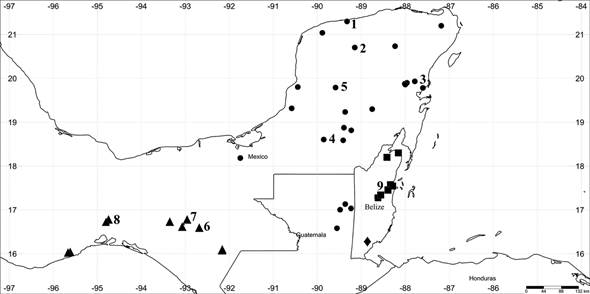
Figure 6: Distribution of the species and subspecies of Gymnopodium Rolfe: G. toledense Ancona & Ortiz-Díaz; ●G. floribundum subsp. antigonoides (Standl. & Steyerm.) Ancona & Ortiz-Díaz; pG. floribundum subsp. chiapensis Ancona & Ortiz-Díaz; ▪G. floribundum Rolfe subsp. floribundum. Numbers 1-9 are populations sampled in this study.
The most recent taxonomic treatment of Gymnopodium (Ortiz-Díaz, 1994) documented a high degree of variation in the indument and the shape of the leaf blade throughout its distribution range. Our findings reveal that the glabrous character of the ochreae, the petiole and the leaf blades of the specimens of Belize correspond to a taxonomic group previously described by Standley and Steyermark (1946) as G. floribundum var. floribundum. In turn, the presence of indument in the ochreae, petiole and leaf blades of the specimens from the Yucatán Peninsula correspond to G. floribundum var. antigonoides (Standley and Steyermark, 1946). Our morphometric analyses helped to reveal a third group including specimens from the Pacific coast which share a set of novel quantitative characters in the inflorescence, such as the reduction in the distance between the floral fascicules of the raceme. This group and characters have not been described in the previous treatments of Gymnopodium. We propose to formally recognize this geographic group at the subspecies rank.
The concept of subspecies was originally conceived at the end of the 19th century as a formal mean of documenting geographic variation or units of variation within species and replacing the term variety as a taxonomic range (Mallet, 2007). According to Mayr (1942; 1963), a subspecies is an aggregation of phenotypically similar populations of a species inhabiting a geographical subdivision within the overall range and differing from other such subdivisions of the species. Tobias and collaborators (2010) mention that the evidence of quantitative variation among the populations of a species should be considered as a subspecies until there is more genetic, ecological, phylogenetic, phylogeographic or behavioral evidence that allows the change of status at the species level.
A phylogenetic analysis of nuclear DNA (LFY and ITS) carried out by Ancona et al. (2018), to delimit G. floribundum of G. toledense, showed that specimens from Belize, the Yucatán Peninsula, and the Mexican Pacific Coast intermingled without forming monophyletic groups (phylogenetic species). This suggests that there is no genetic difference between these geographic regions as shown by morphological variation in this study. Traditionally, subspecies have been defined by morphological traits and geographical isolation without reflecting the underlying genetic structure and phylogenies (Zink, 1989; 2004). In the case of genetic congruence and the formation of monophyletic groups, the subspecies should be recognized as lineages, and therefore, their taxonomic status should be changed to the species level according to the phylogenetic concept of species (Mishler and Theriot, 2000).
The increase of samples of G. floribundum sensu lato and molecular data as well as new species delimitation methods could solve this problem. Based on our assembled character data set and morphometric analyses we propose to recognize the following three groups: G. floribundum var. antigonoides and G. floribundum var. floribundum as two subspecies, and a third new subspecies, Gymnopodium floribundum subsp. chiapensis is described for Chiapas and Oaxaca.
Taxonomic treatment
Key to the species of Gymnopodium
1a.. Leaves, inflorescence rachises and pedicels glabrous or covered with sparse to dense simple trichomes without basal glands; veins not prominent on abaxial surface of leaf blade; pedicel basal segment not hidden by ochreole; outer perianth segments 6.5-8 mm long, 5-6.5(-7) mm wide; inner perianth segments 4-5 mm long, 1-1.5 mm wide; fruit 4-4.5(-5) mm long, 1.5(-2) mm wide …………………...… Gymnopodium floribundum Rolfe
1b.. Leaves, inflorescence rachises and pedicels covered with sparse to dense simple trichomes with basal glands; veins prominent on abaxial surface of leaf blade; pedicel basal segment hidden by ochreole; outer perianth segments 8.5-9(-9.5) mm long, 6-6.5(-7) mm wide; inner perianth segments (5-)5.5-6 mm long, 1.5-1.8 mm wide; fruit 5(-5.5) mm long, 2(-2.5) mm wide ………………………… Gymnopodium toledense Ancona & Ortiz-Díaz
Gymnopodium floribundum Rolfe, Hooker´s Icon. 27(4): t. 2699. 1901. TYPE: BELIZE. Manatee, pine ridges, V.1900, E. J. F Campbell 60 (holotype: K000585032!, isotype: K000585033!).
= Millspaughia leiophylla S.F. Blake, Contr. Gray Herb. 52: 62. 1917. TYPE: BELIZE. Manatee Lagoon, in swampy saline ground, 30.I.1905, M. E. Peck 320 (holotype: GH00036641!, isotype: K000585034!).
Shrubs or trees 2-7 m tall; bark grey to dark brown fissured, young branches bivaricate, flexuous, sparse to dense pilose when young, glabrous when mature; trichomes simple; internodes 0.5-3 cm long; ochreas 1-2 mm long, deciduous, annular, glabrous to densely pilose; trichomes simple; leaves alternate, simple arising from the ochrea, or fascicled (2-3) arising on small vegetative shoots (brachyblasts); petiole 0.4-4 mm long, 1 mm wide, canaliculate, glabrous to densely pilose, dark brown; trichomes simple; leaf blades 2.7-9.5 cm long, (0.9-)2-5 cm wide, elliptic, oblong ovate to obovate, membranaceous to chartaceous, margin entire, apex obtuse, rounded to seldom emarginate, base cuneate to obtuse, abaxially glabrous to densely pilose; trichomes simple; nerves pinnate, not prominent; inflorescence 3-12 cm long, terminal; racemes single or paired, forming a compound panicle, on brachyblasts; rachis 3-14 cm long, glabrous to denseky pilose, trichomes simple; flowers hermaphrodite, born in fascicles of 2-6; ochreolae 1-2 mm long, lanceolate, membranous, sparse to densely pilosulose; trichomes simple; pedicels articulated, lower portion 1.3-1.9 mm long, upper portion 5-7.5(-8) mm long, pilosulose; perianth segments 6, 3 outer ovate-cordate, chartaceous, green to yellowish, sparse to densely pilose when young, glabrous or pilosulose when mature, nerves anastomosed, 3 inner subulate-lanceolate, acuminate, papery, glabrous to pilosulose, accrescent in fruit; perianth accrescent in fruit, 3 outer 6.5-8 mm long, 5-6.5(-7) mm wide, 3 inner 4-5 mm long, 1-1.5 mm wide, nerves reticulate; stamens 9, 6 outer inserted on a basal nectariferous disc, 3 inner arising opposite to ovary sulcus, filaments 2 mm long, anthers suborbicular, 0.5-0.7 mm long, versatile; ovary superior sessile, trigonous, dorsally compressed, 1-1.5 mm long, 0.5 mm wide, glabrous to pilosulose; styles 3, filiform, stigmas 3, capitate; fruit an achene, trigonous, 4-4.5(-5) mm long, 1.5(-2) mm wide, smooth, light brown, shiny, covered by the acrescent perianth segments; seed 1.
Distribution and ecology: Mexico (Campeche, Chiapas, Oaxaca, Quintana Roo, Tabasco, Yucatán), Belize and Guatemala (Petén). Tropical dry forest and savannas, 8-1350 m elevation.
Phenology: this species flowers mainly from March to May, but sporadic blooms occur between November and December.
Common names: sak ts'its'il che', ts' iits' il che', xts' iits' il che' (Yucatán), bastard logwood (Belize).
Key for the identification of subspecies in Gymnopodium floribundum
1a.. Ochrea, petiole and abaxial surface of leaf blades glabrous ….. Gymnopodium floribundum Rolfe subsp. floribundum
1b.. Ochrea, petiole and abaxial surface of leaf blade sparsely to densely pubescent ………………… 2
2a.. Distance between floral fascicles 8.5-10.5 mm; tepals of the outer perianth segments, 7.3-7.6 mm; achene 3.5-4.5 mm long …………… Gymnopodium floribundum subsp. antigonoides (Standl. & Steyerm.) Ancona & Ortiz-Díaz
2b.. Distance between floral fascicles 4.5-6 mm long; tepals of the outer perianth segments, 8.5-9 mm long; achene 4.5-5 mm long ……………….. Gymnopodium floribundum subsp. chiapensis Ancona & Ortiz-Díaz
Gymnopodium floribundum Rolfe subsp. floribundum, stat. nov.
Shrubs or rarely trees, 2-4 m tall; ochrea 1-2 mm long, glabrous; petioles 2-2.5 mm long, glabrous; leaf blades 2-3.5 cm long, 0.8-1.3 cm wide, elliptic-oblong, chartaceous, apex obtuse to rounded, abaxial surface glabrous; raceme 4.5-6.8 cm long; distance between floral fascicles 6.5-10 mm; pedicels 4.5-9 mm long, articulated, lower portion 1.3-1.8 mm long, upper portion 4-5.5 mm long; flowers hermaphrodite; perianth segments 6, 3 outer 7-8 mm long, 4.5-5.5 mm wide, chordate, 3 inner, subulate-lanceolate, acuminate, 4-5 mm long, 0.5-1 mm wide; fruit an achene, trigonous, 3.5-4 mm long, 1.5-1.7 mm wide.
Distribution and ecology: Gymnopodium floribundum subsp. floribundum is restricted to the seasonal savannas of Belize (Fig. 6). It is distributed between 8-20 m elevation.
Additional specimens examined (n=23): BELIZE. District Belize, Indian church, sabana across lagoon, 30.V.1976, T. Arnason and J. Lambert 17165 (MO); 12.5 miles northwest of Belize along northern highway, marshy savannas, 6.VI.1973, T. B. Croat 23257 (MO); ridge Lagoon plantation, ca. 12 miles NW of Belize, 9.VI.1973, T. B. Croat 24018 (MO); 20 mi W of Belize on western highway, 24.III.1967, J. Dwyer et al. 633 (MO); savanna near Hattieville, 5.VII.1972, J. D. Dwyer 10091 (MO); mile 7.5, northern highway, thicket aside Belize River, 25.V.1974, J. D. Dwyer 12397 (MO); mile 11, western highway, 4.VI.1974, J. D. Dwyer 12666 (MO); mile 29, northern highway, 24.III.1973, J. D. Dwyer and L. Dieckman 10447 (MO); mile 19.5, northern highway, 25.I.1974, J. D. Dwyer and R. Liesner 12203 (MO); pine ridge, highway Belice - Cayo, 15.IV.1958, P. H. Gentle 9714 (MO); maskall pine ridge, 24.II.1934, P. H. Gentle and C. L. Lundell 1143 (MO); 1.5 miles east of Hattieville, on coastal pine savanna, 5.VII.1972, J. S. Huston 566 (MO); 28.3 m from Belize, western highway, opposite link to old Belize road, 50 feet, 17°22'N, 88°32'W, 2IV.1979, R. R. Innes 220 (MO); Gracie Rock, Sibun river, 12.IV.1935, C. L. Lundell 1586 (MO); western highway, mile 15, lower pine ridge vegetation, 18.IX.1980, C. Whitefoord 2527 (BM, MO); western highway, mile 31, 3.V.1981, C. Whitefoord 2710 (BM); western highway, 12.5 miles west from Belize city, pine ridge savannah, 10.VIII.1970, J. R. Wiley 192 (MO, UADY). District Cayo, pineland in vicinity of Privaccion Creek, mountain pine ridge ca. 12 mi S Cayo, 15.IV.1972, D. Burch 5861 (MO); 28 miles from the highway, savannah of the Tropical Education Center (Belize Zoo), 17°21'20"N, 88°32'46"W, 10.V.1997, G. Davidse and D. L. Holland 36347 (MO, UADY); mai fire lookout tower, near Hilltop, III.1967, J. D. Dwyer et al. 215 (MO); 11-12 millas al norte de Baxing Pot, G. R. Proctor 30230 (MO). District Corozal, cerros maya ruins, Lowry's Bight, coastal area, 22.II.1983, C. J. Crane 270 (MO); vicinity of Little Belize, 18°12'N, 88°24'W, 17.III.1987, G. Davidse and A. E. Brant 32603 (MO); ca. 4 km SE of Sarteneja adjoining lagoon, 18.III.1987, G. Davidse and A. E. Brant 32690 (MO).
Gymnopodium floribundum subsp. antigonoides (Standl. & Steyerm.) Ancona & Ortiz-Díaz, stat. nov.
≡ Millspaughia antigonoides B.L. Rob., Bot. Jahrb. 36: 14. 1905. TYPE: MEXICO. Yucatán, Progreso, 5.III.1899, C. F. Millspaugh 1657 (lectotype: F! designated here, isolectotype: GH!).
= Millspaughia ovatifolia B.L. Rob., Bot. Jahrb. 36: 14. 1905. TYPE: MEXICO. Yucatán, Progreso, 5.III.1899, C. F. Millspaugh 1672 (holotype F!, isotype GH!).
≡ Gymnopodium antigonoides (B.L. Rob.) S.F. Blake, Bull. Torrey Bot. Club 48(3): 84. 1921.
= Gymnopodium ovatifolium (B.L. Rob.) S.F. Blake, Bull. Torrey Bot. Club 48(3): 84. 1921.
≡ Gymnopodium floribundum var. antigonoides (B.L. Rob.) Standl. & Steyerm. Publ. Field Mus. Nat. Hist., Bot. Ser. 23(1): 5. 1943.
Trees or rarely shrubs, 2-7(-12) m tall; ochrea 1-2 mm long, sparse to densely pubescent; petioles 2.8-3.1 mm long, sparsely pubescent; leaf blades 3.5-3.7(-4.5) cm long, 1.7-2(-3) cm wide, obovate, papery, apex rounded, abaxial surface sparsely to densely pubescent; racemes 7-8 cm long; distance between floral fascicles 8.5-10.5 mm; pedicels 5.7-6.5 mm long, articulated, lower portion 1.2-1.4 mm long, upper portion 4.5-5.2 mm long; perianth segments 6, 3 outer, 7.3-7.6 mm long, 5.5-6 mm wide, cordate, 3 inner 4-5 mm long, 1.1-1.4 mm wide, subulate-lanceolate, acuminate; fruit an achene, trigonous, 4.1-4.5 mm long, 1.7-1.8 mm wide.
Distribution and ecology: Gymnopodium floribundum subsp. antigonoides is restricted to the dry forests of Mexico (Campeche, Quintana Roo, Tabasco, and Yucatán) and Guatemala (El Petén) (Fig. 6). It is distributed between 60-260 m elevation.
Additional specimens examined (n=91): GUATEMALA. District Petén, vicinity of archeological camp on north shore of lake Yaxha, 18.VI.1973, T. B. Croat 24626 (MO); lake Petén Itzá, bordering lake, about 12 km west of Remate, 23.V.1960, C. L. Lundell 989 (MO); Tikal National Park, Tikal, bordering airfield, 20.VI.1962, C. L. Lundell 17210 (MO); camino para San Andrés, 12 km de Santa Elena, 19.III.1970, R. Tún Ortíz 802 (MO); camino para Zocotzal, a 55 km, parque nacional Tikal, 10.VI.1971, R. Tún Ortíz 1828 (MO); NW-Umgebung des Lago Petén Itzá, gestörter Wald-Rest am NE-Abhang und Sekundär-Vegetation am Gipfel des Chakmamantok-Felsen, das ist 0.5 km NNE Zentrum von San José, 16°59'11"N, 89°53'54"W, 22.XI.1994, B. Wallnöfer 9487 (MO). MEXICO. Campeche, municipality Calakmul, D. Álvarez M. 4381 (MO); sobre el camino a Pixoyal, a 30 km al N de Escárcega, 18.IV.1982, E. F. Cabrera C. and H. de Cabrera 2394 (BM, MO); a 30 km al sur de Xpujil, 22.IV.1982, E. F. Cabrera C. and H. de Cabrera 2477 (MO); 5 km al S de Conhuás centro ceremonial de Calakmul, 27.III. 1988, E. F. Cabrera C. and H. de Cabrera 15990 (MEXU, MO); a 31 km al sur de la caseta de vigilancia hacia las ruinas, 25.XI.1997, E. M. Lira et al. 523 (MO); km 57 camino Escalache, 18°59'46"N, 89°17'00"W, 12XI.1997, E. Madrid et al. 421 (MO); 3 km al S de Xcan-ha camino a Xpujil, 19°5'15"N, 89°19'48"W, 30.IV.1997, E. M. Martínez S. and A. M. Pascual 27050 (MO); a 3 km al NE de Narciso Mendoza, 18°15'00"N, 89°26'20"W, 12.III.1998, E. M. Martínez et al. 30337 (MO); a 3 km de Zoh - Laguna, camino a Dzibanchén, 18°37'N, 89°25'W, 1.IV.1996, A. M. Pascual and E. M. Martínez S. 269 (MO). Municipality Calkiní, 4 km al O de Tuncasché, sobre el camino Punta Arenas, 1.XII.1988, E. F. Cabrera C. and H. de Cabrera 15242 (MEXU, MO). Municipality Campeche, Ciudad de Campeche, 24.X.1984, A. Espejo et al. 1207 (MO); Tulia 36 km al NE de Campeche camino a Mérida, 6.II.1983, E. M. Martínez S. and O. Téllez 3004 (MO). Municipality Champotón, ejido Villa Morenos, 19°31'36"N, 90°41'53"W, 23.IV.1985, C. Chan 5040 (MO). Municipality Hopelchén, A 9.28 km al NNO de Bel-ha, 19°00'25"N, 89°17'27"W, 29.V.2004, D. Álvarez 8846 (MO); a 1 km al sur de Bolonchén, 31.III.1982, E. F. Cabrera C. and H. de Cabrera 2370 (MO); Camino blanco entre Ucum a Xk'anja, 19°09'25"N, 89°19'40"W, 9.V.1984, Chan V. 3605 (MO); Bolochén de Rejón, along hwy. 180, S side of town, low deciduous forest, 18.V.1982, G. Davidse et al. 20596 (BM, MO). Quintana Roo, municipality Benito Juárez, Cancún, 21°8'7.08"N, 86°49'9.14"W, 13.II.2001, I. Miranda 31 (UADY). Municipality Cozumel, a 1 km al N del Faro de la punta celarain, 14.I.1986, E. F. Cabrera C. and H. de Cabrera 10564 (MEXU, MO); a 2 km al norte de la carretera transversal, siguiendo las brechas del agua potable, 13.III.1986, E. F. Cabrera C. and O. Téllez V. 11065 (MO). Municipality Felipe Carrillo Puerto, C. P. Cowan et al. 5104 (MEXU, MO); Sian Ka'an Biosphere Reserve, 15-20 km N of Carrillo Puerto, 19°50'N, 87°40'W, 1.II.1984, D. Neill et al. 5753 (MO). Municipality José María Morelos, Dziuché, laguna de Chichancanab, camino Presumida - Santo Tomas 50 m, 26.III.1984, J. J. Ortiz 464 (MEXU, MO). Municipality Lázaro Cárdenas, reserva El Edén, 21°28'N, 87°31'W, 1.IV.1993, J. S. Flores 12381 (UADY); reserva ecológica El Edén, 7 km al N de Leona Vicario, 21°04'N, 87°11'W, 3.III,1994, A. Gómez Pompa 5 (MO); reserva El Edén, estación La Sabana, 21°12.51'N, 87°11.64'W, 23.III.1996, B. Pitzer and E. Misquez 2450 (UADY); reserva El Edén, 21°11.3'N, 87°11.51'W, 3.IV.1996, B. Pitzer and E. Misquez 2655 (UADY); región de Yalahau, reserva El Edén, 21°12'N, 87°11'W, 14,II,1999, G. P. Schultz and R. A. Palestina 1067 (MO). Municipality Puerto Morelos, entrada de la brecha a Vallarta, 7.XI.1980, E. F. Cabrera C. and L. Cortés A. 135 (BM, MO). Municipality Solidaridad, X'cacel - X'cacelito, aprox. 13 km al norte de Tulum sobre la carretera federal 307 Chetumal - Puerto Juárez, estas bahías colindan al norte con las de Chemuyil y al sur con las de Xel-ha, 15.VI.1998, C. Gallardo et al. 2252 (MEXU, MO); a 20 km de Tulum, 5.IV.1981, T. P. Ramamoorthy et al. 2081 (MEXU, MO); Álvaro Obregón, 7.IV.1981, T. P. Ramamoorthy et al. 2162 (MEXU, MO); a 9 km a sur del entronque o a 21 km al W de Ucum, 5.III.1980, O. Téllez V. and E. F. Cabrera C. 1697 (BM, MO); a 1 km al norte de Playa del Carmen, 9.III.1980, O. Téllez V. and E. F. Cabrera C. 1800 (BM, MO). Municipality Tulum, Cobá, roadside north of Cobá ruins, 20.II.1987, D. A. White 297 (UADY). Tabasco, municipality Balancán, carretera al campamento San Pedro por el km 34, 6.IV.1976, J. I. Calzada 2350 (MEXU, UJAT); Balancán, L. E. Matuda 3182 (MEXU). Yucatán, municipality Akil, Carretera a Chetumal, km 102, 20°15'56"N, 89°33'5"W, X.1994, V. M. Navarro 22 (UADY). Municipality Buctzotz, Cenote Azul, a 4.5 km en la carretera a Yalsihón, 21°13'6.53"N, 88°40'7.66"W, 28.II.2004, B. S. Bolívar 13 (UADY). Municipality Cacalchén, V.1917, G. F. Gaumer and Sons 23874 (BM, MO). Municipality Conkal, a 2 km del suroeste de Conkal, 21°03'30"N, 89°31'55"W, 5.VI.1984, R. Rivera 42 (CICY, UADY); 70 (CICY, UADY). Municipality Dzilam de Bravo, reserva ecológica Bocas de Dzilam, 21°27'53.28"N, 88°31'33.01"W, 22.VII.1993, J. J. Ortiz 1796 (UADY). Municipality Hocabá, Sahcabá, 20°41'N, 88°23'W, 7.VII.1993, A. Mizrahi 12 (UADY); Sahcabá entre Xocchel y Huhi, 20°45'N, 89°10'W, 4.V.1994, F. J. Xuluc 04 (UADY). Municipality Hoctún, a 4.5 km de Tahmek, 20°52'N, 89°13'W, 31.VII.1972, G. L. Webster and S. P. Lynch 17569 (MO). Municipality Hunucmá, 15 km E of Celestún, 7.I.1983, S. P. Darwin 2442 (MO); reserva El Palmar, 21°05'30,23"N, 89°59'11.39"W, 20.II.2007, J. J. Ortiz and G. Palma 2650 (UADY). Municipality Izamal, Izamal, 1985, G. F. Gaumer 504 (MO). Municipality Mérida, carretera a Dzibilchaltún, km 3, 20°05'N, 89°26'W, 19.I.1984, J. J. Ortiz 415 (UADY); carretera San José Tzal - Molas, 8.X.2005, G. Pech and C. Guevara 05 (UADY); carretera San José Tzal - Tzununcan, 8.X.2005, G. Pech 02 (UADY); Dzibilchaltún, 19.I.1979, A. S. Bradburn and S. P. Darwin 1213 (BM, MO); Dzoyaxche, Reserva Ecológica Cuxtal, 20°46'23.9"N, 89°35'2.6"W, 1.VII. 2008, T. Andueza and T. Canul 19 (UADY); loc. cit., 1.VII.2008, F. Duarte and A. Pereira 23 (UADY); loc. cit., 1.VII.2008, C. Méndez and E. Pérez 17 (UADY); loc. cit., 19.IX.2004, A. Sánchez and G. Can 17 (UADY); loc. cit., 1.VII.2008, F. Zapata 09 (UADY); ejido Dzidzilché, 20°42'N, 88°14'W, 16.II.1985, E. Ucán 3735 (AUDY, CICY); Xmatkuil, FMVZ, 20°48'N, 89°47'W, 19.XI.2004, B. Ávila 10 (UADY); loc. cit., 20°58'N, 89°38'W, 19.II.1997, L. Casanova 20 (UADY); loc. cit., 20°52'N, 89°37'W, 12.XI.1996, L. G. Espinoza 1 (UADY); loc. cit., 13.II.2002, E. Hau 14 (UADY); loc. cit., 20°51'N, 89°37'W, 18.XI.2003, L. Mendicuti and O. Zamora 05 (UADY). Municipality Opichén, grutas de Calcethok, 20°33'N, 89°48'W, 17.III.1984, J. J. Ortiz 436 (MEXU, MO). Municipality Oxkutzcab, a 3 km al O de Sayil, sobre la carretera a Oxkutzcab, 20°10'12"N, 89°37'12"W, 20.IV.1986, E. F. Cabrera C. and H. de Cabrera 11318 (MO). Municipality Tecoh, Xcanchakan, a 2 km dirección al sur, 20°35'N, 89°32'W, 11.V.1997, I. G. Rodríguez 101 (UADY). Municipality Tekit, Tekit, 20°32'N, 89°20'W, 22.I.1994, M. E. Magaña 147 (UADY). Municipality Tinum, en los alrededores de las grutas de Balancanché, a 36 km al O de Valladolid, 2.X.1985, E. F. Cabrera C. and H. de Cabrera 9725 (MO); jardín botánico de Balancanché, 9.XII.1989, S. Escalante 722 (CICY, UADY). Municipality Uayma, carretera Uayma - Tinum a km 5, 20°47'N, 88°23'W, 26.II.1990, E. Ucán 5809 (UADY). Municipality Valladolid, a 14 km al O de Chemax, carretera Cancún - Valladolid, 16.VII.1985, E. F. Cabrera C. and H. de Cabrera 8897 (MO); Pixoy, 20°42'N, 88°15'W, 6.I.1987, E. Ucán 4981 (UADY); Pixoy rumbo a Valladolid km 3, 20°42'N, 88°14'W, 26.II.1990, E. Ucán 5800 (UADY); Pixoy, San Miguel Arboledas, en parcela de la escuela R. Cházaro Pérez, 20°42'N, 88°14'W, 16.IV.1990, E. Ucán 6035 (UADY). Municipality Yaxcabá, Yaxcabá, 20°31'N, 88°50'W, 21.XII.1993, M. E. Magaña 175 (UADY).
Gymnopodium floribundum subsp. chiapensis Ancona & Ortiz-Díaz, subsp. nov. TYPE: MEXICO. Chiapas, Tuxtla Gutiérrez, camino al parador turístico El Chorreadero, 16°45'4.57"N, 92°58'12.62"W, J. J. Ancona y J. J. Ortiz 178 (holotype: UADY!, isotypes: CICY!, CHIP!, MEXU!). (Figs. 7A-B).

Figure 7: A-B. Type of Gymnopodium floribundum subsp. chiapensis Ancona & Ortiz-Díaz; comparison of the racemes of: B. Gymnopodium floribundum subsp. chiapensis Ancona & Ortiz-Díaz; C. Gymnopodium floribundum Rolfe subsp. floribundum (G. R. Proctor 30230 (BM)); D. Gymnopodium floribundum subsp. antigonoides (Standl. & Steyerm.) Ancona & Ortiz-Díaz (Quijano et al. 919 (UADY)). Scale: 25 mm.
Gymnopodium floribundum subsp. chiapensis is similar to G. floribundum ssp. antigonoides due to the pubescence in the leaves, petiole and ochrea. This taxon can be distinguished from G. floribundum subsp. floribundum and G. floribundum subsp. antigonoides by the smaller distance between the floral fascicles of the raceme (4.5-6 mm), giving the infloresence a densely agglomerated appearance (Figs. 7B-D).
Trees 2-7 m high; ochreae 1-2 mm long, sparsely to densely pubescent; petioles 2.6-2.8 mm long, sparsely pubescent; leaf blades 3.8-4.4(-7) cm long, 1.7-2.3(-4) cm wide, elliptic, membranous, apex acute-acuminate, sparsely to densely pubescent; racemes 6-7 cm long; distance between floral fascicles 4.5-6 mm; pedicels 7.5-8.5(-9) mm long, articulated, lower portion 1.5-1.7 mm long, upper portion 6.5-7.5 mm long; hermaphrodite flowers; perianth segments 6, 3 outer, 8.5-9 mm long, 6.5-7 mm wide, rounded, 3 inner 5-5.5 mm long, 1.4-1.5 mm wide, subulate-lanceolate, acuminate; fruit an achene, 4.5-5 mm long, 1.9-2 mm wide, trigonous.
Etymology: the subspecies epithet refers to the state of Chiapas, region where this taxon has been collected most abundantly.
Distribution and ecology: Gymnopodium floribundum subsp. chiapensis is restricted to the dry forests of Chipas and Oaxaca (Fig. 6). It is distributed from 60 to 1350 m elevation.
Additional specimens examined (n=28). MEXICO. Chiapas, municipality Chiapa de Corzo, steep walled canyon, above El Chorreadero, 16.IV.1972, D. E. Breedlove 24575 (BM, MO); Edge of cliff, at the Rio Grijalva, 10 km west of Chiapa de Corzo along mexican highway 190, 16.V.1972, D. E. Breedlove 25170 (MO); above El Chorreadero, steep walled canyon with tropical deciduous forest, 18.III.1981, D. E. Breedlove 50173 (MO). Municipality Chicoasen, mirador Manos que imploran, cañón del sumidero, O. Farrera 1919 (CHIP); arroyo San Antonio, 26.V.2009, H. Gómez-Domínguez 2055 (HEM); en la rivera de la represa Bombaná, aproximadamente a 9 km del ejido Chicoasén, 16°58'34.7"N, 93°02'53.7"W, 7.V.2009, O. J. Martínez-Meléndez 2057 (HEM); El Aguacero, 17.IV1991, RGGA 4 (HEM); en la carretera P. H. Manuel M. Toorres - Chicoasén, A. Márquez et al., 18 (HEM); a 2 km al O de la estación meteorológica de CFE sobre el camino de terracería, 21.IV.2009, 16°57'53.3"N, 93°08'24"W, O. J. Martínez-Meléndez et al. 2016 (HEM); 2 km al NO de la estación metereológica de CFE, 5.V.2009, N. Martínez-Meléndez 2617 (HEM). Municipality Jiquipilas, ejido Quintana Roo, 16°36'N, 93°33'W, 14.IV.1995, Farrera 698 (HEM); cerro ubicado al NE del poblado de Chicoasen, a 500 metros del puente (carretera Chicoasen - Copainalá), 15.IV.2009, A. López-Cruz 629 (HEM, MO). Municipality Ocozocuautla de Espinosa, rancho La Cabaña, 25.V.2002. 16°43'00"N, 93°28'11"W, A. Ávila-Solís 3 (HEM); 2 km antes de Ocozocuautla, carretera Tuxtla Gutiérrez - Ocozocuautla, 29.III.1987, S. Hernández and A. Espejo 208 (MO); Ocozocuautla, 1 km al NW del entronque aeropuerto - Ocozocuautla sobre la carretera 190, 9.V.1988, A. Reyes-García 558 (BM). Municipality Tuxtla Gutiérrez, 6.5 km west of Tuxtla Gutiérrez along Mexican highway 190, 8. X.1971, D. E. Breedlove 20097 (MO); Terán, Canyon, 4 km N of Juan Crispin along road to San Fernando, 17.XII.1972, D. E. Breedlove 30391 (MO); cañada La chacona al NW de Tuxtla sobre el cauce del arroyo temporal, R. Gallegos-Ramos 28 (CHIP); Trapichito Comitán, 2.VI.1945, L. E. Matuda 5698 (MO); Cañón del Sumidero National Park, Near Mirador La Ceiba, km 7 of Sumidero road, 16°47'N, 93°06'W, 15.III.1983, D. Neill 5518B (MO); 4 km al SE de Tuxtla Gutiérrez, camino a Villaflores, 06.IV.1983, O. Téllez y J. L. Villaseñor 6583 (BM, MEXU, MO). Oaxaca, municipality Juchitán de Zaragoza, San Miguel Chimalapa, 13.III.1982, R. Cedillo y R. Torres 1166 (MEXU, MO); 8 km al SE de Vista Hermosa hacia San Miguel Chimalapa, 13.III.1982, R. Torres y R. Cedillo 109 (MO). Municipality San Pedro Huamelula, al norte de Santiago Astata, 15°5846.92"N, 95°36'58.3"W, 29.X.2002, M. Elorsa C. 6269 (MEXU); La Mishi, 16°1'32.23"N, 95°39'52.35"W, 28.IV.2009, J. Leyva Márquez 73 (MEXU). Municipality Santigo Astata, a 8 km de Vista Hermosa, 15°59'24.14"N, 95°40'45.08"W, 26.VI.2009, E. Lott y A. Sánchez 5889 (MEXU).
Gymnopodium toledenseAncona & Ortiz-Díaz, Willdenowia 48: 433-441. 2018. TYPE: BELIZE. Toledo, Las Sierritas, 20 km W of Big Creek Settlement, ridge and W slopes of Cerrito in Las Sierritas hills, 16°31'45"N, 88°36'05"W, 160-213 m, ridge-top vegetation of mixed hardwood species growing on thin soils over exposed limestone, vegetation severely damaged by recurrent fires, 6.XII.1997, T. Hawkins 1681 (holotype: MO321695!, isotypes: BM000565699!, MEXU898235!).
Shrubs often scrambling, 2-4 m tall; bark grey to dark brown, fissured; young branches bivaricate, flexuous, grey to pale brown; internodes 2-3 cm long; ochrea ca. 1 mm long, deciduous, annular, sparsely pubescent; trichomes with basal glands; leaves alternate, simple, arising from ochrea, fasciculate (2 or 3 together) on small vegetative shoots (brachyblasts); petiole 1.5-2 mm long, ca. 1 mm wide, canaliculate, dark brown, densely pubescent; trichomes with basal glands; leaf blade 5-7 cm long, 3-4 cm wide, obovate to obpyriform, chartaceous, base obtuse, margin entire, apex obtuse to slightly emarginated, abaxial surface densely pubescent, adaxial surface glabrous except puberulent on midvein; trichomes with basal glands; veins prominent abaxially; inflorescence 12-20 cm long, terminal; racemes single or paired, on brachyblasts; rachis 15-18 cm long, densely pilose; trichomes with light yellow basal glands; flowers in fascicles of 2-4(-6); ochreoles lanceolate, 1-2 mm long, membranous, sparsely to densely pubescent; trichomes with basal glands; pedicels articulated, lower portion 0.5-1 mm long, upper portion 5-6.5(-8) mm long, densely pubescent; trichomes with basal glands; flowers hermaphrodite; perianth segments 6, 3 outer ovate-cordate, chartaceous papery, green to yellowish, sparse to densely pilose when younge, glabrous or pilosulose when mature, accrescent in fruit, nerves anastomosed, 3 inner subulate-lanceolate, acuminate, papery, glabrous to pilosulose, accrescent in fruit; perianth accrescent in fruit, 3 outer 8.5-9(-9.5) mm long, 6-6.5(-7) mm wide, 3 inner (5-)5.5-6 mm long, 1.5-1.8 mm wide; nerves reticulate; stamens 9, 6 outer inserted on a basal nectariferous disc, 3 inner arising opposite to ovary sulcus; filaments 2 mm long; anthers suborbicular, 0.5-0.7 mm long, versatile; ovary 1 mm long, 0.5 mm wide, superior, sessile, trigonous, compressed, densely pubescent at vertices; trichomes with basal glands; styles 3, filiform, 1.5-1.7 mm long; stigmas 3, capitate; fruit an achene, light brown, lustrous, trigonous, 5(-5.5) mm long, 2(-2.5) mm wide, smooth, included in perianth segments; seed 1.
Distribution and ecology: Gymnopodium toledense is so far known as an endemic species of the seasonal forests of southern Belize (Fig. 6), in the biogeographic region of Eastern Central America. It could possibly also be found in Guatemala and Honduras.
Phenology: this species has been collected in flower in December.
Comments: in the morphological description of G. toledense, Ancona et al. (2018) erroneously describe the presence of 6 stamens, 3 external ones inserted in a basal disc and 3 internal ones that arise in the opposite sulcus of the ovaries. However, this information is corrected here, G. toledense presents 9 stamens, 6 outer ones inserted on a basal nectariferous disc, and 3 inner arising opposite to ovary sulcus.

