











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Castilleja tenuiflora Benth. es una planta herbácea perenne de la familia Orobanchaceae (Tank y Olmstead, 2008), conocida como hierba del cáncer, atzoyatl y cola de borrego, entre otros nombres (Hernández, 1943). Ha sido utilizada en la medicina tradicional mexicana desde el siglo XVI, en el tratamiento de tos, disentería, nervios, dolores menstruales e inflamación, cáncer y enfermedades gastrointestinales (Béjar et al., 2000).
En plantas silvestres y cultivadas in vitro se ha evaluado la actividad antiinflamatoria y antiulcerogénica en pruebas con ratones (Carrillo-Ocampo et al., 2013; Sanchez et al., 2013), se han reportado sus efectos citotóxicos (Moreno-Escobar et al., 2011), antioxidantes (López-Laredo et al., 2012) y antidepresivos (Herrera-Ruiz etal., 2015). Dicha actividad biológica está relacionada con metabolitos secundarios como iridoides glicosilados (aucubina y bartsiósido) (Jiménez et al., 1995; Carrillo-Ocampo et al., 2013), feniletanoides glicosilados (verbascósidos e isoverbascósidos) (Gómez-Aguirre et al., 2012; Sanchez etal., 2013), flavonoides (apigenina y quercetina) (López-Laredo et al., 2012) y lignanos (Herrera-Ruiz et al., 2015), que fueron identificados tanto en plantas silvestres, cultivadas in vitro, como en plantas aclimatadas en condiciones de vivero (López-Laredo et al., 2012).
La producción de semillas sintéticas, que consiste en estructuras vegetales de origen normalmente asexual que constan básicamente de un embrión encapsulado en un endospermo artificial formado por una matriz polimérica (Morales y Cano, 2012), puede ser una opción para prolongar el tiempo de almacenamiento y mantener el genotipo de plantas élite, facilitar el intercambio de germoplasma, contar con material disponible para cultivo ex vitro, impedir riesgos de contaminación y contrarrestar el saqueo de plantas de su hábitat natural, principalmente. Se ha utilizado la encapsulación en plantas como Centella asiatica (L.) Urban (Prasad et al., 2014), en análisis por cromatografía líquida de alta eficiencia (HPLC), las plantas obtenidas de semillas sintética expresaron uniformidad cuantitativa y cualitativa de los compuestos de interés como los centellósidos. Los agentes encapsulantes más utilizados en la elaboración de semillas sintéticas son el alginato de sodio en complejo con cloruro de calcio dihidratado, ambos en diferentes concentraciones y tiempos de polimerización (Standardi y Micheli, 2012; Sharma et al., 2013), dependiendo de la especie con que se trabaje (Gantait et al., 2015). La elaboración de semillas sintéticas ha permitido la propagación y preservación a largo plazo de embriones somáticos de orquídeas (Teixeira da Silva et al., 2014) y agaves (Chandrasekhara et al., 2012). Por otro lado, el encapsulado de tejidos como segmentos nodales de plantas con importancia frutícola (Rai et al., 2009) se ha aplicado en cítricos (Nieves et al., 1998) en plantas medicinales como Centella asiatica (Prasad et al., 2014), y en alfalfa (Medicago sativa L.) (Fujii et al., 1992). En caña de azúcar (Saccharum officinarum L.) variedad CP-5243, los embriones encapsulados se han plantado directamente en campo (Nieves et al., 2003).
Castilleja tenuiflora es comercializada en los mercados de plantas medicinales. Se extrae de su ambiente natural en zonas altas y semiáridas de México entre 2300 y 3300 m s.n.m. (Calderón y Rzedowski, 2001), áreas que cada día son amenazadas por el deterioro humano (Salcedo-Morales et al., 2009). Debido a las propiedades medicinales de C. tenuiflora, es importante desarrollar métodos que permitan su preservación por tiempos prolongados, así como el mantenimiento de líneas de subcultivo, que contengan los principales compuestos de interés, para su disposición.
La propagación y conservación de microestacas en cápsulas se propone como un método alternativo para preservar plantas élite y contrarrestar el saqueo de individuos de su hábitat natural; además de brindar protección directa al explante, es posible aumentar el tiempo de almacenamiento, reducir la frecuencia de subcultivos, prevenir riesgos de contaminación y de variación somaclonal (Sharma et al., 2013) y mantener líneas de subcultivo de la misma planta para su disposición. Por lo anterior, el objetivo en este estudio fue desarrollar un método para la encapsulación de microestacas de C. tenuiflora, que después del proceso de encapsulación y almacenamiento no afecte el perfil químico de la planta.
Materiales y Métodos
Obtención del material vegetal
Se utilizaron microestacas (segmentos de tallo no apicales con un nodo) obtenidas de una línea de plantas in vitro de C. tenuiflora proporcionada por el Laboratorio de Productos Naturales del Centro de Desarrollo de Productos Bióticos del Instituto Politécnico Nacional (CEPROBI-IPN). Esta línea se mantuvo mediante subcultivos de acuerdo con el método propuesto por Salcedo-Morales et al. (2009), para tener una fuente constante de material vegetal y producir las semillas sintéticas con los diferentes tratamientos. Las plantas fueron subcultivadas cada 21 días en medio MS (Murashige y Skoog, 1962) al 100% (4.4 g l-1) con ácido indol butírico y 6-bencilaminopurina (0.1 mg l-1 IBA y 0.25 mg l-1 BAP), el pH ajustado a 5.8 y como agente gelificante Bactoagar al 0.8% (p/v). Se utilizaron cajas magenta como contenedores y se colocaron nueve explantes por caja, incubadas a 25±2 °C con intensidad luminosa de 1800 Lux, con un fotoperiodo de 16/8 horas luz/obscuridad.
El tejido de encapsulación consistió en microestacas (con un nodo y 2 mm de tallo por cada extremo) de C. tenuiflora de 5 mm de longitud aproximadamente, provenientes de plántulas con 35 días de cultivo in vitro. Las fracciones de tallo útiles para la encapsulación fueron el segundo, tercero y ocasionalmente el cuarto nodo de las plántulas subcultivadas.
Producción de las cápsulas con microestacas
Materiales encapsulantes
Las cápsulas se elaboraron de acuerdo con el protocolo propuesto por Gantait et al. (2015) y Sharma et al. (2013), quienes recomiendan como agentes encapsulantes al alginato de sodio (REASOL) en complejo con cloruro de calcio dihidratado (FERMONT).
Preparación de los componentes de la matriz de encapsulación
Este método consistió en realizar una solución base con medio Murashige y Skoog (1962) (MS) al 100% (4.4 g l-1), adicionada con reguladores de crecimiento vegetal IBA (0.1 mg l-1) y BAP (0.25 mg l-1), sacarosa (3%), el pH ajustado a 5.8. En 300 ml de agua desionizada se adicionaron 1.3 g de MS, 33.8 µl de IBA, 30.5 µl de BAP y 9 g de sacarosa.
En matraces de 250 ml se colocaron 150 ml de la solución base y se adicionó el alginato de sodio para formar soluciones al 2, 3 y 4%, en otros matraces con solución base se adicionó el cloruro de calcio dihidratado con diferentes concentraciones (50, 75 y 100 mM).
Las mezclas fueron sometidas a tres tiempos de polimerización (20, 25 y 30 min), para obtener distintos grados de dureza (Sharma et al., 2013; Gantait et al., 2015) (Cuadro 1). Los matraces de las seis soluciones con las diferentes concentraciones de alginato de sodio y cloruro de calcio dihidratado fueron esterilizadas en autoclave de tres calores (AESA, modelo CV 300, serie 1577, Cd. Mx., México) a 15 lb de presión por 20 min.
Cuadro 1: Combinación y concentració de los agentes encapsulantes alginato de sodio (alg-Na %) y cloruro de calcio dihidratado (CaCl2 2H2O mM), en diferentes tiempos de polimerización.
| Agentes encapsulantes | Concentración utilizada | Tiempo de polimerización (min) | |
| Alginato de sodio (p/v) CaCl2 2H2O (mM) |
2%, 3% y 4% 50, 75 y 100 |
20, 25 y 30 min | |
| Tratamientos | Agentes encapsulantes | Tiempo de polimerización (min) | |
| Alg-Na (%) | CaCl2 2H2O (mM) | ||
| T1-1 | 2 | 50 | 20 |
| T1-2 | 2 | 50 | 25 |
| T1-3 | 2 | 50 | 30 |
| T3-1 | 3 | 50 | 20 |
| T3-2 | 3 | 50 | 25 |
| T3-3 | 3 | 50 | 30 |
| T4-1 | 3 | 75 | 20 |
| T4-2 | 3 | 75 | 25 |
| T4-3 | 3 | 75 | 30 |
| T5-1 | 3 | 100 | 20 |
| T5-2 | 3 | 100 | 25 |
| T5-3 | 3 | 100 | 30 |
| T6-1 | 4 | 75 | 20 |
| T6-2 | 4 | 75 | 25 |
| T6-3 | 4 | 75 | 30 |
Proceso de encapsulación
En condiciones asépticas, las microestacas se suspendieron una por una en la solución de alginato de sodio. Con una micropipeta de 1000 µl cuya punta fue adaptada con una abertura de 7 mm, la microestaca fue revestida y aspirada con un volumen de 100 µl de alginato de sodio y se dejó caer a manera de gota en el matraz con la solución de cloruro de calcio dihidratado. Al entrar en contacto se inició la reacción de polimerización (Figs. 1A-F).

Figura 1: Elaboración de semillas sintéticas de Castilleja tenuiflora Benth.: A. plantas madre con 21 días de edad provenientes de cultivo in vitro; B. microestacas de C. tenuiflora; C. las microestacas se suspenden en la solución de alginato de sodio y luego se aspiran con una micropipeta; D. los explantes se vierten en forma de gota en la solución de cloruro de calcio; E. se decanta la solución de cloruro de calcio y las semillas sintéticas obtenidas se enjuagan 3 veces con agua desionizada estéril; F. obtención de una semilla sintética de C. tenuiflora.
Para dar forma esférica a la semilla sintética, cada matraz fue colocado en un agitador orbital (Prendo, modelo AGO 60-40, Puebla, México), a 100 rpm. Las soluciones se dejaron reaccionar en estas condiciones durante 20, 25 y 30 min y una vez transcurrida la polimerización, las semillas sintéticas fueron recuperadas al decantar el cloruro de calcio dihidratado. Con agua desionizada esterilizada se realizaron tres lavados y se dejaron secar por un minuto sobre papel filtro en condiciones asépticas (Ara et al., 2000; Sharma et al., 2013).
Cada semilla fue sembrada de forma individual en viales de 10 cm de largo y 1 cm de diámetro, con 1.5 ml de fitagel (1.5 g l-1) y se incubaron a 25±2 °C, intensidad luminosa de 1800 Lux, con un fotoperiodo de 16/8 horas luz/obscuridad.
Efecto de las concentraciones de los agentes encapsulantes y el tiempo de polimerización
Las semillas sintéticas obtenidas fueron sembradas sobre fitagel (sin ningún medio nutritivo o regulador de crecimiento) para evaluar viabilidad, germinación, conversión y longitud de la planta. Las observaciones se realizaron a los 9, 12, 15, 18 y 21 días después de la siembra. El criterio para definir la germinación de la semilla sintética se definió cuando el brote rompió la cápsula (Asmah et al., 2011). La viabilidad fue considerada cuando los explantes encapsulados presentaron una apariencia verde, sin amarillamiento y sin partes necrosadas (Micheli et al., 2007). El porcentaje de conversión se consideró como la formación de una planta completa proveniente de una semilla sintética (Standardi y Micheli, 2012). La longitud de las plantas después de la germinación fue medida con escala milimétrica, desde la superficie de la cápsula.
Después de seleccionar la concentración óptima de alginato de sodio, cloruro de calcio dihidratado y el tiempo de exposición, se dio continuidad a los experimentos de almacenamiento en frío de las semillas sintéticas.
Almacenamiento en frío
Con el fin de prolongar el tiempo de almacenamiento de las semillas sintéticas de C. tenuiflora se probaron tres temperaturas de almacenamiento (1.5, 4 y 25 °C) (Islam y Bari, 2012). A estas temperaturas, se evaluó la viabilidad (Fig. 2) de los explantes de las semillas sintéticas almacenadas a 20, 38 y 45 días. Con estos días de almacenamiento, se seleccionó la mejor temperatura y se certificó que los explantes contenidos en las semillas fueran viables.

Figura 2: Criterios de evaluación de viabilidad en la semilla sintética de Castilleja tenuiflora Benth. después del almacenamiento: A. explante viable, que muestra apariencia verde; B. explante con extremos necrosados; C. explante ámbar; D. explante blanco.
Después de seleccionar la temperatura de almacenamiento y viabilidad, se procedió a ampliar el periodo de almacenamiento con la mejor temperatura (4 °C), los tiempos de almacenamiento: 30, 50, 100, 190 y 200 días.
Las condiciones de almacenamiento fueron semillas en grupos de 30 dentro de criotubos de 5 ml y envueltos con papel aluminio, con el fin de mantenerlos en condiciones de oscuridad. Bajo esta condición se evita el deterioro de la matriz encapsulante (San Andres etal., 2010). Además, es posible retardar el crecimiento de los tejidos vegetales (Engelmann, 2011).
Los grupos de semillas, al final del periodo de almacenamiento, fueron sembradas en fitagel y se incubaron por 30 días. Las plantas resultantes se utilizaron para la elaboración de extractos metanólicos, y el análisis del perfil fitoquímico.
Perfil químico por LC-MS de plántulas de Castilleja tenuiflora para la determinación de los compuestos mayoritarios
Se elaboraron extractos metanólicos de las plantas de C. tenuiflora provenientes de semillas sintéticas y obtenidas de las pruebas de almacenamiento a 4 °C, con tiempo de almacenamiento de 30, 50 y 100 días. Estos fueron comparados con extractos metanólicos elaborados con plantas de C. tenuiflora provenientes de cultivo in vitro sin almacenar.
Para realizar el extracto, las muestras secas se colocaron en tubos Eppendorf de 2.5 ml y se molieron en el homogeneizador de tejidos de aire (QIAGEN, Tissue Lyser LT, Hilden, Alemania) durante 5 min a 100 rpm. Posteriormente, la muestra se suspendió en un criotubo de 5 ml con 1.5 ml de metanol (proporción de 1:50) y se sonicó durante una hora a temperatura ambiente. El extracto metanólico fue filtrado con papel filtro Wathman no. 1 y se dejó evaporar hasta sequedad. Los extractos fueron almacenados en la obscuridad hasta su análisis.
Para el análisis del perfil químico se utilizó un equipo de cromatografía líquida (Shimadzu, LC-MS 2020, Tokio, Japón), con un sistema de control CBM-20Aª, dos bombas LC-20AD, un desgasificador DGU-20A 5R, un muestreador automático SIL-20AC HT, un horno para columna CTO 20Aa y un detector de arreglo de fotodiodos (PDA) SPD-M20A, acoplado a un espectrómetro de masas con una fuente de ionización por electrospray (ESI-MS) (Shimadzu, Tokio, Japón).
El análisis se realizó a 40 °C en una columna de alta resolución RP-18 de fase reversa (Chromolith®, 100 mm por 4.6 mm, 5 µm) (Merck, Darmstadt, Alemania). La fase móvil fue previamente tamizada en filtro de membrana de 0.45 μm (Millipore, Milford, EUA) y desgasificada por sonicación durante 15 min. Como fase móvil se ocupó agua Milli-Q (solvente A) y acetonitrilo (solvente B). El método cromatográfico fue en gradiente de la siguiente forma: 0-2.0 min, 100% A - 0% B; 2.01-5.0 min, 90% A - 10% B; 5.01-10.0 min, 85% A - 15% B; 10.01-14.0 min, 80% A - 20% B; 14.01-18.0 min, 75% A - 25% B; 18.01-23 min, 70% A - 30% B; 23.01-27.0 min, 0% A - 100% B; 27.01-30.0 min, 100% A - 0% B.
La absorbancia fue medida a 205, 240, 280, 330, 360 y 380 nm. El volumen de inyección fue de 20 µl con un flujo constante de 1 ml/min-1. El análisis por espectrometría de masas se realizó en modo negativo en un intervalo de 100-1000 m/z (relación masa/carga de los iones que conforman el compuesto), bajo las siguientes condiciones: flujo del gas de nebulización (1.54 l min-1), flujo de N2 (10 l min-1), voltaje de la interfase (4.5 kV) y voltaje del detector (1.2 kV). Los compuestos químicos mayoritarios fueron identificados mediante su espectro UV-VIS de absorción (λ), el tiempo de retención (tr) y la relación masa atómica/carga del ion (m/z).
Análisis estadístico
Para seleccionar la concentración de agentes encapsulantes y tiempos de polimerización, la n fue de 10 semillas y se consideró una semilla sintética como unidad experimental. En la prueba de almacenamiento se probaron 30 semillas sintéticas por tratamiento y como control se utilizaron semillas sintéticas sin almacenamiento. El resultado de ambas pruebas fue analizado con la prueba de Tukey (α=0.05) con el software Infostat (v. 2018 (Di Rienzo et al., 2018).
Resultados
La mejor concentración de los agentes encapsulantes fue el alginato de sodio al 3% en complejo con el cloruro de calcio dihidratado al 50 mM con un tiempo de polimerización de 20-25 min. porque presentó el mayor porcentaje de viabilidad, conversión y las mejores longitudes. Además, la composición de las semillas sintéticas favoreció su manipulación y forma (Fig. 3).
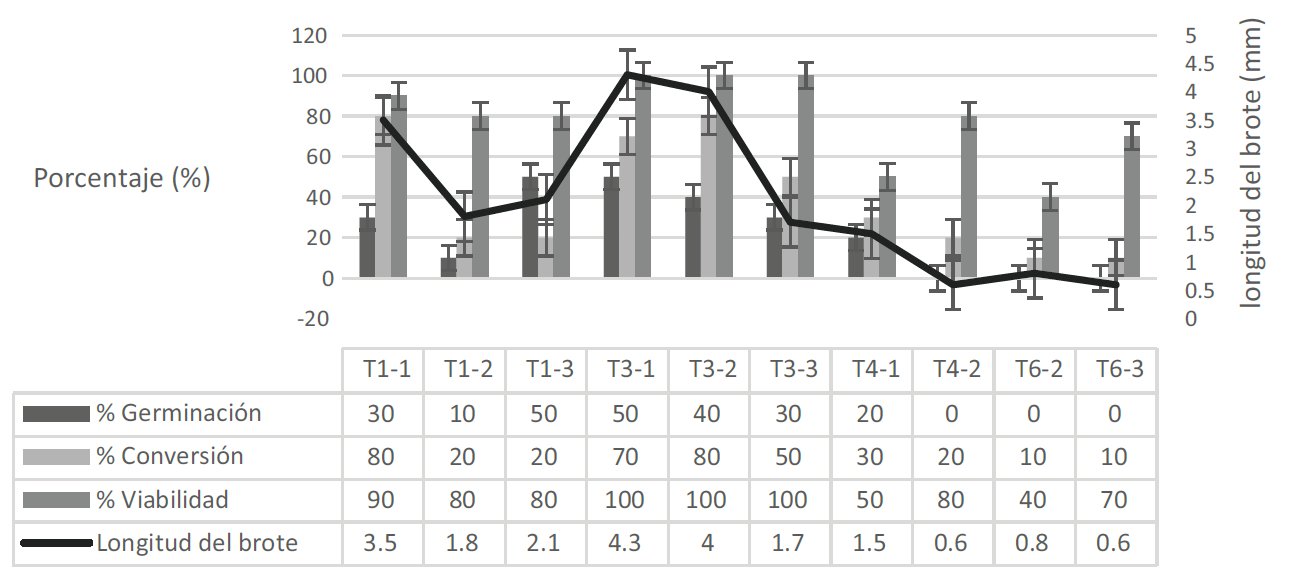
Figura 3: Efecto de las concentraciones de los agentes encapsulantes y tiempo de polimerización sobre el porcentaje de germinación (%), viabilidad (%), conversión (%) y longitud del brote (mm) a los 18 días de cultivo. Los tratamientos incluidos en el T3 registraron el mayor porcentaje de germinación de Castilleja tenuiflora Benth.
En los tratamientos de alginato de sodio 2%, cloruro de calcio dihidratado 50 mM y 30 min (T1-3), la germinación fue de 50%, la conversión de 20%, la viabilidad de 80%, los explantes permanecieron verdes aunque no presentaron desarrollo; de 20% de las muestras que presentaron conversión, la longitud del brote fue de 2.1 mm.
El tratamiento de alginato de sodio al 3%, cloruro de calcio dihidratado 50 mM y 20 min (T3-1) presentó una germinación de 50%, una conversión de 70% y mostró una viabilidad de 100%, la longitud del brote fue de 4.3 mm. En la viabilidad en el resto de tratamientos de T3 (T3-2 y el T3-3) no hubo diferencias significativas entre ellos, los explantes permanecieron verdes aunque no presentaron desarrollo; la viabilidad no tuvo influencia sobre la germinación. Las semillas elaboradas con las mayores concentraciones de alginato de sodio (4%) en combinación con el cloruro de calcio dihidratado (75 mM) registraron porcentajes de germinación menores a 30%.
Aunque el tratamiento T1-1 obtuvo alto porcentaje de conversión y viabilidad (80 y 90% respectivamente), las características físicas de las semillas no favorecieron la forma y la manipulación (Fig. 4); mientras que en la forma de los tratamientos con mayor concentración (T4 y T6) las cápsulas fueron esféricas, de color opaco y el explante no fue visible. El tratamiento T3-2 fue el que se seleccionó para el resto de los experimentos por presentar el mayor porcentaje de conversión y las cápsulas fueron esféricas y transparentes.

Figura 4: Efecto de las concentraciones sobre las características visuales de la semilla sintética de Castilleja tenuiflora Benth.: A. 2%, 50 mM; B. 3%, 50 mM; C. 4%, 75 mM.
En la prueba del efecto del almacenamiento en frío sobre la viabilidad del explante, los resultados muestran que el almacenamiento a 4 °C es la temperatura más efectiva debido a que las semillas sintéticas mostraron una apariencia viable en todos los periodos evaluados (Cuadro 2). En el almacenamiento a 1.5 °C, los explantes se mantuvieron viables hasta los 28 días; además, la cápsula presentó signos de deshidratación del polímero debido al almacenamiento en frío. Bajo la temperatura de almacenamiento a 25 °C no se observó la presencia de explantes viables en ningún periodo de almacenamiento. Debido a estos resultados se seleccionó como condición de almacenamiento la temperatura de 4 °C.
Cuadro 2: Efecto de la temperatura de almacenamiento sobre la viabilidad de semillas sintéticas de Castilleja tenuiflora Benth.
| Tiempo de almacenamiento (días) | Temperatura de almacenamiento (°C) | ||
| 1.5 | 4 | 25 | |
| 21 | viable | viable | café |
| 28 | viable | viable | café |
| 38 | café | viable | café |
| 45 | café | viable | café |
El tratamiento con 50 días de almacenamiento presentó los valores más altos y significativamente diferentes entre todos los tratamientos (Fig. 5). En este tratamiento se obtuvo una viabilidad de 93 y 77% de germinación, una longitud de planta promedio de 7.07 mm y un porcentaje de conversión de 70%.

Figura 5: Efecto del tiempo de almacenamiento a 4 °C sobre el porcentaje de germinación (%), viabilidad (%), conversión (%) y longitud del brote (mm) a los 18 días de cultivo in vitro de Castilleja tenuiflora Benth.
En los tiempos de almacenamiento 180 y 200 días ocurre una disminución significativa de las tres variables evaluadas, no hubo diferencias significativas entre ellos. La frecuencia de conversión a plantas para los diferentes tratamientos fue de la siguiente manera: almacenamiento de 50 días presentó 70 plantas; 100 días, 31 plantas y 30 días, 20 plantas (en las semillas almacenadas a 180 y 200 días no hubo conversión completa).
Perfil químico de compuestos mayoritarios en plantas de Castilleja tenuiflora obtenidas de semillas sintéticas después de un periodo de almacenamiento
Se identificaron los compuestos mayoritarios en muestras obtenidas de extractos metanólicos en plantas control y en las provenientes de semilla sintética con tiempos de almacenamiento: 30, 50 y 100 días (Cuadro 3). El grupo control presentó iridoides (aucubina y bartsiósido), feniletanoides (verbascósido e isoverbascósido) y flavonoides (rutina y luteolina). Por otro lado, el iridoide aucubina, el feniletanoide verbascósido y el lignano se encontraron presentes en todas las plántulas provenientes de semilla sintética, mientras que la presencia de otros compuestos fue variable.
Cuadro 3: Compuestos químicos identificados en plantas de Castilleja tenuiflora Benth., provenientes de encapsulados almacenados a diferentes tiempos. tR: tiempo de retención; Ion (M-H): ion molecular menos un hidrógeno cargado negativamente; (m/z): ion molecular, masa cargada del compuesto; C: plantas control.
| No. | tR (min) | Ion (M-H)-(m/z) | Compuesto | Grupo químico | Tratamientos |
| 1 | 4.36 | 345 | Aucubina | Iridoide | C, 30 d, 50 d, 100 d |
| 2 | 5.69 | 329 | Bartsiósido | Iridoide | C |
| 3 | 7.31 | 389 | 8-epiloganina | Iridoide | 50 d, 100 d |
| 4 | 9.36 | 609 | Rutina | Flavonoide | C, 30 d |
| 5 | 11.23 | 623 | Isoverbascósido | Feniletanoide | C, 30 d, 50 d, 100 d |
| 6 | 12.21 | 623 | Verbascósido | Feniletanoide | C, 30 d, 50 d, 100 d |
| 7 | 20.25 | 299 | Flavonoide | Luteolina | C |
La elaboración de semillas sintéticas y el almacenamiento hasta por 100 días no influyeron en el perfil químico de los compuestos mayoritarios identificados en las plantas donantes de C. tenuiflora.
Discusión
La forma y la consistencia de las cápsulas fueron dependientes de la concentración y tiempo de polimerización de la matriz; los porcentajes de germinación, conversión y viabilidad, así como la longitud de los brotes fueron diferentes de acuerdo a los tratamientos. En especies como Centella asiatica (Prasad et al., 2014), la concentración del alginato de sodio fue del 4% y del cloruro de calcio a 75 mM y se obtuvo 85% de germinación. Cabe mencionar que en su medio de conversión agregaron medio de cultivo MS, y para C. tenuiflora (alginato de sodio 3%, 50 mM de cloruro de calcio dihidratado) solamente se utilizó el agente gelificante (sin ningún medio nutritivo o regulador de crecimiento). Dichas condiciones fueron suficientes para obtener 80% de conversión. La eficiencia de la semilla sintética depende de la matriz de encapsulación y del medio de conversión, estos factores influyen sobre la viabilidad hasta la conversión a planta. Con esto se observó directamente el efecto de la concentración de los agentes encapsulantes sobre las variables evaluadas.
En este estudio el diseño de la composición de la matriz encapsulante incluye un medio nutritivo que se basó en el medio utilizado para micropropagar regularmente a C. tenuiflora en el laboratorio. El medio de conversión probado fue utilizando fitagel sin ningún medio nutritivo o regulador de crecimiento; estas condiciones fueron suficientes para la obtención de plantas. Sin embargo, si se desea alcanzar mayor tamaño en las plantas, es necesario optimizar tanto el medio nutritivo que incluye la matriz de encapsulación, como el medio de conversión.
Tsvetkov et al. (2006) estudiaron el efecto de la composición de la matriz encapsulante sobre el rebrote in vitro de microcortes encapsulados del híbrido Populus tremula L. × P. tremuloides Mincx., determinando que la adición de nutrientes, una fuente de carbono (sacarosa) y reguladores de crecimiento vegetal, dan condiciones que favorecen el desarrollo inicial del explante encapsulado. Así también lo mencionan Sundararaj et al. (2010) al aumentar la concentración de elementos nutritivos (MS-solución base) en la matriz de encapsulación, se favorece la formación de brotes en microcortes encapsulados de Zingiber officinale Rosc. Estos resultados son similares al estudio realizado por Saiprasad (2001) aplicado en embriones somáticos.
Sin embargo, los requerimientos para la disponibilidad de ciertos ingredientes de la matriz de hidrogel son especie específico, por lo que se debe de dar atención especial en los requerimientos de la especie de interés (Tsvetkov et al., 2006). Por ejemplo, estos autores mencionan que en particular la fuente de carbono, presente en forma de sacarosa, representa un elemento clave para dar inicio al evento de re-crecimiento en explantes de Populus, y se comprobó que este elemento tiene mayor influencia sobre la longitud del brote comparado con 6-bencilaminopurina (BAP). Cabe mencionar el estudio realizado en Beta vulgaris L. (Rizkalla et al., 2012) donde se evaluó el efecto de la concentración de la fuente de carbono en la matriz encapsulante sobre el porcentaje de germinación. Los mejores resultados de germinación para la especie se obtuvieron al adicionar medio MS con sacarosa (30 g/l) a la matriz de encapsulación y al aumentar esta concentración se afectó el porcentaje de conversión negativamente.
Es posible que en el presente estudio la aplicación conjunta de ácido indol butírico (IBA), BAP y sacarosa, favorecieran la formación de brotes en las microestacas encapsuladas ya que estimulan la progresión del ciclo celular y son las hormonas claves para inducir la formación de novo de brotes en diversos explantes in vitro (hojas, raíces, medula, cotiledones) (Jordán y Casaretto, 2006).
En este estudio, el medio de conversión no fue enriquecido con nutrientes ni reguladores de crecimiento para observar de forma directa el potencial de la semilla sintética para obtener plantas debido a que la conversión es el aspecto más importante de esta tecnología para su uso comercial (Standardi y Piccioni, 1997).
Standardi y Piccioni (1997) dicen que el medio de conversión puede optimizarse con reguladores de crecimiento vegetal (RCV) o aumentando otras fuentes como la de carbono y elementos nutritivos (Sundararaj et al., 2010), así como antibióticos, herbicidas, biocontroladores y biofertilizantes, entre otros (Islam y Bari, 2012).
Las concentraciones seleccionadas en este estudio favorecieron la viabilidad, germinación, conversión de la planta de C. tenuiflora y su forma; por lo tanto, se facilitó su manipulación, caso contrario con las concentraciones menores. Esto también se observa en plantas medicinales, como lo muestran los resultados expuestos por Lata et al. (2009) en Cannabis sativa L., donde el tamaño promedio de sus cápsulas fue de 5 mm, semejantes a este estudio.
Gantait et al. (2012) reportan que el uso de diferentes concentraciones de alginato de sodio presenta diferencia en la morfología de las cápsulas (opacidad y forma), concluyendo que las concentraciones de alginato de sodio pueden variar según la fuente comercial y la especie con la que se trabaje. Lo más importante es producir cápsulas que faciliten la manipulación durante el cultivo in vitro, siembra o almacenamiento. Elaborar semillas esféricas, uniformes y firmes son indicativos cualitativos que pueden funcionar como indicador para tal propósito.
Standardi y Piccioni (1997) explican que la característica más deseable de un microcorte encapsulado es su capacidad de mantener la viabilidad en términos de rebrote y crecimiento incluso después de un periodo de almacenamiento. En este estudio fue posible obtener valores mayores a 70% en los parámetros de viabilidad, germinación y conversión en el tiempo de almacenamiento de 50 días a 4 °C para semillas sintéticas de C. tenuiflora. Al respecto, es importante mencionar que en las zonas nodales de una planta se encuentran los meristemos, células morfológicamente indiferenciadas que contienen toda la información genética para dividirse de manera ordenada proporcionando a las plantas una mayor actividad celular y dando como resultado la formación de nuevos brotes. Las zonas nodales podrían requerir mayor cantidad de nitrógeno, ya que este elemento es esencial para que las plantas sinteticen ácidos nucleicos y aminoácidos, entre otros compuestos. Por otro lado, Tacoronte et al. (2017) reportan que el contenido de nitrógeno favorece el crecimiento de explantes de papa en condiciones in vitro en medio de cultivo MS (Murashige y Skoog, 1962), el cual se caracteriza por su alto contenido de nitratos y amonio. El valor de las variables disminuye cuando el tiempo de almacenamiento aumenta (100, 180 y 200 días). Este mismo efecto se observa en el estudio realizado por Sundararaj et al. (2010).
Finalmente, se observó que el grupo control y las plantas derivadas de semillas sintéticas comparten los principales compuestos de interés. Tal es el caso de compuestos del tipo iridoide (aucubina) y el feniletanoide (verbascósido), mientras que la presencia de otros compuestos fue variable, ya que la producción de metabolitos secundarios es consecuencia de la respuesta al estrés y a las variaciones ambientales (Chaves et al., 2001). Otro ejemplo es el contenido de D9-tetrahidrocannabinol, de las plantas de Cannabis sativa provenientes de explantes encapsulados, que fue similar en relación a las plantas donantes después de 168 días (24 semanas) de almacenamiento a 15 °C (Lata et al., 2012); por lo que la tecnología de semillas sintéticas es una buena alternativa como método para la conservación de germoplasma. Es necesario realizar estudios detallados sobre la caracterización bioquímica de las plantas derivadas de semillas sintéticas para poder explicar el comportamiento de los metabolitos secundarios durante el proceso de encapsulación de microestacas (Faisal et al., 2013).
Conclusiones
En este estudio se demostró que es posible obtener plantas de C. tenuiflora a partir de semillas sintéticas de microestacas elaboradas con los agentes encapsulantes de alginato de sodio en complejo con el cloruro de calcio dihidratado. Este también fue un método efectivo para su propagación y almacenamiento in vitro.
Las condiciones de encapsulación de microestacas de C. tenuiflora que se proponen, permitió el almacenamiento del material vegetal manteniendo los principales compuestos mayoritarios identificados en esta planta. Se considera entonces que el método para la encapsulación y conservación de microestacas es una alternativa para mantener el genotipo de plantas élite, facilitar el intercambio de germoplasma, contar con materia prima disponible para el cultivo ex vitro, minimizar los riesgos de contaminación y contrarrestar el saqueo de plantas de su hábitat natural.

