











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
The genus Hydrangea L. (Hydrangeaceae) consists of approximately 200 taxa, distributed in Asia and America, classified in 15 sections (De Smet et al., 2015). The relatively poorly known Hydrangea section Cornidia (Ruiz & Pav.) Engl. consists of 13 currently accepted taxa (12 species and one variety) and a yet undefined number of species new to science. The section is distributed from northern Mexico to southern Chile and Argentina with one species, Hydrangea integrifolia Hayata, in China, Taiwan and the Philippines (Samain et al., 2014; Samain and Martínez Salas, 2015). These are evergreen root climbers growing up to 60 m high in the canopy of mostly primary forests, or rarely on boulders and rock walls (Granados Mendoza et al., 2014), with coriaceous leaves and hortensia-like whitish, greenish, yellowish or purplish inflorescences, with or without enlarged marginal flowers.
According to several recent molecular studies, Hydrangea section Cornidia (hereafter shortened as Cornidia) is monophyletic, including the single Asian species (Samain et al., 2010; Granados Mendoza et al., 2013a, b, 2015; De Smet et al., 2015). Cornidia is sister to the section Calyptranthe Maxim., consisting of Asian climbing species (De Smet et al., 2015). Both clades together are sister to the section Asperae (Rehder) Y. De Smet & Samain, encompassing Asian shrubby species (De Smet et al., 2015). We note here that we do not follow the nomenclatural changes proposed by Ohba and Akiyama (2016), who propose to recognize Cornidia again as a segregate genus within tribe Hydrangeeae, together with most of the other sections published by De Smet et al. (2015), as they do not take into account the evolutionary context in which the new classification of the tribe Hydrangeeae was presented.
The monography of the genus Hydrangea s.s. by McClintock (1957), which is entirely based on herbarium specimens, is the only available recent revision of this section and includes 12 accepted species, classified in two subsections, Monosegia Briq. and Polysegia Briq. As mentioned by Samain et al. (2014), the revision by McClintock (1957) oversimplifies the high morphological variation within Cornidia in the Neotropics. This is not only due to inappropriate synonymization of taxa (since the description of Cornidia as a genus by Ruiz and Pavón (1794), more than 40 taxa have been described within this group), but especially to the fact that many diagnostic morphological characters such as leaf and floral traits were not considered, making the identification key only practicable with some difficulty (Samain et al., 2010). For instance, distinction is made between the two Cornidia subsections based on their inflorescence architecture: a single terminal cluster in Monosegia and several clusters in Polysegia (McClintock, 1957). However, several specimens of different species we have observed both in the field and in herbarium collections show both inflorescence types, indicating that these subsections are not natural, which is also reflected by our ongoing molecular work (unpublished data). Moreover, several herbarium specimens included in the revision of McClintock (1957) in a specific taxon bear apart from that first annotation on the respective specimen also a later identification by the same author with another taxon name. However, any reasons for these changes in identification or in taxon circumscription and concept are not given.
An additional challenge in Cornidia is the functional dioecism, as mentioned already by Nevling and Gómez-Pompa (1968), Freire-Fierro (2004), Samain et al. (2014), as well as Samain and Martínez Salas (2015). This sexual dimorphism, which we have observed in most individuals during our extensive field work throughout the Neotropics, is reflected by a notoriously different size and shape of flower receptacle anthers and pistils between functionally female and male flowers. Additional morphological differences between female and male plants such as noted for the leaf shape of Hydrangea albostellata Samain, Najarro & E. Martínez (Samain et al., 2014) and the robustness of the inflorescence of several of the new species published here may result in some difficulty to match female and male individuals of a particular species. Interestingly, during our ongoing revision of this group, we have noticed that some species do not show this functional dioecism at all (e.g., H. seemannii L. Riley which has bisexual flowers, see this treatment), whereas some others may show functionally dioecious or perfect flowers depending on the individual.
The abovementioned issues emphasize the need for a complete and urgent revision of Cornidia, especially in the light of conservation of these species. Indeed, their pristine habitat with very specific conditions (near water, flat topography, efficient drainage) makes them not only promising bio-indicators for this kind of habitat, but also highly endangered as these habitats become rarer because they are highly appreciated by local people for agriculture. The present work aims at a revision of the genus Hydrangea in Mexico, based on 186 collections, 179 from Mexico and seven from Guatemala. Ninety-three of these were realized in the framework of the current project, all of which we observed thoroughly in living conditions in their natural habitat, before preparing herbarium specimens that were used to elaborate detailed morphological descriptions.
Mexico is the fourth most diverse country in the world in terms of vascular plant diversity (e.g., Villaseñor, 2016), yet many new plant species still awaiting discovery are already threatened with extinction as a consequence of deforestation and habitat destruction (e.g., Calderón-Aguilera et al., 2012). Mexico´s impressive biodiversity is mainly due to its extremely heterogeneous physical-geographical context with 11 morphotectonic provinces with distinct physiographic and geologic-tectonic features, three geological plates, two biogeographic realms and nearly all globally known vegetation and climate types (e.g., Miranda y Hernández X., 1963; Ramamoorthy et al., 1993; García, 2004).
The genus Hydrangea exemplifies the lack of taxon-specific botanical collection activities in Mexico, especially in areas which are logistically difficult to reach, or which have the reputation of being unsafe to develop such activities. Of the 93 previous collections made before we started our survey in 2009, 48 are from the temperate species H. seemannii which is restricted to northwestern Mexico, and 45 are from the rest of the country, mainly H. albostellata (14 collections) and H. nebulicola Nevling & Gómez Pompa (12). Based on preliminary niche modeling data (O. Téllez Valdés, unpublished results), we especially explored in the state of Veracruz on the mountain slopes towards the Gulf of Mexico and in the state of Chiapas, where we collected in total 71 specimens, more than doubling the number of known collections in this area. The multiplication of the total number of Hydrangea collections in tropical southeast Mexico since we have started our survey is even more surprising taking into account the considerable size of these plants, which may measure more than 40 m.
McClintock (1957) mentions for Mexico only H. seemannii for the state of Durango in the northwest. This species is now known to occur also in the states of Chihuahua, Nayarit, Sinaloa and Sonora. In several localities in Durango where we observed this species, it is highly endangered and ecological niche modeling as well as observations in its habitat show that it may be close to extinction in the wild in about 50 years (Granados Mendoza et al., 2010). Floristic treatments of the family for the Flora de Veracruz (Durán-Espinosa, 1999) and the Flora del Bajío (Pérez-Calix, 2004), respectively, report H. nebulicola for the states of Querétaro and Veracruz.
Hydrangea steyermarkii Standl. is a species described from Guatemala in the border area with Mexico (Standley, 1940) and the few Hydrangea collections of that country as well as of the state of Chiapas have been annotated as such, irrespective of their highly different morphological traits, ecological preferences and geological affinities of the vegetation type in which they occur. According to our current insights (see also Samain et al., 2014), H. steyermarkii is restricted to the Central American tectonic plate, whereas all other Mexican species grow on the North American tectonic plate, with the exception of H. sousae Samain, Najarro & E. Martínez sp. nov. which occurs exactly on the border of both.
Based on meticulous observations in the field and of herbarium specimens, we describe here six new climbing Hydrangea species of Mexico, bringing the total amount of known species for this country to ten. We present a key to all native Mexican Hydrangea species and one widely cultivated and naturalized species, as well as two amended descriptions of earlier described species, illustrations, and distribution maps, and discuss their diversity, distribution and general conservation status within the country, as well as their affinities with the other species of section Cornidia.
Material and Methods
Field work has been carried out in the Mexican states of Chiapas, Durango, Hidalgo, Oaxaca, Puebla, Querétaro and Veracruz, during dry and rainy seasons of 2009 to 2017, coinciding with the flowering seasons of all species included. Initially, exploring field work was carried out in the areas where previous collections had been made. Consequently, when we realized that Hydrangea is much more common than initially assumed, we explored in many forest patches with primary, humid habitats of which we observed they are favoured by Hydrangea, especially in the state of Chiapas. Branches with inflorescences, flowers and fruits of all stages were collected and preserved. Moreover, in several individuals where architectural traits seemed to be important, whole branches were collected, cut and subsequently numbered in order to maintain the architecture available for further study. All specimens were deposited in the National Herbarium of Mexico (MEXU), in the herbarium of the Instituto de Ecología, A.C. in Pátzcuaro, Michoacán, Mexico (IEB), and in the herbarium of Ghent University, Belgium (GENT), whereas most of them are also deposited in the herbarium of the Instituto de Historia Natural in Tuxtla Gutiérrez, Chiapas (CHIP). Holotypes of each new species were deposited in MEXU, and, apart from the abovementioned herbaria, isotypes were also sent to other herbaria in Mexico, the United States of America and Europe.
The Red List categories of the ten native Mexican Hydrangea species were obtained according to the IUCN Red List criteria (IUCN, 2012). All known localities, including those of herbarium specimens without coordinates, were georeferenced using Google Earth (2018). Extent of Occurence (EOO) and Area of Occupancy (AOO) of all species were calculated with GeoCat (Bachmann et al., 2011). Distribution maps were realized with the same set of coordinates using ArcMap 10.1 (ESRI, 2010).
Results
Taxonomy
The present treatment covers, apart from the morphological description of the section to which the species belong, a full description of eight species. The descriptions and additional information of the two species treated by Samain et al. (2014) are not repeated here as the original article has been published as an Open Access paper.
Hydrangea section Cornidia (Ruiz & Pav.) Engl., Nat. Pflanzenfam. 3(2a): 76. 1891. TYPE: Cornidia umbellata Ruiz & Pav. (currently accepted name Hydrangea preslii Briq.).
≡ Cornidia Ruiz & Pav., Fl. Peruv. Prodr. 53: pl. 35. 1794.
Root-climbing lianas up to 40 m (exceptionally 60 m) high, generally growing on trees (or very rarely on rocks and vertical rock walls), remaining below the canopy, or protruding above the canopy with an apical shrub-like portion, often with juvenile reddish branches near the base of the individual creeping on the host stem or on the soil towards other host trees; adult stem at the base up to 20 cm diameter, consistently maintaining this width until the flowering apex, or gradually tapering towards the apex; twigs glabrous, or with different indument types, flexible or brittle depending on the species; leaves decussate, coriaceous, glabrous or with different indument types, venation variable and species-specific, adaxial lamina side often with acarodomatia in the axils between the first and second order veins, exceptionally such acarodomatia between second and third order veins, margin entire, dentate, serrate, lobed or undulated, sometimes with glands; inflorescences lateral, clusters umbel-like, small to large, branching loosely to densely, in bud enveloped by whitish, yellowish or greenish broadly ovate involucral bracts, shedding quickly at the start of anthesis, leaving noticeable scars, inflorescence stalks with or without small leaves, shedding during or after anthesis, leaving noticeable scars, often rhytmic zones of scars; flowers whitish, yellowish, or greenish, rarely pinkish, rarely with enlarged marginal flowers (these in the Mexican species only in H. seemannii consistently present; enlarged marginal flowers much more common in Central and South American species), hypanthium relatively flat to cup-shaped, broad to narrow, sepals reduced to small lobes on the hypanthium, 4-5, petals flag-shaped, rounded, 4-5, shedding quickly after anthesis, stamens 8-9, shorter than pistils and with shriveled anthers in female plants, longer than pistils in male plants, shedding quickly after anthesis, not leaving any visible scar after shedding, pollen tricolpate, pistils 2, rarely 3 (the presence of 3 or more pistils is more common in some Central and South American species), longer than stamens in female plants, shorter than stamens and without stigmatic surface in male plants; fruits a capsule, dehiscing apically between the dried pistils, brownish, shape similar to hypanthium shape, size generally similar to hypanthium size, sometimes enlarging (e.g., in H. albostellata); seeds numerous, very small.
Key to the species of Hydrangea section Cornidia in Mexico
The flower corolla of all Mexican species is white, sometimes slightly greenish or pinkish, or with reddish lines, but never red or purple as in several Central and South American species. Only one species, H. seemannii, consistently shows enlarged marginal flowers, whereas several other species occasionally or very rarely possess these flowers (H. albostellata, H. sousae and H. nebulicola). This key also includes H. macrophylla (Thunb.) Ser., a hortensia native of Japan, which is widely cultivated at global level and naturalizing in Mexico and other countries in the Neotropics.
1a. Shrubby plants of 1-2 m high, with inflorescences consisting entirely of blueish, whitish or pinkish enlarged flowers, naturalised ............................................................ H. macrophylla (Thunb.) Ser.
1b. Root-climbing plants measuring up to 60 m high, with whitish, yellowish or greenish inflorescences without or with enlarged flowers ..................................................... 2
2a. Enlarged marginal flowers always present, numerous and large, plants from northwestern Mexico...................................................................................................... H. seemannii L. Riley
2b. Enlarged marginal flowers generally absent, if present few and small, irregularly placed within the inflorescence, plants from central and southeastern Mexico ............. 3
3a. Leaves, inflorescences and branch apices covered by a very dense indumen ......... 4
3b. Leaves, inflorescences and branch apices glabrous to slightly pubescent ............... 5
4a. Pubescence dense reddish, lamina spoon-shaped; inflorescence 4-5 × 3-4 cm ........................................................................................................................................ H. nebulicola Nevling & Gómez Pompa
4b. Pubescence dense whitish to yellowish, lamina flat, inflorescences. 4-5 × 2.5-3 cm ..................................................................................................................................... H. otontepecensis Samain & E. Martínez
5a. Lamina spoon-shaped, acarodomatia on abaxial leaf side absent .............................................................................................................................................. H. albostellata Samain, Najarro & E. Martínez
5b. Lamina flat, acarodomatia on abaxial leaf side present ................................................ 6
6a. Leaf margin lobed, undulated, crenate or dentate ......................................................... 7
6b. Leaf margin entire ...................................................................................................... 9
7a. Leaf margin slightly undulated to lobed or dentate with glands, marginal vein present ...................................................................................................................................... H. tapalapensis Samain, Najarro & E. Martínez
7b. Leaf margin slightly crenate ........................................................................................ 8
8a. Apical shrub-like growth above its host tree canopy, internodes very reduced, with compacted leaves and inflorescences, leaves elliptic to lanceolate, leaf margin revolute with marginal vein ...................................................................................................................................... H. sousae Samain, Najarro & E. Martínez
8b. Plant never growing above its host tree canopy, internodes very elongated, without compacted leaves and inflorescences, leaves lanceolate-obovate, leaf margin not revolute .......... H. steyermarkii Standl.
9a. Leaves abaxially relatively densely pubescent with adpressed yellowish-brownish-reddish stellate hairs .................................................................................................................. H. nahaensis Samain & E. Martínez
9b. Leaves abaxially glabrous ......................................................................................... 10
10a. Leaves of the inflorescence axes narrowly lanceolate, pistils of the female flowers 1.3-1.5 mm long .............................................................................................. H. breedlovei Samain, Najarro & E. Martínez
10b. Leaves of the inflorescence axes ovate, pistils of the female flowers 1.5-2.5 mm long ...................................................................................................................... H. carroniae Samain & E. Martínez
Hydrangea albostellata Samain, Najarro & E. Martínez, Phytotaxa 162: 183. 2014. Fig. 1.
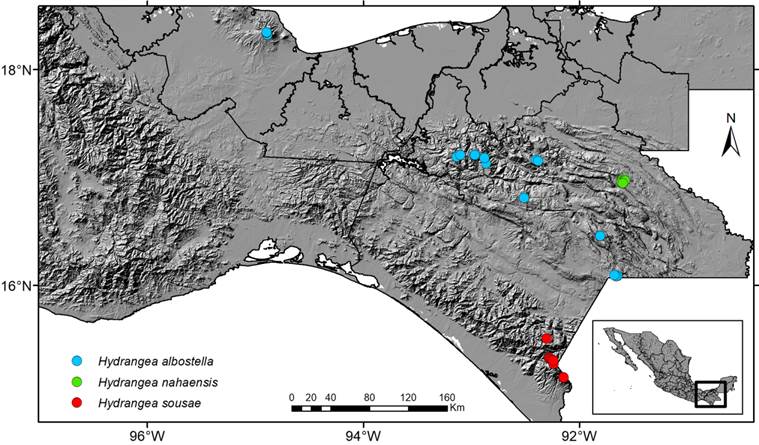
Figure 1: Distribution of Hydrangea albostellata Samain, Najarro & E. Martínez, H. nahaensis Samain & E. Martínez and H. sousae Samain, Najarro & E. Martínez in Mexico. Map by Rosario Redonda Martínez.
See Samain et al. (2014) for a complete description and notes on habitat, distribution, phenology, additional observations as well as a list of all then known collections. Additionally, among herbarium specimens of the region of Los Tuxtlas, Veracruz, we encountered several specimens which had been identified as H. nebulicola, but which also belong to this species. Only its distribution in Mexico is shown in Figure 1.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), Hydrangea albostellata is Endangered, with an Area of Occupancy (AOO) of 184 km2, despite its Extent of Occurence (EOO) being nearly 260000 km2, its populations allover its distribution area being severely fragmented and quickly being reduced because of logging.
Additional specimens examined: MEXICO. Veracruz, municipality Soteapan, 2 km antes de Santa Martha, Sierra de Santa Martha, 1250 m, 27.XII.1978, J. I. Calzada 5129 (F, MEXU, XAL) (functionally female); ejido de Santa Martha, Sierra de Santa Martha, 1220 m, 22.XII.1978, J. I. Calzada 5138 (XAL) (functionally female); ejido de Santa Martha, 1250 m, 11.IX.1980, F. Vázquez and D. Hernández 2 (F, IEB, MEXU, XAL) (functionally male).
Hydrangea breedlovei Samain, Najarro & E. Martínez, sp. nov. Figs. 2, 3, 4.
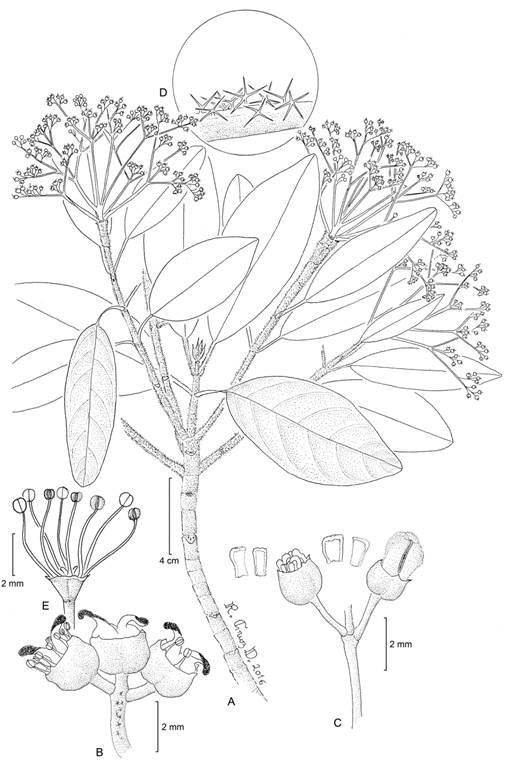
Figure 2: Hydrangea breedlovei Samain, Najarro & E. Martínez: A. branch with inflorescences; B. functionally female flowers; C. portion of the inflorescence, petals were removed from the left flowers; D. indument on the adaxial portion of the inflorescence; E. functionally male flowers. A, C, D, M. S. Samain and E. M. Martínez Salas 2012-002 (holotype: MEXU); B, M. S. Samain and E. M. Martínez Salas 2012-001 (MEXU); E, M. S. Samain et al. 2012-004 (MEXU). Drawing by Ramiro Cruz Durán.

Figure 3: A-C, Hydrangea breedlovei Samain, Najarro & E. Martínez: A. habit; B. inflorescence of male individual; C. inflorescence of female individual; D-F, H. carroniae Samain & E. Martínez: D. inflorescence of female individual, E. apical portion of flowering branch, with kataphylls; F. young inflorescence with inflorescence bract and flower buds. Photos by Marie-Stéphanie Samain and Esteban Manuel Martínez Salas.
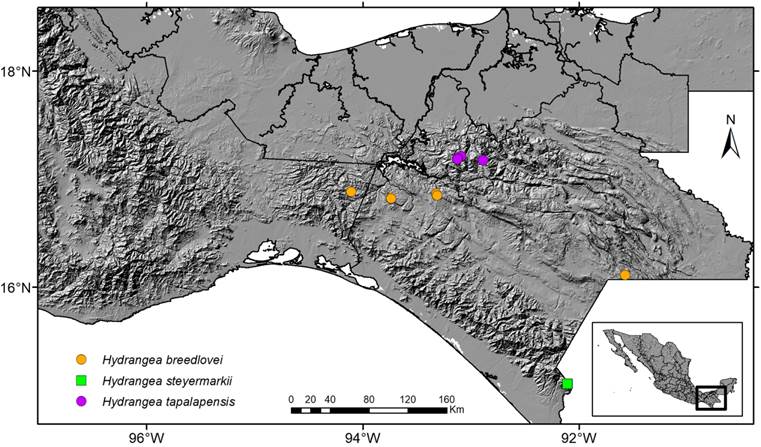
Figure 4: Distribution of Hydrangea breedlovei Samain, Najarro & E. Martínez, H. steyermarkii Standl. and H. tapalapensis Samain, Najarro & E. Martínez in Mexico. Map by Rosario Redonda Martínez.
TYPE: MEXICO. Chiapas, municipality Berriozábal, Vistahermosa, 1195 m, 16°50'57.01"N, 93°18'40.55"W, 31.XII.2011, M. S. Samain and E. M. Martínez Salas 2012-002 (holotype: MEXU!, isotypes: BIGU!, CAS!, CHIP!, CIIDIR!, CR!, DUKE!, F!, GENT!, GH!, iBUG!, IEB!, K!, MEXU(3)!, MICH!, MO!, NY!, US!, XAL!) (functionally female).
Functionally dioecious species endemic to southern Mexico which can be distinguished from Hydrangea steyermarkii by the narrowly lanceolate to elliptic leaves, entire leaf margin, glabrous mature leaves, yellow-brown abaxial leaf side in dried specimens, and 3-4 decussate leaf pairs below the inflorescence.
Root-climbing liana of up to 25 m high, functionally dioecious; main stem clinging to the host tree with many adventitious roots, 10-12 cm diameter, bark grey brown to reddish brown, fissured and exfoliant, with few lenticels; runner shoots (stolons) reddish, with reddish adventitious roots and stellate hairs, with small kataphylls; branches many-ribbed, slightly angular, with many kataphyll and leaf scars, branches attached to host tree with adventitious roots, these roots fibrous, slightly flattened, with minute reddish deciduous indument, free-growing branches many-ribbed, slightly angular to strongly quadrangular, slightly fissured and exfoliant; leaves decussate, petiole sulcate adaxially, terete with well-defined lateral ribs, margin of ribs sometimes slightly corchy, colour dark red to brown, when young with adnate whitish stellate hairs, when mature glabrous, 1-2.5 cm long, 2-4 mm broad, broadened at its basis, leaving a concave semicircular to triangular scar on the branch when leaves fall; lamina flat, narrowly lanceolate to elliptic, 5-15 cm long, 3-6.5 cm broad, base cuneate, apex slightly acuminate, leaf margin entire, venation brochidodromous, veins 6-9 pairs, adaxial leaf side with well-defined reddish-brown midvein, secondary veins very lightly marked, greenish, tertiary veins not visible, glabrous, opaque green, abaxially with protruding midvein, secondary veins slightly protruding, tertiary veins slightly marked, glabrous in mature leaves, in younger leaves with deciduous pubescence consisting of adnate white stellate hairs, yellow-brown, young leaves abaxially with adnate whitish stellate hairs, adaxially with slightly more dense reddish stellate hairs, acarodomatia consisting of a well-defined cavity without hairs; inflorescence axis nearly glabrous with a few reddish simple hairs, 5-15 cm long, robust in functionally female plants, delicate in functionally male plants, with 3-4 decussate leaf pairs below the inflorescence, petiole 1-2.5 cm long, 1-3 mm broad, lamina narrowly lanceolate, 7-10 cm long, 2-4 cm broad, sometimes lower or apical leaf pair with reduced lamina, apex of the floral axis woody, cone-shaped, elongated bract scars visible, thickening at the top, quadrangular, 4 mm broad, 3-4 mm high in functionally female plants, slightly tapering towards the apex, 3 mm broad, 3-5 mm high in functionally male plants, inflorescence bracts not seen, inflorescences lateral, opposite, 2 opposite to 3-4, sometimes 6 decussate inflorescences per flowering branch which continues growing vegetatively during inflorescence development, with 2-5 leaf pairs and 3-5 pairs of kataphylls above the node where the inflorescences originate in the axil of a bract, 4 mm long, linear with rounded apex kataphylls linear, with reddish branched hairs, pubescence especially dense adaxially, glabrous adaxially, inflorescence axes with basal lignified parts of inflorescences of previous years visible in robust well-collected specimens, allowing to observe growth and flower periodicity, these rests 8-16 cm apart, internodes between inflorescence pairs and leaf or kataphyll pairs characteristically short, medulla central in the branch, disappearing in older branches, leaving a hole only, inflorescence umbellate, buds not seen, in flowering stage in functionally male plants 10-11 cm wide, 5-6 cm high, with 8 main axes, in functional female plants 9-16 cm wide, 7-8 cm high, with 8-10 main axes, partial inflorescences cymes, secondary and tertiary inflorescence axes pubescent with reddish white stellate hairs; enlarged marginal flowers always absent; flower pedicel of reduced flowers 0.5-3 mm long in functionally male flowers, (0.2-)0.5-1(-5) mm long in functionally female flowers, receptacle campanulate with a well-defined rim in functionally male flowers, semiglobose in functionally female flowers, ovary inferior, calyx lobes 4, triangular in functionally male flowers, triangular with slightly rounded apex in functionally female flowers, in functionally male flowers 0.5-0.7 mm wide, 0.25-0.30 mm high, in functionally female flowers 0.4-0.5 mm wide, 0.2 mm long, petals 4, white, valvate, cucullate, triangular, apex rounded to acute, in functionally male flowers 1.4 mm long, 1.2 mm broad, in functionally female flowers 0.7-1.4 mm long, 0.8-1.5 mm broad, white; functionally male flowers: hypanthium 1.2-2 mm diameter, 0.9-1 mm high, stamens 8, well-developed, filaments 3 mm long, anthers 0.5-0.6 mm long, 0.3-0.4 mm broad, pistils 2, reduced, 0.5-0.8 mm long, conical, gradually tapering towards the apex, stigmas not penicellate; functionally female flowers: hypanthium 1.2-1.6 mm diameter, 1-1.2 mm high, stamens 8, reduced, filaments 0.2-0.4(-0.6) mm long, anthers 0.2-0.3 mm long, 0.2 mm broad, pistils 2, rarely 3, 1.3-1.5 mm long, stigmas apically and laterally clavate and shortly penicellate; fruits not seen.
Habitat: conserved cloud forest fragments exposed to the north. Additionally, the locality in Cintalapa in the state of Chiapas is an isolated mountain range between an area with hot and dry climate to the south and a zone characterized by a warm and humid climate to the north. The locality in Chimalapas in the state of Oaxaca is a secundary pine-oak forest according to the herbarium label, but we have not been able to visit this locality due to safety reasons.
Distribution: small populations in forest fragments of a few hectares, between 900 and 1250 m a.s.l. The particular sites where the species occurs are a geographical prominence or isolated geoformation with very high humidity levels.
Phenology: Hydrangea breedlovei has been collected in flower in the months of December and January.
Etymology: dedicated to Dr. Dennis Eugene Breedlove (1939-2012), the only botanist who, together with his collaborators, collected the genus Hydrangea in the field in southeastern Mexico. Thanks to these collections, we became aware of the occurence of several distinct Hydrangea species in the state of Chiapas and of the fact that the genus required thorough exploration work as well as a taxonomic revision.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. breedlovei is Endangered, wit an Extent of Occurence (EOO) of slightly over 3330 km2, and known from two populations. The habitat of the population in Berriozábal is severely threatened. The habitat of the population in Cintalapa is nearly entirely destroyed by pastures and coffee plantations.
Additional specimens examined: MEXICO. Chiapas, municipality Berriozábal, 13 km north of Berriozábal near Pozo Turipache and Finca El Suspiro, 900 m, 25.XII.1972, D. E. Breedlove and R. F. Thorne 30882 (CAS, DUKE, F, MICH, MEXU, MO) (functionally female); Vistahermosa, 1195 m, 16°50'57.01"N, 93°18'40.55"W, 21.VII.2011, M. S. Samain et al. 2011-202 (GENT, MEXU) (sterile); loc. cit., 31.XII.2011, M. S. Samain and E. M. Martínez Salas 2012-001 (CAS, CHIP, GENT, IEB, K, MEXU(2), MO) (functionally female); loc. cit., 12.II.2017, M. S. Samain and E. M. Martínez Salas 2017-007 (DUKE, IEB, MEXU(2), US, XAL) (functionally female). Municipality Cintalapa, between Colonia Francisco I. Madero and Colonia A. López Mateos, 1250 m, 31.XII.1980, D. E. Breedlove 49034 (DUKE, MEXU, MO) (functionally male); loc. cit., 1070 m, 16°49'23"N, 93°44'10.9"W, 28.XII.2009, C. Granados and G. Flores 487 (GENT, MEXU) (sterile); loc. cit., 1029 m, 16°49'10.87"N, 93°44'1.23"W, 2.I.2012, M. S. Samain et al. 2012-004 (CAS, CHIP, F, GENT, iBUG, IEB(3), K, MEXU(6), MICH, MO, XAL, US) (functionally male). Municipality La Trinitaria, 10 km east northeast of Dos Lagos above Santa Elena, 1170 m, 19.I.1987, D. E. Breedlove 57570 (DUKE) (functionally male). Oaxaca, district Juchitán, municipality Juchitán, Santa María Chimalapa, alrededores de Nuevo San Juan, without elevation data, 13.I.2000, E. Torres B. et al. 1941 (MEXU) (flower buds).
Observations: all specimens belonging to Hydrangea breedlovei had been erroneously identified as Hydrangea steyermarkii, with the exception of the specimen Torres et al. 1941 (MEXU) which had been identified as H. seemannii. The two latter species can be easily distinguished from H. breedlovei by the crenate leaf margin with marked glandular teeth, and the enlarged marginal flowers, respectively. Additionally, the three species occur in distinct geological provinces: limestone rocks of the Sierra Madre Oriental, the Central American volcanic system, and metamorphic rocks of the Sierra Madre Occidental, respectively
The collection made by D. E. Breedlove in La Trinitaria almost certainly concerns a labeling error with respect to the locality, although this cannot be checked anymore as there is no primary forest left in that area.
The label on the specimen D. E. Breedlove 57570 (DUKE) states “epiphytic vine”. This is not correct as none of the species of section Cornidia are epiphytic.
Material of the only known male individual of this species has been collected from the single adult individual we could find, despite meticulous exploration.
The locality in the state of Oaxaca should be visited as soon as safety issues allow in order to assess the conservation status of its population.
Hydrangea carroniae Samain & E. Martínez, sp. nov. Figs. 3, 5, 6.
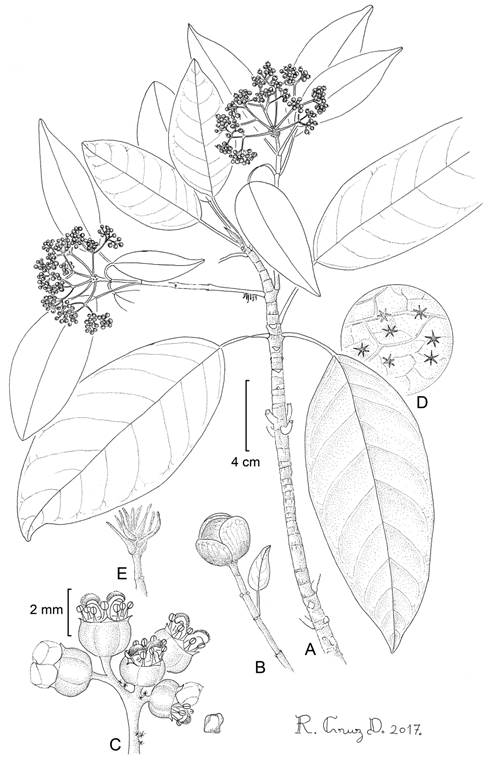
Figure 5: Hydrangea carroniae Samain & E. Martínez: A. branch with inflorescences; B. inflorescence bud; C. functionally female flowers; D. trichomes on abaxial leaf side; E. branch apex. M. S. Samain et al. 2011-003 (holotype: MEXU). Drawing by Ramiro Cruz Durán.

Figure 6: Distribution of Hydrangea carroniae Samain & E. Martínez, H. nebulicola Nevling & Gómez Pompa, and H. otontepecensis Samain & E. Martínez in Mexico. Map by Rosario Redonda Martínez.
TYPE: MEXICO. Veracruz, municipality Soteapan, Santa Martha, 1.5 km NNE of Santa Martha near foot path to La Ventana, 1218 m, 18°21'09.9"N, 94°53'21.2"W, 23.I.2011, M. S. Samain et al. 2011-003 (holotype: MEXU!, isotypes: GENT!, IEB!, K!, MEXU(2)!, MO!, US!) (functionally female).
Functionally dioecious species endemic to the area of Los Tuxtlas, Veracruz, Mexico, which can be distinguished from Hydrangea nebulicola by the apical shrub-like growth above its host tree canopy, glabrous and more coriaceous mature leaves, flat lamina, and much less pubescent inflorescence axes.
Root-climbing liana of up to 30 m high, apically growing shrub-like above its host tree canopy, functionally dioecious; main stem clinging to the host tree with many reddish adventitious roots, up to 15 cm diameter, bark greyish brown, with lenticels; runner shoots (stolons) reddish brown, with adventitious roots, nearly glabrous with only a few simple reddish hairs; branches many-ribbed, slightly quadrangular, attached to host tree with adventitious roots, these roots fibrous, slightly flattened, glabrous, free-growing branches terete, pubescent, brownish hairs stellate, deciduous, slightly fissured; leaves decussate, petiole sulcate adaxially, terete and slightly corchy abaxially, this corchy line sometimes continuing in the abaxial midvein, colour greyish brown, nearly glabrous with only a few simple reddish hairs, 1.5-4 cm long, leaving a triangular scar on the branch when leaves fall (scar sometimes with slightly rounded borders); lamina flat, ovate to lanceolate, 7-17 cm long, 3-8 cm broad, base cuneate to slightly rounded, apex slightly acuminate, leaf margin entire, revolute, thick, with a marginal vein, venation brochidodromous, veins 6-7 pairs, adaxial leaf side with slightly sunken midvein and secondary veins, tertiary veins invisible or only very lightly visible, secondary veins join to form submarginal or intramarginal vein, glabrous to pubescent with deciduous short, simple, white hairs, abaxially with protruding veins, sparsely pubescent, hairs stellate, whitish or reddish, inserted on top of a gland, deciduous, adnate, opaque whitish-green, acarodomatia triangular to elongated, without indument; inflorescence axis ribbed, with pubescence consisting of reddish stellate hairs, 7.5-8 cm long, with 3-5 opposite or decussate leaf pairs and 1-3 kataphyll pairs below the inflorescence, kataphylls lanceolate, petiole nearly absent to 5 mm long, 1-2 mm broad, lamina ovate, 1.8-2.5 cm long, 0.7-1.1 cm broad, starting from the base of the floral axis, each leaf pair with gradually diminishing size towards the inflorescence, apex of the floral axis woody, pyramidal, elongated bract scars visible, thickening at the top, 2.5-4 mm broad, 2.5 mm high in functionally female plants (measured in flowering stage only), 3.5-6 mm broad, (2.5-) 4.5-5 mm high in functionally male plants (measured in plant with old inflorescences only), inflorescence bracts cucullate with acuminate apex, reddish brown, membranous, adaxially slightly pubescent, abaxially densely pubescent with brownish stellate hairs, veins nearly invisible because of pubescence, bracts increasing in size, lowermost bract 0.5 cm large, 0.8 cm broad, higher bracts up to 2 cm large, 2.5 cm broad, consecutively and rapidly deciduous during inflorescence development, inflorescences lateral, opposite, one pair of inflorescences per flowering branch, rarely 3 or 4 inflorescences per flowering branch, flowering branch only continues growing vegetatively very slowly during inflorescence development, with maximum three leaf pairs and 3-5 kataphyll pairs above the inflorescences, inflorescence axes with basal lignified parts of inflorescences of previous years visible in robust well-collected specimens, allowing to observe growth and flower periodicity, these rests 13-18 cm apart (but relatively constant within one specimen), the number of scars of decussate leaf pairs not clearly visible because of lignification, but number over 10-15, medulla central in the branch, disappearing in older branches, leaving a hole only, leaves at the base of the inflorescence absent, inflorescence umbellate, buds up to 2.5 cm broad and 2 cm high before opening, in flowering stage 6-10 cm diameter, 3.5-7 cm high, with 6-9 main axes in functionally male plants, 7-10 main axes in functionally female plants, partial inflorescences corymbs, secondary and tertiary inflorescence axes with scarce reddish stellate pubescence, sometimes locally dense; enlarged marginal flowers absent; flower pedicel of reduced flowers 0.5-2.5 mm long in functionally male flowers, (0-)1.5-2.5 mm long in functionally female flowers, receptacle broadly campanulate in functionally male flowers, semiglobose in functionally female flowers, ovary inferior, calyx lobes 4, triangular, 0.2 mm long, 0.8 mm broad, petals 4, greenish white, valvate, cucullate, margin revolute, membranous, 1.5 mm long, 1-1.2 mm broad, white; functionally male flowers: hypanthium 2 mm diameter, 1-1.5 mm high, 1-2.2 mm broad, stamens 10, well-developed, filaments 1.5-2.5 mm long, anthers 0.8 mm long, 0.5-0.6 mm broad, pistils 2, reduced, 0.4-1 mm long, stigmas not penicellate; functionally female flowers: hypanthium 2 mm diameter, 1.5 mm high, 1.5 mm broad, stamens 7-10, reduced, filaments 0.2-0.8 mm long, anthers 0.3-0.5 mm long, 0.3-0.4 mm broad, pistils 2, 1.5-2.5 mm long, stigmas apically clavate and shortly penicellate; mature fruits not seen.
Habitat: extremely humid cloud forests (the nearby meteorological station “Los Tuxtlas II”, located at approximately 5 km, has the highest registered rainfall in Mexico: 8000 mm/year, Á. Campos Villanueva, pers. comm.), on volcanic rocks.
Distribution: north-exposed slopes of the higher parts of the volcanoes in the Sierra de Los Tuxtlas, an isolated volcanic mountain range along the southeastern coast of the Gulf of Mexico in the state of Veracruz, between 950 and 1300 m elevation.
Phenology: Hydrangea carroniae has been collected in flower in January and March.
Etymology: this species is named in honour of Rita Carron of Gent, Belgium, who has generously supported the start of our Hydrangea field work in Mexico.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. carroniae is Endangered, with an AOO of only 16 km2, and severaly fragmented and continuously declining populations, as its habitat continues to be destroyed. Moreover, there have also been fires in the area.
Additional specimens examined: MEXICO. Veracruz, municipality Catemaco, hillside above Perla de San Martín, 892 m, 18°32'32.8"N, 95°07'38.3"W, 30.VII.2016, M. S. Samain and E. M. Martínez Salas 2016-002 (IEB, MEXU) (sterile); loc. cit., 886 m, 18°32'31.2"N, 95°07'42.5"W, 2.VIII.2016, M. S. Samain and E. M. Martínez Salas 2016-003 (MEXU) (sterile). Municipality San Andrés Tuxtla, Reserva Ecológica de Calería, Ruiz Cortines - Sihuapan, 1090 m, 18°31.328'N, 95°9.341'W, 19.I.2010, A. Campos et al. 6700 (MEXU, MEXU-Los Tuxtlas) (functionally male); Reserva Ecológica Calería - Ruiz Cortines - Belem Chico, 1124 m, 9.8 km al N de San Andrés Tuxtla, 18°30.631'N, 95°9.869'W, 19.I.2011, A. Campos and R. Coates 7178 (MEXU, MEXU-Los Tuxtlas) (inflorescence buds); border of the ejidos Calería and Ruiz Cortínez, 11.5 km NE of San Andrés Tuxtla, 1046 m, 18°31'21.7"N, 95°09'19.5"W, 5.VIII.2010, C. Granados et al. 497 (GENT, MEXU(2)) (sterile). Municipality Santiago Tuxtla, entre Cerro Tuxtla (cerro Vigía) y Cerro Blanco, 950 m, 24.I.1972, J. H. Beaman 5510 (CAS, F, MICH, NY, XAL) (functionally male). Municipality Soteapan, 1.2 km NE of Santa Martha on foot path to La Ventana, 1271 m, 18°21'04.4"N, 94°53'23.7"W, 7.I.2010, C. Granados and A. Campos 489 (GENT, MEXU, MEXU-Los Tuxtlas) (functionally female); 1.3 km NE of Santa Martha on foot path to La Ventana. 1209 m, 18°21'8.7"N, 94°53'21.2"W, 7.I.2010, C. Granados and A. Campos 490 (GENT, MEXU) (sterile); 1.2 km NE of Santa Martha on foot path to La Ventana, 1271 m, 18°21'04.4"N, 94°53'23.7"W, 8.VIII.2010, C. Granados et al. 498 (GENT, MEXU) (sterile); loc. cit., 23.I.2011, M. S. Samain et al. 2011-002 (GENT, MEXU) (sterile).
Observations: this species has been confused with H. nebulicola, from which it can easily be distinguished by the much less dense indument, the pubescence in H. nebulicola covering different structures so densely that the details of these cannot be observed.
Hydrangea albostellata is known from two collections in the same area (see above) but this species can be easily distinguished by the dense white pubescence, the slightly undulated-spoon-shaped lamina and the thinner leaf texture.
In the locality of Perla de San Martín, municipality of Catemaco, the local name “hierba de tuza” is given to this species; “tuza” refers to a small rodent of the family Geomyidae, pocket gophers.
Hydrangea nahaensis Samain & E. Martínez, sp. nov. Figs. 1, 7, 8.
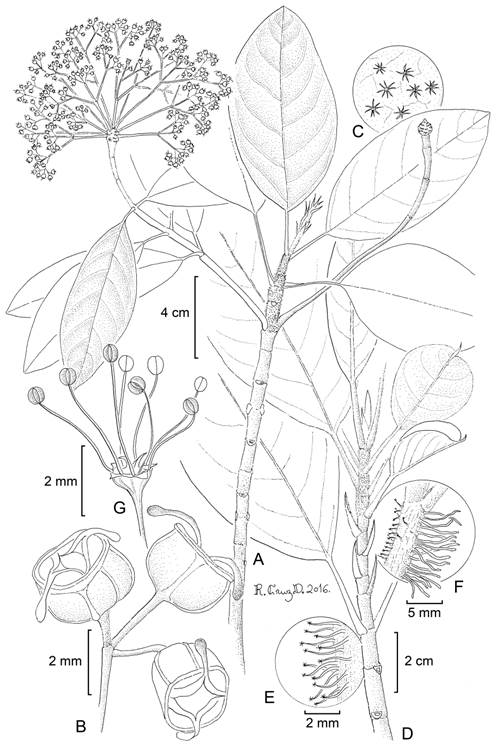
Figure 7: Hydrangea nahaensis Samain & E. Martínez: A. branch with inflorescences; B. fruits; C. indument on abaxial leaf side; D. branch with kataphylls; E. stem indument; F. stolon adventitious roots and trichomes; G. male flowers. A-C, M. S. Samain and E. M. Martínez Salas 2014-001 (holotype: MEXU); D, E, M. S. Samain et al. 2011-010 (MEXU); F, G, M. S. Samain and E. M. Martínez Salas 2012-019 (MEXU). Drawing by Ramiro Cruz Durán.

Figure 8: A, B, Hydrangea nahaensis Samain & E. Martínez: A. apical part of vegetative branch with kataphylls; B. apical part of flowering branch with left the rest of the woody inflorescence axis and right a young female inflorescence; C-F, H. nebulicola Nevling & Gómez Pompa: C. main stem clinging to its host tree with many adventitious roots; D. apical part of a flowering branch, with dense reddish indument, inflorescence bracts and base of inflorescences; E. flowering branch with male inflorescences; F. female inflorescence. Photos by Marie-Stéphanie Samain and Esteban Manuel Martínez Salas.
TYPE: MEXICO. Chiapas, Ocosingo, 3.51 km SSW of Nahá, Área de Protección de Flora y Fauna de Nahá, 944 m, 16°56'56.6"N, 91°35'33.0"W, 20.II.2014, M. S. Samain and E. M. Martínez Salas 2014-001 (holotype: MEXU!, isotypes: AAU!, C!, CAS!, CHIP!, CIIDIR!, CR!, F!, GH!, GENT!, IBUG!, IEB(3)!, K!, MEXU(3)!, MICH!, MO!, NY!, QMEX!, US!, XAL!) (functionally female).
Functionally dioecious species endemic to the Lacandon forest in the state of Chiapas (Mexico) which can be distinguished from Hydrangea steyermarkii by the ovate to elliptic or slightly lanceolate leaves, entire leaf margin, (2-)3-4 decussate leaf pairs below the inflorescence, and the presence of 5-7 kataphyll pairs on the flowering branch which continues growing during inflorescence development.
Root-climbing liana of up to 50 m high, branching above its host tree canopy or only reaching the lower branches of its host tree canopy and bending downwards, functionally dioecious; main stem clinging to the host tree with many adventitious roots, up to 20 cm diameter, bark dark brown, with lenticels, exfoliating when older; runner shoots (stolons) ribbed, reddish, with white adventitious roots and brown, stellate hairs, with few kataphylls and regular leaves, leaf pairs 5-7 cm distant from each other, 8 cm long, 3 cm broad, acarodomatia present (see below for detailed description); branches terete, slightly quadrangular at the height of the inflorescences, with many petiole scars, pubescent with simple hairs above the inflorescences, branches attached to host tree with adventitious roots, these roots fibrous, slightly flattened, with deciduous stellate indument, free-growing branches terete, very brittle when dry, rough because of the many leaf scars, fissured with many lenticels; leaves decussate, petiole sulcate adaxially, terete, with a thin corchy line, grey to reddish brown, when young slightly pubescent, when mature more pubescent, pubescence deciduous when older, consisting of whitish simple erect hairs and stellate adpressed hairs, 1-2(-3) cm long, leaving a triangular scar with rounded borders and 4 vascular bundles at each side on the branch when leaves fall; lamina flat, ovate to elliptic or slightly lanceolate, 5-18 cm long, 2.5-8 cm broad, base acute to rounded, apex acute, leaf margin entire, venation eucamptodromous, veins 4-8 pairs, adaxial leaf side with midvein slightly sunken at the base, slightly protruding in the basal half of the lamina, barely visible in the apical portion, secondary veins marked at the base, rest barely visible, tertiary veins very slightly visible, glabrous, green, abaxially with protruding veins, especially the primary ones, relatively densely pubescent with adpressed yellowish-brownish-reddish stellate hairs, reddish green, young leaves slightly pubescent, slightly reddish, indument deciduous, acarodomatia very small, consisting of a small cavity without hairs in the axils of the midvein and primary veins, in the lower 2/3 of the lamina, occasionally a few stellate hairs at the entrance of the cavity; inflorescence axis pubescent with dark brown branched hairs, base densely pubescent with both dendroid and stellate hairs, ribbed, 14 cm long, robust and gradually broadening towards the apex in both sexes, with (2-)3-4 decussate leaf pairs below the inflorescence, petiole (1-)1.5-2 mm long, 1.5-2 mm broad, sulcate adaxially, pubescent with adpressed stellate hairs and few simple hairs, leaves deciduous, elliptic, lamina 4-10 cm long, 2.5-4.5 cm broad, these leaves within a specimen of a similar size, apex of the floral axis woody, spherical, elongated to slightly conical, elongated bract scars visible, 6-8 mm broad, 6-8 mm high in functionally female plants, 5 mm broad, 5 mm high in functionally male plants, inflorescence bracts cucullate, reddish, coriaceous, margin membranous, adaxially slightly pubescent, especially at the base, few hairs at the apex, abaxially densely pubescent with reddish stellate hairs, veins nearly invisible because of pubescence, bracts increasing in size, lowermost bract 1.8 cm large, 0.9 cm broad, higher bracts up to 2.5 cm large, 1.4 cm broad, consecutively and rapidly deciduous during inflorescence development, inflorescences lateral, decussate, 1-3 pairs of inflorescences per flowering branch, flowering branch continues growing vegetatively during inflorescence development, with 5-6 leaf pairs and 5-7 kataphyll pairs above the inflorescences, all decussate, kataphylls linear, profoundly wedge-shaped, adaxially glabrous, adaxially densely pubescent, with reddish stellate hairs, 0.7-1 cm long, apical kataphylls forming a transition to the leaves, inflorescence axes with basal lignified parts of inflorescences of previous years visible in robust well-collected specimens, allowing to observe growth and flower periodicity, these rests 12-16 cm apart, with 4-5 decussate leaf pairs, up to 7 decussate kataphyll pairs (or the scars of these leaves and kataphylls) and sometimes again up to 4 decussate leaf pairs, medulla central in the branch, leaving a hole only, leaves at the base of the inflorescence axis absent, inflorescence corymbous, buds not seen, in flowering stage 8-10 cm wide, 7 cm high, with 4-10 main axes in functionally male plants, 5 main axes in functionally female plants (most inflorescences were already broken, so this could not be well observed), partial inflorescences cymes, secondary and tertiary inflorescence axes slightly pubescent with slightly reddish hairs, these hairs white on older axes; enlarged marginal flowers absent; flower pedicel of reduced flowers 0.3-1 mm long in functionally male flowers, 1-2 mm long in functionally female flowers, receptacle campanulate in functionally male flowers, semiglobose in functionally female flowers, ovary inferior, calyx lobes 4, triangular, sometimes slightly visible, 0.2 mm long, 0.5 mm broad, petals 4, white, valvate, cucullate, apex acuminate, margin and central vein slightly thicker, membranous, slightly transparent, 2 mm long, 1-2 mm broad, white; functionally male flowers: hypanthium 1.3-1.5 mm diameter, 1 mm high, stamens 6, well-developed, filaments (1.5-)3-4(-5) mm long, anthers 0.5-0.6 mm long, 0.5-0.6 mm broad, pistils 2, rarely 3, reduced, 0.6 mm long, stigmas not penicellate; functionally female flowers: hypanthium 2.3-2.7 cm wide, 1.2-2.3 cm high, stamens not observed as they shed quickly, pistils 2, rarely 3, 2 mm long, stigmas apically clavate and penicellate in horseshoe shape; fruit a semiglobose capsule with 6 lateral ribs, apically with a conspicuous border, brownish, 1.5-2 mm high, 2.7-2.9 mm broad above, 1.8-2 mm diameter, opening between the pistils to release seeds, seeds reddish, elongated, with pointed apices, 0.7 mm long, 0.10-0.15 mm thick.
Habitat: this species grows in the ecotone of of cloud forest to tropical rainforest, in flat or slightly inclined localities with well-developed, profound soil, and characterized by a mediane rainfall and humidity (2000-2500 mm). It climbs on several tree species, e.g. Magnolia lacandonica A. Vázquez, Pérez-Farr. & Martínez-Camilo, Lauraceae, Quercus spp., and Hedyosmum mexicanum C. Cordem. The protected area where it grows is a glen (“cañada”) with very slight slopes wich are exposed to the north. The contiguous elevations of the area are very small with a maximum difference of 100 m.
Distribution: this species is endemic to the Área de Protección de Fauna y Flora de Nahá y Metzabok in the Lacandon forest in the state of Chiapas. Individuals of this species grow allover the protected area where its habitat is well-conserved, and wherever neither deforestation or fires have occurred, between 830 and 1130 m elevation. The localities of all observed individuals are north-exposed. The area is characterized by a high and constant humidity along the year (pattern of the Gulf of Mexico), and suffers strong effects of trade winds, or the so-called “nortes”, during the winter season.
Phenology: Hydrangea nahaensis has been collected in flower in January and February.
Etymology: this species is dedicated to the community of Nahá, core of the Maya-Lacandon ethnicity, who conserve their surrounding forest by their own decision and as part of their tradition. Moreover, the authorities of the Comisión Nacional de Áreas Naturales Protegidas (CONANP) have staff in the area who performs different conservation and reforestation projects in cooperation with the community of Nahá and neighbouring communities.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), Hydrangea nahaensis is Critically Endangered as it has an Extence of Occurence (EOO) of slightly over 8 km2 and is known from only one population in a fragile habitat which despite serious conservation efforts runs the constant risk of being destroyed, e.g. by fires. Moreover, we cannot distinguish between individuals in a particular locality as all plants seem to be connected by runner shoots (see also in the discussion about this observation), so the actual number of individuals is probably much lower than estimated. In fact, we observed only one fertile female individual.
Additional specimens examined: MEXICO. Chiapas, municipality Ocosingo, Nahá, Área de Protección de Flora y Fauna de Nahá, 5 km S of Nahá on the way to Jopeta (Cabeza de la Laguna), 952 m, 16°56'55.3"N, 91°35'34.4"W, 22.VII.2010, C. Granados et al. 494 (GENT, MEXU) (sterile); Área de Protección de Flora y Fauna de Nahá, 2.09 km W of CONANP office, 833 m, 16°58'42.4"N, 91°36'06.2"W, 22.VII.2010, M. S. Samain et al. 2010-092 (GENT, MEXU) (sterile); loc. cit., 2.22 km W of CONANP office, 865 m, 16°58'40.9"N, 91°36'10.6"W, 22.VII.2010, M. S. Samain et al. 2010-093 (GENT, MEXU) (sterile); loc. cit., 2.07 km W of CONANP office, 828 m, 16°58'41.3"N, 91°36'05.5"W, 22.VII.2010, M. S. Samain et al. 2010-094 (GENT, MEXU) (sterile); loc. cit., 5.37 km SW of CONANP office, 1072 m, 16°57'22.0"N, 91°36'17"W, 22.VII.2010, M. S. Samain et al. 2010-095 (GENT, MEXU) (sterile); loc. cit., 5.37 km SW of CONANP office, 1119 m, 16°57'20.1"N, 91°36'21.4"W, 22.VII.2010, M. S. Samain et al. 2010-096 (GENT, MEXU) (sterile); loc. cit., 5.47 km SW of CONANP office, 1129 m, 16°57'19.9"N, 91°36'24.8"W, 22.VII.2010, M. S. Samain et al. 2010-097 (GENT, MEXU) (sterile); loc. cit., 0.9 km ENE of CONANP office, 1055 m, 16°58'55.9"N, 91°34'29.9"W, 23.VII.2010, M. S. Samain et al. 2010-098 (GENT, MEXU) (sterile); loc. cit., 1.94 km ESE of CONANP office, 960 m, 16°58'32.2"N, 91°34'30.6"W, 23.VII.2010, M. S. Samain et al. 2010-099 (GENT, MEXU) (sterile); loc. cit., 0.88 km ESE of CONANP office, 993 m, 16°58'30.1"N, 91°34'27.5"W, 23.VII.2010, M. S. Samain et al. 2010-100 (GENT, MEXU) (sterile); loc. cit., 2.09 km SW of CONANP office, 942 m, 16°57'30,6"N, 91°35'37,0"W, 29.I.2011, M. S. Samain et al. 2011-010 (CAS, GENT, iBUG, IEB, K, MEXU, MO, US, XAL) (functionally male); loc. cit., 2.5 km SSW of Nahá, 961 m, 16°57'31.83"N, 91°35'36.36"W, 17.I.2012, M. S. Samain and E. M. Martínez Salas 2012-019 (BIGU, CAS, CHIP, CIIDIR, CR, F, GENT, GH, IBUG, IEB, K, MEXU(2), MICH, MO, NY, QMEX, US, XAL) (functionally male).
Observations: this is the largest continuous population of any Hydrangea species we have observed in Mexico. Actually, this species had never been collected before we started this project. Moreover, it is the most viable and healthy population we have visited as many juvenile branches creep on the substrate towards other host trees. Perhaps as a consequence of its size, the flowering period of the population as a whole lasts up to one month, whereas much smaller populations of other Hydrangea species only flower during one week. Likewise, in contrast to the other Mexican species we observed more variation within H. nahaensis, as we were able to study individuals across their distribution range, whereas in rarer species, we only saw a small number of remaining individuals, that possibly do not reflect anymore the complete former morphological variation of the species.
Hydrangea nebulicola Nevling & Gómez Pompa, J. Arnold Arbor. 49: 231. 1968, amended description. Figs. 6, 8.
TYPE: MEXICO. Veracruz, municipality Yecuatla, Sierra de Chiconquiaco y Misantla, 1540 m, 9.VII.1966, A. Gómez-Pompa 1541 (holotype: MEXU!, isotypes: CAS!, GH!, MEXU(4)!, MO!, XAL!) (young functionally male).
Root-climbing liana of up to 30 m high, never reaching above the lower branches of its host tree canopy, sometimes bending downwards, functionally dioecious; main stem clinging to the host tree with many adventitious roots, up to 20 cm diameter, bark dark brown, with lenticels; runner shoots (stolons) reddish brown, with reddish brown adventitious roots and stellate hairs, with small decussate leaves; branches many-ribbed, slightly angular, branches attached to host tree with adventitious roots, these roots fibrous, slightly flattened, with deciduous stellate indument, free-growing branches terete, slightly fissured, kataphylls or their scars visible, allowing to observe growth periodicity, although the distance between them varies, even within a single individual; leaves decussate, petiole sulcate adaxially, terete and abaxially, colour reddish brown, densely pubescent with mostly reddish and a few white stellate hairs, 1-3 cm long, leaving a semicircular scar on the branch when leaves fall; lamina spoon-shaped, ovate to lanceolate-elliptic, 7-23 cm long, 3-10 cm broad, base cuneate to rounded, apex acute to acuminate, leaf margin entire, venation brochidodromous, veins 5-9 pairs, adaxial leaf side with midvein slightly protruding in the basal 2/3 of the lamina, upper portion marked, primary and secondary veins lightly marked, primary veins join to form submarginal or intramarginal vein, nearly glabrous with the exception of the basal third of the midvein which has dense reddish stellate pubescence as the petiole, abaxially with protruding veins, opaque olive green, densely pubescent with stalked stellate reddish hairs, slightly reddish, especially on the veins, acarodomatia present but often not very conspicous as they may lay hidden under de midvein pubescence, in axils of midvein and primary veins, sometimes partially hidden under de primary vein which may be broadened at its base; inflorescence axis densely pubescent with adnate, reddish, stellate hairs, 3-15 cm long, with 1-3 opposite or decussate leaf or kataphyll pairs below the inflorescence, deciduous, petiole 3-6 mm long, 2 mm broad, lamina 1-5 cm long, 1-2.5 cm broad, kataphylls reddish, densely pubescent, up to 1.5 mm long, apex of the floral axis woody, cone-shaped, elongated bract scars visible, thickening at the top, 2-3 mm broad, 1-3 mm high in functionally female plants, 2-4 mm broad, 3 mm high in functionally male plants, inflorescence bracts cucullate, green, light brown to reddish, membranous to coriaceous, adaxially and abaxially slightly to densely pubescent with whitish or reddish simple and stellate hairs, veins darker, bracts increasing in size, lowermost bract 1 cm large, 0.7 cm broad, higher bracts up to 2 cm large, 1.5 cm broad, consecutively and rapidly deciduous during inflorescence development, bracts of partial inflorescences linear, reddish, 2.5 mm long, 0.8 cm broad at the base, gradually tapering towards a pointed apex, deciduous, inflorescences lateral, opposite, 1-4 pairs of inflorescences per flowering branch, sometimes only one inflorescence developing, flowering branch only continues growing vegetatively extremely slowly during inflorescence development, with just 2-4 kataphyll pairs above the inflorescences, linear, with dense reddish stellate hairs, inflorescence axes with basal lignified parts of inflorescences of previous years visible in well-collected specimens, allowing to observe growth and flower periodicity, these rests 6-9 cm apart (but relatively constant within one specimen), with 7-9 decussate leaf pairs (or the scars of these leaves, this number constant within one specimen), the first 4-5 leaf pairs (mostly only visible as scars) notoriously close to each other, medulla central in the branch, disappearing in older branches, leaving a hole only, kataphylls at the base of the inflorescence present, 1-1.5 cm long, inflorescence umbellate, buds up to 3 cm broad and 2.5 cm high before opening, in flowering stage 4-5 cm diameter, 3-4 cm high, with 5-9 main axes in functionally male plants, 7-9 main axes in functionally female plants, partial inflorescences umbels, secondary and tertiary inflorescence axes with reddish simple and stellate hairs, pubescence gradually decreasing towards flower insertion; enlarged marginal flowers sometimes present, terminally placed in a cyme, sepals with marked veins, pistils 2, stigma not developed, further characters not observed in detail; flower pedicel of reduced flowers 1-3 mm long in functionally male flowers, 1-2 mm long in functionally female flowers, receptacle broadly campanulate in functionally male flowers, semiglobose in functionally female flowers, ovary inferior, calyx lobes 4, triangular, 0.3-0.5 mm long, petals 4, white, valvate, cucullate, membranous, 1.5-2 mm long, 1.5 mm broad, white; functionally male flowers: hypanthium 1.5-2.5 mm diameter, stamens 6-8, well-developed, filaments 2.3-3 mm long, anthers 0.4-1 mm long, 0.3-0.5 mm broad, pistils 2, reduced, 0.4-0.5 mm long, stigmas not penicellate; functionally female flowers: hypanthium 1.5-2 mm diameter, stamens 5-9, reduced, filaments 0.4-1.6 mm long, anthers 0.2-0.7 mm long, 0.3-0.4 mm broad, pistils 2, 1.5-2 mm long, enlarging up to 2.5 mm during fruit maturation, stigmas apically clavate and shortly penicellate; fruit a semiglobose capsule, apically with a conspicuous border, dark reddish brown, 1.5-2 mm high, 2-2.5 mm broad above, 2-2.5 mm diameter, opening between the two pistils to release seeds, seeds not seen.
Habitat: well-conserved cloud forests, Liquidambar forests and oak-Liquidambar forests, with a very high relative humidity all year round, on moderate to very steep north-facing slopes, on rocks of volcanic origin or metamorphic rocks. In the state of Puebla, this species grows near the ridge of valleys with permanent water watercourses, with cloud forest elements, surrounded by temperate dry oak and pine forests, on basaltic substrate.
Distribution: this species is known from isolated populations in the highly diverse floristic region known as La Huasteca in the states of Hidalgo, Puebla, Querétaro and Veracruz. It occurs in two areas in the heart of the Huasteca: the northern slopes of the Sierra de Chiconquiaco in Veracruz, directly exposed to the northern winds, between 1080 and 1600 m a.s.l., and on conserved massifs in the north and center of the state of Hidalgo, between 1500 and 1900 m elevation. Near the western limits of the Huasteca region, in Querétaro near the border with the state of Hidalgo, a population is known from the Sierra Gorda, an isolated mountaineous area in the system of the Sierra Madre Oriental, between 1350 and 1400 m elevation. Finally, two small populations occur near the southern margins of the Huasteca in the zone of Zacatlán and Chignahuapan in the state Puebla, in a valley with NE-SW orientation, between 2000 and 2200 m a.s.l.
Phenology: Hydrangea nebulicola has been collected in flower from July to September.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. nebulicola is Endangered, with an AOO of 40 km2, despite an EOO of nearly 20000 km2, as a consequence of habitat destruction allover its distribution area. The populations are very small. Hydrangea nebulicola was already considered as threatened or endangered more than thirty years ago in one of the first ever contributions on plant extinction (Vovides and Gómez-Pompa, 1976). Moreover, the species was listed by the Mexican authorities as endangered in 1994 (NORMA Oficial Mexicana NOM-059-ECOL-1994, INE, 1994) and as threatened with extinction since 2001, NOM-059-SEMARNAT-2001 (SEMARNAT, 2002) and NOM-059-SEMARNAT-2010 (SEMARNAT, 2010). The only known small population in Querétaro of this species consists of few adult and juvenile individuals. The small population of the Cascada (waterfall) de San Pedro in Zacatlán is probably relatively well-protected as it is a small-scale ecotouristic place. Nevertheless, the continuous presence of the waterfall should be ensured, as from the nearby locality of Chignahuapan, it can be seen that the decrease of humidity causes the plants to reduce development and growth. With respect to the locality of Cascada Quetzalapan in the municipality of Chignahuapan, it will be essential that the community lets pass some water near the plant (the water tubes are still present), in order to ensure its survival.
Additional specimens examined: MEXICO. Hidalgo, municipality Eloxochitlán, road Ixmolintla - Almoloya, km 10.5, 1881 m, 20°45'33.75"N, 98°46'30.24"W, 15.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-195 (GENT, IEB, MEXU) (functionally male); loc. cit., 15.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-196 (CAS, CHIP, F, GENT, GH, IBUG, IEB(2), K, MEXU(3), MO, QMEX, XAL) (functionally male). Municipality Molango de Escamilla, 8-9 kms al norte de Ixmolintla (written as Ismolinda), 1900 m, 27.IV.1981, R. Hernández M. et al. 5913 (MEXU) (sterile). Municipality Tlanchinol, 234 m NE of Rancho El Temazate, 1509 m, 21°1'42.22"N, 98°38'26.27"W, 14.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-194 (CHIP, GENT, HUAP, iBUG, IEB(3), K, MEXU(4), MO, NY, QMEX, XAL) (functionally male). Puebla, municipality Chignahuapan, Cascada Quetzalapan, 2045 m, 19°52'19.85"N, 97°58'50.97"W, 8.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-192 (GENT, MEXU) (sterile). Municipality Zacatlán, Cascada de San Pedro, 2170 m, 19°52'19.85"N, 97°58'50.97"W, 7.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-188 (GENT, IEB, MEXU, MO) (functionally male); loc. cit., 7.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-189 (DUKE, F, GENT, HUAP, IEB, MEXU) (functionally male); loc. cit., 7.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-190 (GENT, IEB, MEXU(2)) (functionally male); loc. cit., 25.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-215 (GENT, IEB, F, K, MEXU) (functionally male); loc. cit., 25.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-216 (GENT, MEXU) (functionally male); loc. cit., 11.IX.2012, M. S. Samain and E. M. Martínez Salas 2012-092 (GENT, GH, HUAP, IEB, MEXU) (functionally male). Querétaro, municipality Landa de Matamoros, 300 m S of El Pemoche on the road to Agua Zarca, 1347 m, 21°13'46.1"N, 99°06'47.5"W, 27.VII.2010, C. Granados et al. 495 (GENT, MEXU) (inflorescence buds); 4 km NW of Agua Zarca, 1374 m, 21°13'54.3"N, 99°06'49.4"W, 27.VII.2010, C. Granados et al. 496 (GENT, MEXU) (sterile); al sur de El Pemoche, 4.X.1997, E. Pérez and E. Carranza 3757 (IEB, MEXU) (functionally male); 1 km al sur de El Pemoche, 1300 m, 7.IX.1998, E. Pérez and E. Carranza 3875 (IEB, MEXU, XAL) (functionally male); 1.5 km al SW de El Pemoche, 1360 m, 29.VIII.1998, H. Rubio 1035 (IEB, MEXU, XAL) (functionally male); 4 km NW of Agua Zarca, 1 km al sureste de El Puerto del Sabino, 23.VIII.1991, H. Rubio 2559 (IEB, QMEX) (functionally male); 4 km al N de Agua Zarca, sobre el camino al Lobo, 13.IV.1988, J. Rzedowski 46378 (IEB) (sterile); 300 m S of El Pemoche near the road to Agua Zarca, 1347 m, 21°13'46.1"N, 99°06'47.5"W, 27.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-217 (CAS, F, GENT, IBUG(2), IEB(2), K, MEXU(5), MICH, MO(2), NY, QMEX) (functionally female); 1.5 km al SW de El Pemoche, 1360 m, 21.X.1997, S. Zamudio et al. 10562 (IEB) (sterile). Veracruz, municipality Yecuatla, Santa Rita, Sierra de Chiconquiaco, 1540 m, 9.XII.1967, A. Gómez-Pompa 1808 (GH, MEXU) (sterile); 0.5 km after Capulines on road from Chiconquiaco to Misantla, 1450 m, 19°49'25.7"N, 96°49'44.6"W, 9.I.2010, C. Granados and A. Campos 491 (GENT, MEXU) (sterile); curve “La Z”, 4 km after Santa Rita on road from Chiconquiaco to Misantla, 1081 m, 19°50'42.8"N, 96°49'14.3"W, 10.I.2010, C. Granados and A. Campos 492 (GENT, MEXU), 493 (GENT, MEXU) (sterile); Las Paredes dirección Gutiérrez Nájera, 1600 m, 19°49'48"N, 96°49'48"W, 10.IX.1990, C. Gutiérrez B. 4078 (MEXU. NY, XAL) (functionally male); Santa Rita, 29.III.1967, R. Hernández M. 306 (MEXU(2)) (sterile); Santa Rita, Sierra de Chiconquiaco, 1310 m, 16.VII.1967, L. Nevling and A. Gómez-Pompa 22 (GH, MEXU) (sterile); loc. cit., 1350 m, 16.VII.1967, L. Nevling and A. Gómez-Pompa 39 (GH(2), MICH) (inflorescence and flower buds, it cannot be confirmed that the partial male inflorescence on one of the GH specimens is part of the same individual); loc. cit., 1540 m, 6.IX.1967, L. Nevling and A. Gómez-Pompa 583 (MEXU(3), GH) (functionally male); Sierra de Chiconquiaco, 1462 m, 19°49'29.59"N, 96°49'44.18"W, 24.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-214 (F, GENT, iBUG, IEB(2), K, MEXU(3), MO, XAL) (functionally female).
Observations: the original description by Nevling and Gómez-Pompa (1968) is based on a functionally male specimen and therefore details on fully developed female structures of functionally female specimen are lacking. These authors were, based on their observation of H. nebulicola, the first ones to hypothesise the occurrence of functionally unisexual flowers and dioecism in this genus.
This is one of the three Mexican Hydrangea species known by local people and with a vernacular name, the other ones being H. carroniae and H. seemannii. The local name given to this species in the state of Hidalgo is “hoja de zapote”, sapote leaf. Inhabitants of the village of San Miguel Tenango in Puebla name this plant “pagua xihuitl“ in Náhuatl, meaning similar to the leaf of avocado.
Collection M. S. Samain and E. Martínez Salas 2011-192 (GENT, MEXU), from the Cascada Quetzalapan in Chignahuapan, is made from an individual which is poorly developed compared to all other known plants, as it has smaller leaves and retarded flowering. It might even be that the very small inflorescence buds which have been observed do not develop further, due to the fact that the permanent water course coming from the hydro-electrical factory has stopped since about 10 years.
The stamens of collection M. S. Samain and E. Martínez Salas 2011-217 (CAS, F, GENT, IBUG(2), IEB(2), K, MEXU(5), MICH, MO(2), NY, QMEX) have relatively long filaments for a functionally female specimen, as a consequence the flowers could be erroneously interpreted as bisexual; however, the anthers are shriveled and thus do not contain pollen.
Hydrangea otontepecensis Samain & E. Martínez, sp. nov. Figs. 6, 9, 10.
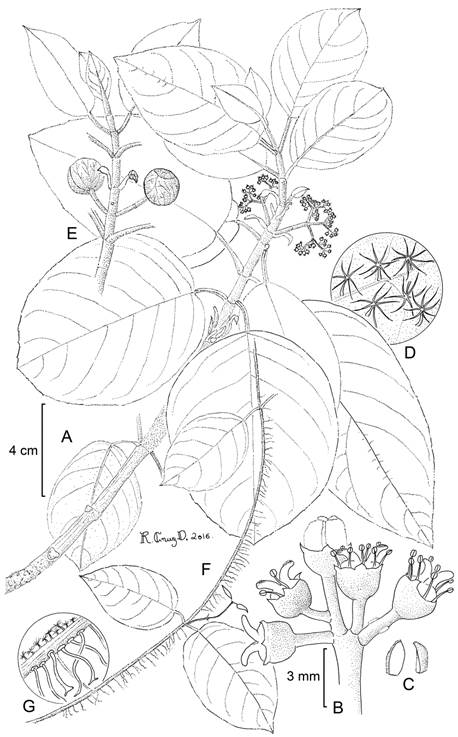
Figure 9: Hydrangea otontepecensis Samain & E. Martínez: A. branch with inflorescences; B. functionally female flowers; C. petals; D. trichomes on the abaxial leaf surface; E. branch with inflorescence buds; F. young branch with adventitious roots; G. stolon with adventitious roots and trichomes. M. S. Samain et al. 2011-193 (holotype: MEXU). Drawing by Ramiro Cruz Durán.

Figure 10: A, B, Hydrangea otontepecensis Samain & E. Martínez: A. young runner shoot climbing on host tree; B. apical part of flowering branch with inflorescence buds, inflorescence bracts and flower buds (right); C, D, H. seemannii L. Riley: C. young runner shoot with adventitious roots and reddish brown simple and stellate hairs; D. flowering individual on host tree; E, F, Hydrangea sousae Samain, Najarro & E. Martínez: E. individuals on host trees, apical branches growing about the tree canopy; F. female inflorescence with maturing fruits and one enlarged marginal flower. Photos by Marie-Stéphanie Samain and Esteban Manuel Martínez Salas.
TYPE: MEXICO. Veracruz, municipality Chontla, Sierra de Otontepec, NNE of San Nicolasillo, 1078 m, 21°14'47.67"N, 97°53'38.80"W, 13.VIII.2011, M. S. Samain et al. 2011-193 (holotype: MEXU!, isotypes: CHIP!, F!, GENT!, iBUG!, IEB!(3), K!, MEXU!(4), MO!, US!, XAL!(2)) (functionally female).
Functionally dioecious species endemic to the Sierra de Otontepec, Veracruz, Mexico, which can be distinguished from Hydrangea nebulicola by the dense whitish to yellowish lanuginose pubescence in the meristematic zones, flat lamina, leaf margin slightly dentate, kataphylls regularly not well-differentiated, and the comparatively much smaller inflorescences.
Root-climbing liana of up to 20 m high, functionally dioecious; main stem clinging to the host tree with many adventitious roots, 10-12 cm diameter, bark reddish brown, slightly fissured and exfoliant, with few lenticels; runner shoots (stolons) many-ribbed (up to 20), reddish brown, with whitish adventitious roots and reddish stellate hairs, with small leaves, internodes up to 10-15 cm long; branches many-ribbed, slightly angular, with many kataphyll scars, branches attached to host tree with adventitious roots, these roots fibrous, slightly flattened, with deciduous stellate indument, free-growing branches many-ribbed, ribs pronounced in young branches, slightly fissured and exfoliant, with rests of adventitious roots with white pubescence, more pubescent towards the apex, lenticels few, medulla central in the branch; leaves decussate, petiole slightly sulcate adaxially, terete, colour brown, densely pubescent, hairs stellate, white to brown, 1-2.5 cm long, leaving a triangular scar on the branch when leaves fall (bottom of the slightly rounded), remains of 6 vascular bundles slightly visible; lamina flat, elliptic, ovate or oblong, chartaceous, 3-(6-15) cm long, 1.5-(4-9) cm broad, base rounded, slightly asymmetrical, apex acute to slightly acuminate, leaf margin slightly dentate (teeth are not always placed where a vein ends in the margin), especially in young leaves, venation eucamptodromous, veins 5-7 pairs, adaxial leaf side with yellowish to reddish veins, midvein sunken to slightly protruding, densely pubescent in young leaves, secondary veins slightly sunken to well-marked, glabrous, tertiary veins not to partially visible, glabrous, (opaque olive) green, abaxially with reddish protruding densely pubescent midvein, secondary veins protruding, tertiary veins well-marked, sparsely pubescent resulting in a spongy effect, bright green to white-reddish (because of the indument), young leaves adaxially glabrous to pubescent (pubescence deciduous), abaxially with dense pubescence, this indument lanuginose, whitish to yellowish in very young leaves, more reddish while maturing, acarodomatia partially covered with reddish stellate hairs, present along the midvein but absent or very small in the apical third of the leaf, barely visible on young leaves because of the very dense indument; inflorescence axis densely pubescent with reddish, stellate hairs, 1-1.5 cm long, apex of the floral axis woody, cone-shaped, reduced, 2 mm high, 2-3 mm wide, 1.3 mm broad, inflorescence bracts cucullate, reddish, coriaceous, margin membranous, adaxially and abaxially densely pubescent with reddish stellate hairs, slightly darker coloured veins nearly invisible because of dense pubescence, bracts increasing in size, lowermost bract 0.7 mm high, 0.9 mm broad, higher bracts 1.5 mm large, consecutively and rapidly deciduous during inflorescence development, bracts scars not visible because of the indument, inflorescences lateral, opposite or decussate, main axis continues growing slowly vegetatively during inflorescence development, with 2-3 pairs of not well-differentiated kataphylls and sometimes 2-3 leaf pairs as well above the node where the inflorescences originate, inflorescence axes with basal lignified parts of inflorescences of previous years visible, allowing to observe growth and flower periodicity, these rests 12 cm apart, inflorescence umbellate, buds up to 1.5 cm broad and 1.5 cm high before opening, in flowering stage 4-5 cm wide, 2.5-3 cm high, 2-7 main axes, partial inflorescences cymes, all inflorescence axes with dense reddish deciduous stellate hairs (but less dense than the main inflorescence axis), bract at the basis of each partial inflorescence linear, 6 mm long, 2.5-3 mm broad at the base, apex rounded; enlarged marginal flowers always absent; functionally male flowers not seen, the following descriptions refer to functionally female flowers; flower pedicel of reduced flowers 0.5-2(-5) mm long, slightly pubescent with reddish erect stellate hairs, receptable semiglobose, with 2 lobes, ovary inferior, calyx lobes 4, rounded, 0.7-1 mm wide, 0.2-0.4 mm long, petals 4, greenish white, valvate, cucullate, triangular, apex acute, 1-1.5 mm long, 0.8-1 mm broad, white; hypanthium 1.2-1.6 mm diameter, 1.7-2.2 mm high, stamens 8, reduced, filaments 0.7-1.2 mm long, anthers 0.2 mm long, 0.2 mm broad, pistils 2, 1.3-1.8 mm long, stigmas papillate, green; mature fruits not seen.
Habitat: marginal cloud forest, surrounded by vegetation of hot and humid climates (oak forest and tropical rain forest). Only occuring in a small protected valley of the highest parts of the Sierra de Otontepec, an isolated mountain system that functions as an island.
Distribution: south-facing slope of the Sierra de Otontepec where it is protected from the cold winds coming from the northeast. The Sierra de Otontepec is a mountain range on the coastal plain of the Golf of Mexico, isolated from the Sierra Madre Oriental.
Phenology: Hydrangea otontepecensis has been collected in flower in August.
Etymology: this species is named after the Sierra Otontepec, to which this species is endemic.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. otontepecensis is Critically Endangered as it is only known from one small population with female individuals. Despite extensive exploration in the area, no male specimens could be found.
Observations: this species had been collected once before and this specimen (which we have not seen, see below) is mentioned in the Flora of Veracruz (Durán-Espinosa, 1999) as H. nebulicola. It can be distinguished from this species, based on the relatively smaller inflorescences (4-5 × 2.5-3 cm vs. 4-5 × 3-4 cm), the whitish to yellowish (vs. reddish) pubescence and flat (vs. spoon-shaped) lamina.
The specimen Castillo-Campos et al. 2508 (according to Durán-Espinosa (1999) deposited in XAL) is apparently not present in that herbarium collection and has thus not been seen by us).
Hydrangea seemannii L. Riley, Bull. Misc. Inform. Kew 1924(5): 207. 1924, amended description. Figs. 10, 11.
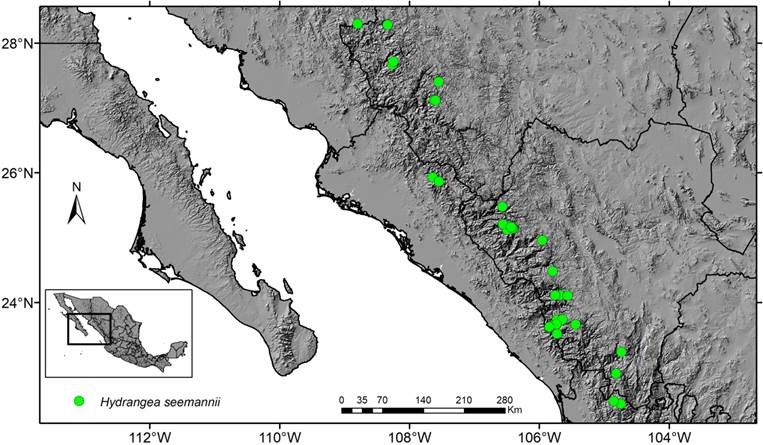
TYPE: MEXICO. Durango, municipality unknown, Sierra Madre, Rancho de Guadalupe, s.f., Seemann 2142 (holotype: K!, isotypes: BM!, GH!) (fruits).
Root-climbing liana of up to 60 m high, sometimes apically growing shrub-like above its host tree canopy or boulders on which it grows, monoecious with bisexual flowers; main stem clinging to the host tree or to rock walls and boulders with many adventitious roots, up to 20 cm diameter, bark dark brown, with lenticels; runner shoots (stolons) greyish to reddish, with adventitious roots and reddish brown simple and stellate hairs, with kataphylls or small leaves; branches many-ribbed, slightly angular, with many leaf scars, branches attached to host tree or other surfaces with adventitious roots, these roots fibrous, slightly flattened, free-growing branches terete, slightly fissured, exfoliating and with whitish stellate hairs; leaves decussate, petiole sulcate adaxially, terete abaxially, glabrous to scarcely pubescent with pale reddish brown simple and stellate hairs, 1-3 cm long, leaving a triangular scar with rounded, slightly elevated borders on the branch when leaves fall; lamina flat, ovate, lanceolate to elliptic, 4-13 cm long, 3-7 cm broad, base cuneate to decurrent, apex acuminate to acute, leaf margin entire, venation brochidodromous, veins 8-10 pairs, adaxial leaf side with midvein and primary veins slightly sunken, secondary veins nearly invisible, secondary veins join to form submarginal or intramarginal vein, glabrous, abaxially with protruding midvein and secondary veins, tertiary veins slighty marked, very scarce white or seldom reddish stellate pubescence, rapidly deciduous, these hairs inserted on reddish dots, opaque olive green, acarodomatia present in axil of midvein and secondary veins, covered with tufts of whitish or slightly reddish stellate hairs; inflorescence axis nearly glabrous to densely pubescent with adnate, whitish or reddish stellate hairs, 5-20 cm long, with 6-7 opposite or decussate leaf pairs below the inflorescence, rapidly deciduous, petiole 1-3 cm long, 1-2 mm broad, lamina 5-12 cm long, 3-7 cm broad, starting from the base of the floral axis, each leaf pair with gradually diminishing size towards the inflorescence, apex of the floral axis woody, cone-shaped, elongated bract scars visible, 2-4 mm broad, 3-4 mm high, inflorescence bracts from outside to inside triangular, elliptic and cucullate, respectively sometimes with apiculate apex, reddish, coriaceous, border membranous, adaxially glabrous to slightly pilose with mostly reddish, rarely whitish stellate hairs, abaxially glabrous to densely pilose, external ones generally densely pilose abaxially with shortly ciliate margin, internal ones becoming more glabrous abaxially towards the center of the inflorescence bud, veins strikingly marked, especially on the internal bracts, bracts increasing in size, lowermost bract 1.1-1.8 cm large, 0.2-1.5 cm broad, higher bracts up to 1.5-2 cm large, 0.6-1.5 cm broad, consecutively and rapidly deciduous during inflorescence development, bracts of partial inflorescences linear, reddish, glabrous to adaxially slightly pilose with stellate hairs at the apex, 1.6-2.5 mm long, 1.5-1.8 mm broad, apex rounded, margin slightly crenate from the central part to the apex, deciduous, inflorescences lateral, opposite, 1-2(-3) pairs of inflorescences per flowering branch, sometimes one inflorescence developing only, flowering branch only continues growing quickly during inflorescence development, with pairs of leaf of normal size, however, this branch never reaching above the inflorescences, inflorescence axes with basal lignified parts of inflorescences of previous years not observed, medulla central in the branch, disappearing in older branches, leaving a hole only, leaves at the base of the inflorescence absent, inflorescence umbellate, buds up to 1.6-3.5 cm broad and 2-3.2 cm high before opening, in flowering stage 4-20 cm diameter, 4-15 cm high (both these measurements including enlarged marginal flowers), 4-10 main axes partial inflorescences cymes with enlarged marginal flowers terminally placed in each cyme, secondary and tertiary inflorescence axes glabrous; enlarged marginal flowers nearly always present, rarely absent, flower pedicel 2-4 cm long, receptacle sometimes nearly invisible, but mostly semiglobose to campanulate (both may occur within one individual), with 4 lateral ribs, up to 3-4 mm high, 1-2 mm broad, 1-2 mm diameter, ovary inferior, sepals 4(-5), yellowish to greenish, coriaceous with marked veins, ovate to obovate, margin membranous, 1.2-2 cm long, 1.2-2 cm broad, petals reduced and deciduous, 1.5-2 mm long, 1-1.3 mm broad, stamens 6, often fertile, filaments reduced and flattened, 0.8-1.5 mm long, 0.2 mm broad, anthers 0.3-0.5 mm long, pistils (1-)2, style 1-2 mm long, stigma spathulate to rounded, very shortly penicillate, sometimes a fruit is developed, seeds not observed but opening between two pistils often present; flowers bisexual, flower pedicel of reduced flowers 1-3 mm long, receptable semiglobose, ovary inferior, calyx lobes 4, broadly triangular, 0.1-0.3 mm long, reddish to dark red, petals 4-5, white, valvate, cucullate, margin membranous, 1.2-2.5 mm long, 1-1.5 mm broad, white; hypanthium 1.8-2 mm diameter, 1.5-1.8 mm high, semiglobose, reddish to dark red, stamens 6-8, shedding immediately after fertilization filaments 1.5-3 mm long, anthers 0.6-1 mm long, 0.5-0.8 mm broad, shedding after fertilization, pistils 2(-3), 0.8-1.5 mm long, enlarging up to 1-1.5(-2.5) mm during fruit maturation, stigmas apically clavate and shortly penicellate, light brown when fertile, changing to dark brown after fertilization; fruit a semiglobose capsule with 8 lateral ribs, apically with a conspicuous border, brownish, 3-4 mm high, 3-4 mm broad above, 2-4 mm diameter, opening between the two pistils to release seeds, seeds reddish, elongated, with pointed apices, 1.1-1.6 mm long, 0.2 mm thick.
Habitat: mainly in temperate forests such as pine forest, pine-oak forest, coniferous forest, and seasonally deciduous forest, rarely montane rainforest with temperate elements, sometimes growing on granitic boulders or vertical rock walls with waterfalls instead of on trees, sometimes in steep riparian canyons, nearly always associated with small water streams resulting in a microhabitat with extremely high and constant relative humidity.
Distribution: Hydrangea seemannii is the only species of Hydrangea section Cornidia which distribution area is restricted to the temperate climate zone of the northern hemisphere. More specifically, it is endemic to the Sierra Madre Occidental in the states of Chihuahua, Durango, Nayarit, Sinaloa and Sonora at elevations between 1050 and 2750 m.
Phenology: Hydrangea seemannii has been collected with buds, flowers or fruits in all months of the year. However, the majority of flowering collections is from June to September.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. seemannii is Endangered, with an AOO of 144 km2, despite an EOO of nearly 55,000 km2, with severaly fragmented and continuously declining populations, as a consequence of habitat destruction.
Additional specimens examined: MEXICO. Chihuahua, municipality Batopilas, Barranca de Batopilas, vicinity of La Bufa, Arroyo Bakosiachi, XI.1973, R. A. Bye 5925 (GH) (bisexual; fruits); side arroyo west of Arroyo La Bufa, on south side of Barranca Batopilas, 1067 m, 13.X.1975, R. A. Bye 7095 (GH) (sterile); Arroyo La Bufa, on S side of Barranca de Batopilas, 1050-1070 m, 29.XII.1976, R. A. Bye 7365 (MEXU) (sterile). Municipality Guazapares, north of Rokoloibo, 1859 m, 5.IX.1973, R. A. Bye 5734 (GH) (fruits); between Wasachic and Rokoloibo, on east side of Barranca de Chinipas, 1829 m, 25.VII.1974, R. A. Bye 6504 (GH) (fruits). Municipality Ocampo, bottom of Arroyo Durazno, ca 3 km upstream from La Bateria and 3.6 mi by road from Pinos Altos, 1730 m, 28°16'54"N, 108°20'4"W, 7.VII.1994, R. S. Felger et al. 94-301 (CAS) (sterile); Arroyo Durazno, 2.5 km above crossing of Pinos Altos - Las Baterías Road, 1730 m, 28°17'N, 108°20'W, 7.VII.1994, M. Fishbein et al. 1828 (MEXU, MO) (fruits); in rocks beside the Río Durazno in Pine-Oak Woodlands (Quercus crassifolia), upriver from the road from Pinos Altos to Bateria, 1850 m, 28°16'N, 108°20'W, 28.VII.1988, P. Jenkins et al. 88-331 (CAS) (fruits); ca. 2 mi W of Pinos Altos on road to Las Baterías, along Río Haciendito, about 1/4 mi upstream from first bridge crossing (tributary to Río Concheño), 1700 m, 28°17'N, 108°19'W, 28.VI.1991, Mahrt et al. 158 (F) (bisexual; fruits); La Batería, Río Haciendito (=Río Durazno) crossing, 1700 m, 28°17.50'N, 108°20'W, 19-20.V.1988, P. S. Martin et al. s.n. (MEXU) (bisexual); 5.25 rd km NW of Pinos Altos on road descending to Las Baterías, at the crossing of the Río Haciendita (a tributary of the Río Cocheño), 1700 m, 28°17'N, 108°19'W, 28.VI.1991, R. Spellenberg and P. Martin 10838 (MEXU, NY) (bisexual; fruits; enlarged marginal flowers absent). Municipality Urique, along Río Urique, NW of Humira bridge, 27°24'12.50"N, 107°32'44.07"W, 1640-1500 m, 5.I.1981, R. A. Bye et al. 10113 (MEXU) (sterile). Durango, municipality Canelas, 7 km from Cañada del Macho on way to Canelas, 2184 m, 23°07'16.9"N, 106°29'36"W, 4.VIII.2009, C. Granados and M. S. Samain 481 (CIIDIR, GENT, MEXU) (fruits); 23 miles before Topia along road from Tepehuanes, 2621 m, 15.I.1976, M. Kimnach and Sánchez-Mejorada 1760 (MEXU, US) (fruits); Paraje “arboles vestidos”, camino Vascajil - Topia, II.1962, Ing. Pimentel B. s.n. (MEXU) (bisexual). Municipality Mezquital, Llano Grandre, aprox. 4 km, 3.VI.1988, I. Solís 1292 (CIIDIR) (bisexual), 1297 (IEB, MEXU) (bisexual); Taxicaringa, camino a 23°14'N, 104°44'W, 2640 m, 25.V.1987, M. González and S. González 2255 (MEXU) (inflorescence buds). Municipality Pueblo Nuevo, Hwy. 40, 37 km from the Sinaloa - Durango border, 2560 m, 28.IX.1974, F. C. Boutin and D. R. Hunt 3907 (MEXU) (fruits); Cascada Mexiquillo, unos 4 km al SW de La Ciudad, 2500 m, 30.VI.1993, M. Cházaro B. et al. 7167 (IEB, MEXU, MICH, XAL) (bisexual); Barranca de Santa Bárbara, stream Santa Bárbara, ejido El Brillante, 15.56 airline km of El Salto, 2685 m, 23°39'38.8"N, 105°26'10"W, 2.VIII.2009, C. Granados et al. 476 (CIIDIR, GENT, MEXU) (sterile), 478 (CIIDIR, GENT, MEXU) (fruits); loc. cit., 11.VIII.2010, C. Granados et al. 499 (CIIDIR, GENT, MEXU) (fruits); loc. cit., 14.VIII.2010, C. Granados et al. 500 (CIIDIR, GENT, MEXU) (fruits); Mexiquillo, El Salto waterfall, 2.33 airline km SE of La Ciudad, 2644 m, 23°42'48.4"N, 105°40'15.5"W, 2.VIII.2009, C. Granados and M. S. Samain 479 (CIIDIR, GENT, MEXU) (fruits); near divide, 36 mi. W of El Salto, km 1119, 24 mi. W of Las Adjuntas, 2743 m, 22.III.1963, E. L. Little, Jr. 18947 (K, US) (sterile); along highway from Durango to Mazatlán, moist ravines about 35 miles west of El Salto, 2400-2500 m, 24.III.1951, R. McVaugh and D. B. Hoover 11536 (MICH) (sterile); steep narrow in precipitous mountainsides along the Mazatlán - Durango highway, 3-15 km toward El Salto from the Sinaloa boundary at El Palmito, 2200 m, 13.IV.1965, R. McVaugh et al. 23606 (MICH) (bisexual); El Salto, waterfall on the headwaters of the Río de la Ciudad, 1.8 miles southeast of La Ciudad, just above the partially constructed, but abandoned, railroad (near 105d 40'W, 23d 41'N), 8150 ft, 26.III.1984, A. C. Sanders et al. 4875 (MO) (sterile); El Salto, Sierra Madre Occidental, rim of plateau W of La Ciudad, 3.1 km NW of Hwy 40 (Durango - Mazatlán) off Neveros rd, upper Arroyo Tascanto watershed, 2720 m, 23°43'46"N, 105°43'40"W, 16.IV.1999, A. C. Sanders et al. 22687 (F, MICH, MO) (bisexual; flowers, fruits); alrededores de El Zapote, aprox. 5 km de Candelarita Chico, 13.V.1988, I. Solís 912 (CIIDIR) (flower buds); El Salto, 2 km al SW de la Ermita, 2100 m, 26.VI.1982, P. Tenorio L. et al. 639 (F, MEXU) (bisexual; fruits); El Salto, El Espinazo del Diablo, carr. Durango - Mazatlán, 2340 m, 23°27' N, 105°48'W, 30.VI.1984, P. Tenorio L. et al. 6099 (MEXU) (fruits); 11.5 mi W of La Ciudad on Rte. 40, 2500 m, 20.VIII.1979, W. L. Wagner and J. C. Solomon 4298 (GH) (fruits). Municipality San Dimas, 2 km de entronque, El Indio, km 180 carretera Durango - Mazatlán, 2280 m, 14.IX.1988, S. González 4257 (CIIDIR, IEB, MEXU) (bisexual; fruits); cascada de arroyo el Madroño, al NE de Vencedores, 24°29'N, 105°48'W, 23.V.1992, S. González 5136 (IEB) (inflorescence buds); Los Chorros waterfall, Ecotourism center “El Madroño”, 2237 m, 24°28'37.7"N, 105°47'28.2"W, 5.VIII.2009, C. Granados and M. S. Samain 483 (CIIDIR, GENT, MEXU) (fruits); El Riito, in ravine near San Luis Villa Corona, 2274 m, 24°06'24.3"N, 105°40'30.9"W, 5.VIII.2009, C. Granados and M. S. Samain 484 (CIIDIR, GENT, MEXU) (juvenile; sterile); San Luis, 51 road miles northwest of Coyotes (about 70 air-line miles west of C. Durango), 2600 m, 1.VII.1950, J. H. Maysilles 7223 (CAS, MEXU, MICH(2), NY) (bisexual); loc. cit., 22.VII.1955, J. H. Maysilles 7931-A (DUKE) (sterile); loc. cit., 22.VII.1955, J. H. Maysilles 7941 (MICH, NY) (bisexual; fruits; enlarged marginal flowers absent); loc. cit., Laguna del Progreso, 34 road miles north of railroad at Coyotes, 2500-2600 m, 1.VII.1955, J. H. Maysilles 8381 (MICH, NY) (bisexual; fruits (MICH); sterile (NY)); loc. cit., 10.VIII.1955, J. H. Maysilles 8387 (MICH) (sterile). Municipality Santiago Papasquiaro, 18 km al W de Ciénega de Nuestra Señora sobre la carretera a Canelas, 2700 m, 28.VIII.1991, J. L. Panero et al. 2249 (MEXU) (fruits); Sierra Madre Occidental, entre Saltares y San Diego, IV.1962, Ing. Pimentel B. s.n. (MEXU) (bisexual; fruits). Municipality Tamazula, ejido El Tecúan, Rancho Agua Azul, 1500 m, 25°34'9"N, 107°0'57"W, 3.IV.2008, F. Mercado et al. 244 (IEB) (juvenile; sterile). Municipality Tepehuanes, El Espíritu, a 2 km de Las Cruces, por el camino de El Huacal a Quebrada Honda, 2260 m, 10.X.1990, M. González 2500 (MEXU) (fruits). Municipality Topia, 0.5 km del Campamento Cuevecillas de la A.U.F. “Topia”, 2650 m, 4.VIII.1990, O. Bravo Bolaños 1159 (CIIDIR, IEB, MEXU) (fruits); Cuevecillas, desviación Canelas - Topia, 2480 m, 25°8'N, 106°24'W, 4.VI.1992, A. García and J. Nocedal 1331 (IEB) (bisexual; fruits); 31.5 km por el camino a Durango, 2530 m, 6.IX.1985, S. González 3466-b (CIIDIR, IEB, MEXU) (bisexual; fruits); Ravine Santa Lucía, 8.41 airline km SE of Topia, 2452 m, 25°10'34.8"N, 106°29'44.8"W, 3.VIII.2009, C. Granados and M. S. Samain 480 (CIIDIR, GENT, MEXU) (fruits); old way to Topia, 1 km from Puerto Cuevecillas, 2617 m, 25°08'53.6"N, 106°24'34.6"W, 4.VIII.2009, C. Granados and M. S. Samain 482 (CIIDIR, GENT, MEXU) (fruits); Topia, 1450-1800 m, 19.IX.1985, P. Tenorio L. et al. 9870 (CIIDIR, IEB, MEXU) (bisexual; fruits). Nayarit, municipality El Nayar, along the Arroyo Santa Rosa W of Santa Teresa 100 airline km NNE of Tepic, 2095 m, 21-24.X.1979, D. E. Breedlove 44610 (MEXU, MO) (fruits); Barranca, 1.5 km al S de Santa Teresa del Nayar, 1900 m, 8.VI.1978, C. L. Díaz Luna 9542 (MICH) (bisexual). Sinaloa, municipality Badiraguato, 5 miles NE of La Cienega along road Santa Rita, 2133 m, 5.XI.1969, D. E. Breedlove and F. S. Kawahara 17040 (CAS (2), MICH) (fruits); 3 miles N of Los Ornos along road to Ocurahui, Sierra Surutato, 6500 ft, 7.III.1971, D. E. Breedlove 19206 (CAS (2), MICH, MO) (bisexual); Sierra Madre Occidental, Sierra Surutato, site 2, Mesa del Frío, north side of Monte Frio, 3.2 miles NNE of town of Surutato (25°51'47"N, 107°32'32.1"W, ± 1000 m), 1939 m, 24.XII.2001, P. Lubinsky et al. 244 (MO) (sterile). Sonora, municipality Yécora, Trigo Moreno (=La Ciénega del Trigo), Sierra El Chuchupate, ca 15 km (by air) southeast of Yécora, 1480 m, 28°17'45"N, 108°47'45"W, 3.VI.1999, A. L. Reina G. et al. 99-201 (MEXU, NY) (bisexual); loc. cit. 1450-1800 m, 17.VIII.1998, T. R. Van Devender et al. 98-1015 (CAS) (bisexual; young fruits).
Observations: Riley (1924) did not include any description of male characters, given that anthers have shed on the type material. The type locality and even its area in the state of Durango are currently unknown.
In the state of Durango, this species together with its host tree is known as “arboles vestidos”, literally traduced as dressed trees, due to the massive white flowering of H. seemannii which covers nearly entirely the trunk and branches of the host tree. Hydrangea seemannii has the broadest altitudinal distribution of all Mexican species.
The stamens of collections Maysilles 7223, Solís 1292 and Reina G. et al. 99-201 have longer filaments than those in the other specimens.
In the following list, we only mention “bisexual” for those specimens where we were able to observe both female and male structures in a single flores. The stamens shed very quickly after fertilization, and as a consequence, when fruits are present, male structures are not available anymore.
Hydrangea sousae Samain, Najarro & E. Martínez, sp. nov. Figs. 1, 10, 12.
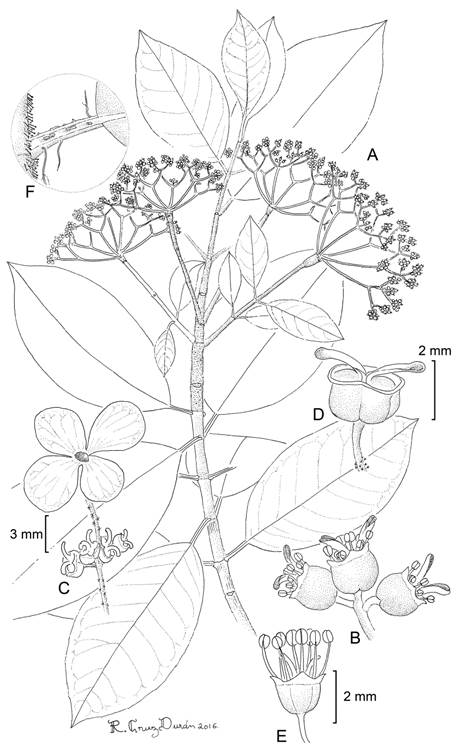
Figure 12: Hydrangea sousae Samain, Najarro & E. Martínez: A. branch with inflorescence; B. functionally female flowers; C. enlarged marginal flower; D. fruit; E. functionally male flower; F. leaf petiole with indument. M. S. Samain et al. 2012-083 (holotype: MEXU). Drawing by Ramiro Cruz Durán.
TYPE: MEXICO. Chiapas, municipality Motozintla, along the road Las Cruces - Emiliano Zapata, Los Alizos, 2180 m, 15°19'24.14"N, 92°16'12.66"W, 5.IX.2012, M. S. Samain et al. 2012-083 (holotype: MEXU!, isotypes: BIGU!, CAS!, CHIP!, GENT!, iBUG!, IEB!, K!, MEXU!, MO!) (functionally female, mature fruits, enlarged flowers).
Functionally dioecious species endemic to southern Mexico which can be distinguished from Hydrangea steyermarkii by the apical shrub-like growth above its host tree canopy, elliptic to lanceolate, coriaceous leaves, strikingly acuminate apex, revolute leaf margin with marginal vein, much less crenate with glands which are much smaller, and 2-3 leaf pairs below the inflorescence which shed less quickly after inflorescence development.
Root-climbing liana of up to 60 m high, apically growing shrub-like above its host tree canopy, functionally dioecious; main stem clinging to the host tree with many adventitious roots with much indument (vs. nearly glabrous in H. steyermarkii), up to 20 cm diameter, bark reddish, with lenticels; runner shoots (stolons) reddish, with adventitious roots and scarce white stellate hairs, with small opposite leaves with dentate margin, also with a few white stellate hairs at both sides; branches terete, notoriously many-ribbed, with many petiole scars, branches attached to host tree with many adventitious roots, densely pubescent, indument deciduous, resulting in glabrous older branches, apical zone densely pubescent with stellate brown-reddish hairs, petioles slightly clasping the stem and forming a continuous ring connecting both petioles; juvenile branches with less indument than H. steyermarkii; leaves decussate, petiole slightly to profoundly sulcate adaxially, glands present near the border of the sulcus, terete abaxially, colour brownish grey, sometimes broader at the basis, adaxially glabrous, abaxially with reddish-brown to whitish penicellate hairs, when mature glabrous, with small adventitious roots, 1-3 cm long, corchy in older leaves, leaving a triangular scar on the branch when leaves fall (scar sometimes with slightly rounded borders); lamina flat, elliptic to lanceolate, 9-18 cm long, 4.5-8 cm broad, base rounded, cuneate to decurrent, sometimes slightly asymmetric, apex strikingly acuminate, leaf margin revolute with marginal vein, slightly crenate with striking glands (smaller than in H. steyermarkii), especially in young leaves and in the upper leaf portion, sometimes with small teeth, venation brochidodromous, especially notorious in the upper leaf part, veins 6-10 pairs, adaxial leaf side with sunken (in this case sulcate), marked or protruding midvein, primary and secondary veins only lightly marked (more so in younger leaves), primary veins sometimes sunken, tertiary veins sometimes not visible, areoles formed by secondary veins smaller than in H. steyermarkii (resulting in a higher number of areoles between the primary veins than in H. steyermarkii), abaxial side with marked midvein and primary veins and slightly sunken, smooth or marked secondary veins, areoles well-marked, glabrous or with relatively little white stellate indument on both adaxial and abaxial sides, abaxial side also with reddish indument, especially on the secondary veins, opaque green, young leaves sparsely to densely pubescent with stellate indument, adaxial side with both whitish and reddish hairs (generally more whitish), abaxial side with reddish hairs on the veins and whitish hairs on the lamina or yellowish-reddish hairs allover, indument of leaves on young branches stellate and whitish, whereas reddish on leaves in meristematic zones, acarodomatia consisting of (often broad) cavities with indument inside, rarely glabrous, present in axils between midvein and primary veins, as well as in axils between primary and secondary veins; inflorescence axis densely pubescent with whitish or reddish, stellate hairs, (2.5-)5-9 cm long, with 2-3 opposite or decussate leaf pairs below the inflorescence, deciduous, petiole 2-4 mm long, 1 mm broad, lamina 2-5 cm long, 0.7-2 cm broad, these leaves within one specimen of a similar size, apex of the floral axis woody, triangular to cone-shaped, with dense reddish-brown pubescence, elongated bract scars visible, thickening at the top, 3-8 mm broad, 3-7 mm high (up to 1 cm high in old lignified inflorescence axes), inflorescence bracts cucullate, pale to reddish brown, coriaceous, border membranous, adaxially and abaxially with scarce whitish stellate hairs, especially the outer ones and near the base, veins slightly darker, bracts increasing in size, lowermost bract 1.2 cm large, 1.4 cm broad, higher bracts up to 3 cm large, 2.5 cm broad, consecutively and rapidly deciduous during inflorescence development, bracts of partial inflorescences linear, reddish, up to 2.5 cm long, 1 mm broad at the base, gradually tapering towards a pointed apex, deciduous, inflorescences lateral, opposite, 1-3 pairs of inflorescences per flowering branch, sometimes one inflorescence developing only, flowering branch only continues growing vegetatively relatively quickly during inflorescence development, with maximum eight leaf pairs above the inflorescences, rarely some small kataphylls as well, inflorescence axes with basal lignified parts of inflorescences of previous years visible in well-collected specimens, allowing to observe growth and flower periodicity, these rests 15-17 cm apart, with up to 10 decussate leaf pairs (or the scars of these leaves, this number constant within one specimen) in between, medulla central in the branch, disappearing in older branches, leaving a hole only, leaves at the base of the inflorescence generally absent, inflorescences umbellate, opposite, buds up to 4 cm broad and 3 cm high before opening, in flowering stage 8-13 cm diameter, 3.5-8 cm high, with 5-6 main axes in both sexes, but robust in functionally female plants, and delicate in functionally male plants, partial inflorescences cymes (when enlarged marginal flowers are present, these terminally placed in each cyme), secondary and tertiary inflorescence axes with reddish-brown stellate hairs; enlarged marginal flowers rarely present (only one observed), flower pedicel 0.6 mm long, receptacle rectangular, with 4 lateral ribs, 1 mm high, 1 mm broad, 1 mm diameter, ovary inferior, sepals 4, white, coriaceous with marked veins, spathulate to broadly elliptic, 3-5 mm long, 2.5-4.5 mm broad, petals reduced and deciduous, stamens and stigmas not differentiated; flower pedicel of reduced flowers (0-)0.2-2 mm long in functionally male flowers, 1-2 mm long in functionally female flowers, receptacle broadly campanulate in functionally male flowers, semiglobose in functionally female flowers, ovary inferior, calyx lobes 4, triangular, sometimes nearly spathulate, breaking or drying when older, in the latter case revolute, 0.3-0.8 mm long, petals 4, white, valvate, cucullate, margin revolute, membranous, 1.9-2.8 mm long, 1.5-1.8 mm broad, white; functionally male flowers: hypanthium 1.5-2 mm diameter, stamens 6-7, well-developed, filaments 1-1.2(1.5) mm long, anthers 0.8-1 mm long, 0.4-0.8 mm broad, pistils 2, reduced, 1-1.2 mm long, stigmas not penicellate; functionally female flowers: hypanthium 1.5-2 × 2.5 mm, stamens (4-)5-8, reduced, filaments 0.1-0.4 mm long, anthers 0.3-0.5 mm long, 0.3 mm broad, pistils 2, 1.2-1.6 mm long, enlarging up to 2 mm during fruit maturation, stigmas apically clavate and shortly penicellate; fruit a semiglobose capsule with, often with a notorious constriction in between the pistils, apically with a conspicuous border, brownish, 1.5-2 mm high, mature fruits not seen, opening between the two pistils to release seeds, seeds not seen.
Habitat: northwards exposition in conserved cloud forest, sometimes dominated by Quercus L. with trees up to 70 m high and diameters of more than 1 m, with many epifytes as well as large lianas and dense undergrowth. All known localities are characterised by a permanent high humidity and high nubosity, on very steep slopes with metamorphic rocks, often near the top of the mountain (range). The population of the Tacaná Volcano grows in secondary forest (“acahual”) of 50 years old, owned by Benjamin Zacarías, with a cloud forest composition with amongst others Hedyosmum mexicanum and Lauraceae, trees of 25-30 m high and diameters of 50-70 cm, with few epiphytes, small lianas and without undergrowth of shrubs and herbs. This is an extraordinary example of long term succession where the tree composition is changing from cloud forest to oak forest.
Distribution: this species is endemic to Mexico known from conserved northwards-oriented slopes in the Sierra Madre de Chiapas between Motozintla and Niquivil, between 2175 and 2540 m a.s.l., and the northwestern slopes of the Tacaná Volcano, between 1960 and 2056 m, on metamorphic and igneous volcanic rocks.
Phenology: Hydrangea sousae has been collected in flower in the months of September and January.
Etymology: this species is dedicated to Dr. Mario Sousa Sánchez (1940-2017), to honour his extraordinary vision and dedication to establish the National Herbarium of Mexico.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. sousae is Endangered, with an EOO of slightly over 121 km2, severely fragmented and continuously declining populations, as a consequence of habitat destruction, increasing human population and fires. A volcanic eruption may be an additional threat for this species.
Additional specimens examined: MEXICO. Chiapas, municipality Cacahoatán, Volcán Tacaná, 2.2 km in straight line from Agua Caliente, 1960 m, 15°08'53.0"N, 92°08'38.7"W, 27.I.2011, M. S. Samain et al. 2011-007 (CHIP, GENT, IEB, MEXU) (juvenile; sterile); loc. cit., 2.44 km in straight line from Agua Caliente, 1960 m, 15°08'49.2"N, 92°08'33.2"W, 27.I.2011, M. S. Samain et al. 2011-008 (CHIP, GENT, IEB, MEXU) (juvenile; sterile); loc. cit., 2.4 km in straight line from Agua Caliente, 2056 m, 15°8'48.33"N, 92°8'36.77"W, 6.I.2012, M. S. Samain et al. 2012-013 (CHIP, GENT, K, IEB, MEXU) (functionally male). Municipality Motozintla, 45-50 km northeast of Huixtla along road to Motozintla, 1900 m, 28.XII.1972 D. E. Breedlove and R. F. Thorne 31029 (MEXU) (sterile); along the road Niquivil - Motozintla, in front of the entrance to the road to Justo Sierra, 2492 m, 15°18'15.20"N, 92°14'12.02"W, 7.I.2012, M. S. Samain et al. 2012-014 (CAS, CHIP, GENT, IEB, MEXU(2), MO) (functionally male); loc. cit., 2467 m, 15°19'7.91"N, 92°15'28.47"W, 7.I.2012, M. S. Samain et al. 2012-015 (GENT, IEB, MEXU) (sterile); along the road Las Cruces - Emiliano Zapata, 2537 m, 15°18'32.43"N, 92°14'54.23"W, 8.I.2012, M. S. Samain et al. 2012-016 (CHIP, GENT, IEB, MEXU) (inflorescence buds); along the road Las Cruces - Emiliano Zapata, 2537 m, 15°18'32.43"N, 92°14'54.23"W, 8.I.2012, M. S. Samain et al. 2012-017 (CAS, CHIP, F, GENT, iBUG, IEB, K, MEXU(2), MO, US, XAL) (functionally male); carretera Niquivil - Motozintla, crucero a San Lucas, 2516 m, 15°16'24.23"N, 92°13'59.58"W, 5.IX.2012, M. S. Samain et al. 2012-081 (CHIP, GENT, MEXU) (sterile); along the road Las Cruces - Emiliano Zapata, Los Alizos, 2472 m, 15°19'7.69"N, 92°15'24.94"W, 5.IX.2012, M. S. Samain et al. 2012-082 (CHIP, GENT, IEB, MEXU) (functionally female); loc. cit., 2175 m, 15°19'24.23"N, 92°16'13.07"W, 5.IX.2012, M. S. Samain et al. 2012-084 (CHIP, GENT, IEB, MEXU) (functionally male). Municipality Siltepec, on the ridge above Siltepec on the road to Huixtla, 2000-2400 m, 1.II.1982 D. E. Breedlove and F. Almeda 58251 (MEXU) (functionally female).
Observations: this species had only twice been collected before we made the type collection.
Hydrangea steyermarkii Standl., Publ. Field Mus. Nat. Hist., Bot. Ser. 22(4): 233. 1940. Fig. 4.
See Samain et al. (2014) for an amended description and notes on habitat, distribution, phenology, additional observations, conservation status as well as a list of all known collections.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. steyermarkii is Endangered, with an Area of Occupancy (AOO) of slightly over 32 km2, severely fragmented and continuously declining populations, as a consequence of habitat destruction, increasing human population and fires. A volcanic eruption may be an additional threat for this species. Morever, no recent collections from Guatemala are known, the last herbarium specimens being collected by Julian Steyermark in the 1940s.
Hydrangea tapalapensis Samain, Najarro & E. Martínez, sp. nov. Figs. 4, 13, 14.
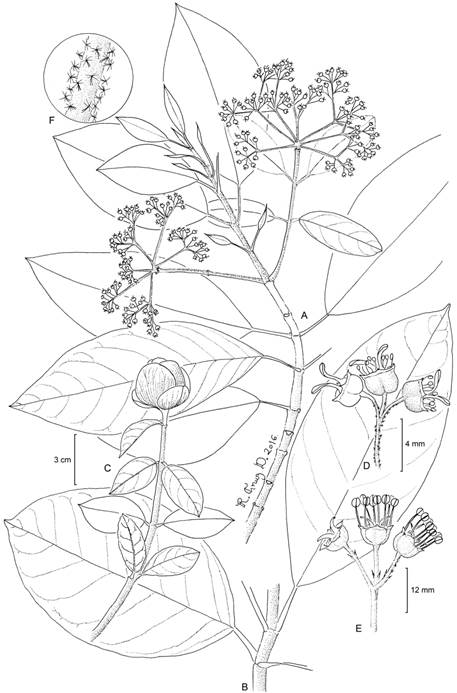
Figure 13: Hydrangea tapalapensis Samain, Najarro & E. Martínez: A. branch with functionally female flowers; B. adult leaves of a specimen with particularly large leaves; C. branch with inflorescence bud; D. functionally female flowers; E. functionally male flowers, F. indument of inflorescence axis. A, F, M. S. Samain et al. 2012-007 (holotype: MEXU); B, M. S. Samain et al. 2011-005 (MEXU); C, M. S. Samain and E. M. Martínez Salas 2012-091 (MEXU); D, M. S. Samain et al. 2012-076 (MEXU); E, M. S. Samain et al. 2017-003 (MEXU). Drawing by Ramiro Cruz Durán.

Figure 14: Hydrangea tapalapensis Samain, Najarro & E. Martínez. A. habit, the arrow indicates a branch with lignified old inflorescence axes growing above its host tree canopy; B. vegetative branch with pubescence; C. young inflorescence with inflorescence bracts and flower buds; D. young inflorescence with flower buds, showing the pinkish petals. E. female inflorescence; F. female inflorescence with maturing fruits. Photos by Marie-Stéphanie Samain and Esteban Manuel Martínez Salas.
TYPE: MEXICO. Chiapas, municipality Tapalapa, Reserva Municipal, 6.1 km before Tapalapa (distance measured along the road), 62 m from the roadside, 2142 m, 17°11'17.07"N, 93°7'22.96"W, 3.I.2012, M. S. Samain et al. 2012-007 (holotype: MEXU!, isotypes: CAS!, CHIP!, F!, GENT!, GH!, iBUG!, IEB(3)!, K!, MEXU(3)!, MO!, NY!, US!, XAL!) (functionally female).
Functionally dioecious species endemic to southern Mexico which can be distinguished from Hydrangea steyermarkii by the apical shrub-like growth above its host tree canopy, leaf margin slightly undulated to lobed or dentate with glands which are much smaller, 3-5 decussate leaf or kataphyll pairs below the inflorescence, petals white with reddish lines or pinkish.
Root-climbing liana of up to 45 m high, apically growing shrub-like above its host tree canopy, functionally dioecious; main stem clinging to the host tree with many adventitious roots, up to 20 cm diameter, bark dark brown, with lenticels; runner shoots (stolons) reddish, with red adventitious roots and white stellate hairs; branches many-ribbed, slightly angular, with some corchy zones, with many leaf scars, with reddish stellate hairs, branches attached to host tree with adventitious roots, these roots whitish with reddish-brown corchy base, free-growing branches slightly angular, slightly fissured and exfoliant, pubescent with reddish penicellate pairs; leaves decussate, petiole sulcate adaxially, except sometimes for its basis which is usually broadened, terete and sometimes corchy abaxially, glabrous to pubescent reddish penicellate hairs, when mature (nearly) glabrous, 1-3 cm long, leaving a triangular scar with rounded borders on the branch when leaves fall; lamina flat, elliptic-obovoid, rarely slightly orbicular or lanceolate, in this case sometimes with 3 large apical lobes, 6-20(-28) cm long, 3-8(-11) cm broad, base cuneate to rounded, apex slightly acuminate, acute or rounded, leaf margin slightly undulated to lobed or dentate with glands, marginal vein present, margin sometimes revolute, venation brochidodromous, veins 6-9 pairs, adaxial leaf side with midvein sunken in its lower half, even in upper half, primary and secondary veins slightly sunken, marked to protruding (this latter character especially in larger leaves), primary veins join to form submarginal or intramarginal vein, glabrous, abaxially with protruding midvein and primary veins, marked secondary veins, areoles well-marked, glabrous to sparsely whitish pubescent in mature leaves and even less hairy on primary and secondary veins, opaque olive green to brownish green, midvein brownish to reddish, young leaves pubescent, brownish with paler brown crystals on the surface, with deciduous white stellate hairs in meristematic zones, base of stellate hairs on adaxial side shorter than that of hairs on abaxial side, acarodomatia small to relatively large membranous cavities, glabrous or with few white hairs, present in the axils between midvein and primary veins, and in the lower leaf portion in the axils between primary and secondary veins as well; inflorescence axis densely pubescent with adnate, whitish to reddish, stellate hairs, 5-20 cm long, with 2-3 opposite or decussate leaf pairs below the inflorescence, deciduous, petiole 0.3-1 cm long, 1 mm broad, lamina 1-3 cm long, 0.4-2 cm broad, apex of the floral axis woody, cone-shaped, elongated bract scars visible, thickening at the top, 3-4 mm broad, 3-4 mm high, inflorescence bracts cucullate, pale to reddish brown, membranous, adaxially and abaxially nearly glabrous to densely pubescent with whitish stellate hairs, veins darker, bracts increasing in size, lowermost bract 1 cm large, 1 cm broad, higher bracts up to 2.5 cm large, 2 cm broad, consecutively and rapidly deciduous during inflorescence development, bracts of partial inflorescences linear, pale brown, up to 2.5 cm long, 4 mm broad at the base, gradually broadening towards a pointed apex (sometimes with 2 apices), deciduous, inflorescences lateral, opposite, 1-3 pairs of inflorescences per flowering branch, sometimes one inflorescence developing only, flowering branch continues growing vegetatively quickly during inflorescence development, with leaf and/or kataphyll pairs, inflorescence axes with basal lignified parts (or even entire inflorescence axes) of inflorescences of previous years visible in robust well-collected specimens, allowing to observe growth and flower periodicity, these rests 8-15 cm apart (but relatively constant within one specimen), with 3-5 decussate leaf or kataphyll pairs (or scars, this number constant within one specimen), deciduous, medulla central in the branch, disappearing in older branches, leaving a hole only, leaves at the base of the inflorescence present, inflorescence umbellate, buds up to 3 cm broad and 2 cm high before opening, in flowering stage 6-10 cm diameter, 3-6 cm high, with 5(-)7-9 main axes in functionally male plants, 6-9 main axes in functionally female plants, partial inflorescences corymbs, secondary and tertiary inflorescence axes with reddish stellate hairs, especially adaxially; enlarged marginal flowers always absent; flower pedicel of reduced flowers 1-4 mm long in functionally male flowers, 1-4 mm long in functionally female flowers, receptacle campanulate in functionally male flowers, semiglobose in functionally female flowers, sometimes slightly narrower near its margin, ovary inferior, calyx lobes 4, triangular, 0.3-0.8 mm long, petals 4, white with reddish lines or pinkish, valvate, membranous, 1.1-2.6 mm long, 0.7-1.2 mm broad, white with reddish lines or pinkish; functionally male flowers: hypanthium 2-2.5 mm diameter to completely round, stamens 6, well-developed, filaments 2.5-4 mm long, anthers 0.7-1 mm long, 0.3-0.5 mm broad, pistils 2, reduced, 0.5-0.6 mm long, stigmas not penicellate; functionally female flowers: stamens 6-9, reduced, filaments 0.4-0.8 mm long, anthers 0.3-0.6 mm long, 0.2 mm broad, pistils 2-3, 1.5-2.8 mm long, enlarging up to slightly over 3 mm during fruit maturation, stigmas apically clavate and shortly penicellate; fruit a semiglobose capsule with 4 slightly protruding lateral ribs, sometimes two of them forming a constriction in between the pistils, apically with a conspicuous slightly paler border, dark reddish brown, 1.5-1.6 mm high, 2.3-3.2 mm broad above, 0.7-1.2 mm diameter or rectangular, opening between the two pistils to release seeds; seeds not seen.
Habitat: very dense cloud forest, in localities with a very high rainfall and cloudiness as well as constantly high relative humidity, on north-facing very steep slopes, or near the ridge of north-facing very steep slopes, exposed directly to the humid northern winds. In well-developed soil on metamorphic rocks, between 1990 and 2180 m a.s.l.
Distribution: endemic to the municipality of Tapalapa, Chiapas, in the municipal reservate on the so-called Cerro de las Antenas as well as on the highest parts in the northern portion of the Cerro La Bandera. The locality of the collection realized by Arturo Gómez-Pompa in the neighbouring municipality of Pueblo Nuevo Solistahuacán has been destroyed.
Phenology: Hydrangea tapalapensis has been collected in flower and fruit in January and September.
Etymology: dedicated to the community of Tapalapa who decided to conserve this area of which we sincerely hope they will continue to do so.
Conservation status: following the IUCN Red List categories and criteria (IUCN, 2012), H. tapalapensis is Critically Endangered, with an EOO of slightly less than 39 km2, severely fragmented continuously declining populations as logging is continued throughout the area.
Additional specimens examined: MEXICO. Chiapas, municipality Pueblo Nuevo Solistahuacán, al NE de Pueblo Nuevo Solistahuacán, 2050 m, 11.VIII.1967, A. Gómez-Pompa 2509 (MEXU (3)) (sterile). Municality Tapalapa, Reserva Municipal, 6.1 km before Tapalapa (distance measured along the road), 340 m from the roadside, 1989 m, 17°11'25.0"N, 93°07'25.4"W, 25.I.2011, M. S. Samain et al. 2011-005 (CAS, GENT, IEB(2), MEXU(3), MO) (functionally female; mature fruits); loc. cit., 40 m from the roadside, 2194 m, 17°11'14.86"N, 93°7'22.94"W, 19.VIII.2011, M. S. Samain et al. 2011-197 (CAS, CHIP, F, GENT, GH, iBUG, IEB(2), K, MEXU(2), MO, XAL) (functionally female; young fruits); loc. cit., 2112 m, 17°11'46.37"N, 93° 7'6.08"W, 31.VIII.2012, M. S. Samain et al. 2012-073 (CAS, CHIP, F, iBUG, IEB(2), GENT, MEXU(2), MO, XAL) (functionally female), 2012-074 (BR, CAS, CHIP, CR, GENT, IBUG, IEB, K, MEXU(2), MO, QMEX, US, XAL) (functionally female); loc. cit., 2161 m, 17°11'45.43"N, 93°7'4.52"W, 31.VIII.2012, M. S. Samain et al. 2012-075 (CHIP, IEB, GENT, MEXU) (functionally female); loc. cit., 2252 m, 17°11'14.56"N, 93° 7'22.11"W, 31.VIII.2012, M. S. Samain et al. 2012-076 (CAS, CHIP, F, GENT, IBUG, IEB(2), MEXU(3), MO, QMEX, XAL) (functionally female); loc. cit., 2254 m, 17°11'15.82"N, 93°7'22.49"W, 31.VIII.2012, M. S. Samain et al. 2012-077 (CHIP, GENT, MEXU) (functionally female; fruits); Cerro de la Bandera, 2177 m, 17°12'42.62"N, 93°5'11.44"W, 19.VIII.2011, M. S. Samain et al. 2011-198 (GENT, MEXU(2)) (juvenile); loc. cit., 2169 m, 17°12'42.90"N, 93°5'11.90"W, 19.VIII.2011, M. S. Samain et al. 2011-199 (BIGU, GENT, iBUG, IEB(2), MEXU(2), MO) (functionally female; fruits); loc. cit., 2155 m, 17°12'44.46"N, 93°5'12.36"W, 19.VIII.2011, M. S. Samain and E. M. Martínez Salas 2011-200 (CAS, CHIP, DUKE, F, GENT, iBUG, IEB(3), K, MEXU(2), MO, NY, XAL) (functionally female); Cerro de las Antenas, 2257 m, 17°11'15.71"N, 93°7'19.00"W, 10.II.2017, M. S. Samain et al. 2017-003 (F, iBUG, IEB(2), MEXU(2), MO) (functionally male).
Specimen of a putative hybrid between H. albostellata and H. tapalapensis: MEXICO. Chiapas, Tapalapa, camino Coapilla - Ocotepec, 1200 m desde la curva con el cruce del Río Quachi, 1710 m, 17°11'42,98"N, 93°08'39,7"W, 9.IX.2012, M. S. Samain and E. M. Martínez Salas 2012-091 (CHIP, GENT, iBUG, IEB(2), MEXU(2), MO, NY, QMEX, XAL) (functionally male).
Observations: the leaf size may be variable. The specimen Samain et al. 2011-005 has large leaves of 18-28 cm long and 10-11 cm broad, whereas the type collection has much smaller leaves of 15-17 × 5-7 cm. Hydrangea tapalapensis is one of the five Mexican species which may both reach and flower above the tree canopy, together with H. carroniae, H. nahaensis, H. seemannii and H. sousae.
In the population of Cerro la Bandera in the municipality of Tapalapa, there is a clear gradient in amount of inflorescences depending on the humidity: the higher the relative humidity, the more massively the individuals flower.
Hydrangea tapalapensis can be distinguished from the morphologically slightly similar Hydrangea breedlovei by the following characters: apical shrub-like growth above its host tree canopy vs. the absence of such terminal growth, elliptic-orbicular vs. narrowly lanceolate to elliptic leaves, leaf margin slightly undulated to lobed or dentate with glands vs. entire margin, pinkish vs. white petals, and the filaments in the male flowers of H. tapalapensis measuring only half of those of H. breedlovei. Additionally, these species grow on different geological rocks: volcanic vs. limestone rocks.
Hydrangea albostellata, the most common Hydrangea species in southeastern Mexico which we have recently described (Samain et al., 2014), also occurs in Tapalapa, but can be easily distinguished from H. tapalapensis on the basis of its pubescent leaves and very large fruits. In fact, we observed and collected material of a putative hybrid individual with intermediate characters between both species; most of its morphological characters clearly distinguish it as H. tapalapensis, but its indument is closer to that of H. albostellata. The data of this specimen are included after the data of the specimens of H. tapalapensis.
Discussion
Dubious species circumscriptions and extensive synonymization in Hydrangea section Cornidia
Since the revision by McClintock (1957), species circumscriptions in Cornidia are very dubious, lumping very distinct entities in one species, e.g. Hydrangea steyermarkii, the name of a rare species, currently known from only one population with adult individuals on the border of Mexico and Guatemala, applied to up to five distinct species in Mexico and Central America (own observations on herbarium specimen annotations; Christenhusz, 2009), such as H. albostellata and several here newly described species, or H. peruviana Mor., another rare species from the lower western Andes in Ecuador, widely used to name up to seven or eight morphologically completely different species in Mexico (H. seemannii), Central and South America (own observations on herbarium specimen annotations; McClintock, 1957; Freire-Fierro, 2004; Christenhusz, 2009). This situation seems to be due to the fact that most studies since McClintock (1957) were largely or even entirely based on herbarium specimens and additionally, type specimens were probably not always observed, as was assumed that McClintock had done so.
Diversity of some morphological and architectural characters of Hydrangea section Cornidia
This study shows that the diversity of morphological and architectural characters within the largely Neotropical section Cornidia has been underestimated, overlooked and/or misunderstood. Although many characters remain visible on herbarium specimens, our field work has made clear that observation of whole individuals in the wild is essential to obtain a good insight in intra- and interspecific variation within this group. It is obvious that this is mainly due to the impressive size of most individuals, which makes that any herbarium specimen is only partially representative.
Undoubtedly the most controversial character in this group is the size of the stamens versus the size of the pistils in particular species, or even the absence or quick shedding of stamens, which has led to the hypothesis that flowers are often unisexual and that species are dioecious (e.g., McClintock, 1957; Nevling and Gómez-Pompa, 1968; Christenhusz, 2009), whereas we observed that the majority of the species are functionally dioecious and generally have both female and male structures, though one of them is reduced.
A second controversial observation in previous studies and on herbarium specimen labels is the architecture of the Neotropical Hydrangeas, which is often referred to as shrubby or epiphytic (e.g., Christenhusz, 2009). However, our field observations without any exception show root climbing and lianous individuals rooting near the base of their host trees. Nevertheless, in some cases the apical portion of the plant, protruding above the canopy of its host tree, or rarely above the rock wall on which the plant grows, does show affinities to a “shrubby“ growth form (Granados Mendoza et al., 2014).
Distribution of Hydrangea in Mexico and affinities with Central and South America
Species of the genus Hydrangea occur in all large mountain systems of Mexico, with the exception of the Sierra Madre del Sur, as well as in some isolated mountain ranges in the coastal plain of the Gulf of Mexico, between 830 and 3000 m a.s.l., with the majority of the individuals between 1000 and 2000 m elevation. The well-known species Hydrangea seemannii has the widest distribution area of all Mexican species and occurs in the Sierra Madre Occidental. This species is also the northernmost representative of Hydrangea section Cornidia in America, occurring up to 28°17'N latitude.
The genus is especially abundant across the Sierra Madre Oriental, although further exploration lacks in the state of Oaxaca, and in parts of the states of Puebla and Veracruz. Two species, H. carroniae and H. nebulicola occur in zones which belong to the geological province of the Transmexican Volcanic Belt, although these specific areas are floristically more closely related to the Sierra Madre Oriental than to the Transmexican Volcanic Belt. Finally, the genus is also represented by several species in the extreme east of the Sierra Madre de Chiapas, as well as on the Volcano Tacaná on the border with Guatemala which belongs to the Central American volcanic system. Hydrangea tapalapensis occurs in mountains of volcanic origin which form a connection between de Transmexican Volcanic Belt and the Central American volcanic system.
The majority of the species show a very high degree of endemism, restricted to a relatively small latitudinal and altitudinal zone. In some species, this endemism is at least partially the result of climatological (e.g., H. breedlovei) or geographical (H. otontepecensis) isolation from surrounding areas. However, in most species, the currently observed distribution probably is a relict of a once wider distribution area. This is not only confirmed by the fact that several localities where Hydrangea previously had been collected do not exist anymore, but also by preliminary ecological niche modelling analyses (unpublished data). These analyses show that all Mexican Hydrangea species occur in very specific microniches in temperate to cold climates with a high relative humidity. The majority of the species grow on slopes exposed to the north.
Observations during extensive recent field work in Central and South America show that, with the exception of H. steyermarkii which occurs on the border of Mexico and Guatemala, only one Hydrangea species (H. albostellata) is shared between Mexico and Central America and that Mexico does not share any species with South America, where the genus is especially diverse. Therefore, Mexico is one of the countries with the highest proportions of endemic Hydrangea species and could be considered as a secondary hotspot for the section Cornidia.
Conservation of Hydrangea in Mexico
Following the IUCN Red List categories and criteria (IUCN, 2012), all ten currently known Mexican Hydrangea species are threatened. Seven species are Endangered, and three are Critically Endangered. The three latter are all new to science, H. nahaensis, H. otontepecensis and H. tapalapensis and are endemic to a very small area despite extensive explorations in a large zone; therefore, they have been named after the village that is doing efforts to conserve them. The most direct causes of their threatened status are deforestation and land use changes. However, the most dramatic cause that might bring several species to extinction in the near future is the current and future predicted climate change which is driving cloud forest species in eastern and southern Mexico towards upward migration to higher elevations (e.g., Rojas-Soto et al., 2012). As many Mexican Hydrangea species are restricted to the higher or even highest elevations in their respective distribution areas in cloud forest habitat, their future does not look very bright.
As mentioned above, preliminary work by Granados Mendoza et al. (2010) shows that even the most widespread Mexican Hydrangea, H. seemannii, might be on the brink of extinction by 2080. This especially may happen if no immediate action is undertaken to protect it from illegal logging of its host trees, as well as ensuring that new or extended conservation areas include populations of this highly attractive species, as currently, nearly none of them grows within one of the Mexican federal natural protected areas which are present in their distribution area.
Moreoever, their preference to grow mostly in primary forest with very specific conditions (often near water, flat topography, efficient drainage) makes them promising bio-indicators. Interestingly, in very humid areas, such as the cloud forest on the Tacaná Volcano, they grow in secondary forest where the succession process has nearly reached its climax. Additionally, they also occur on relict trees of primary forest which are surrounded by near-climax secondary forest.
On several locations throughout Mexico, Hydrangea macrophylla, a shrubby hortensia from Asia, is cultivated or naturalized in or near native Hydrangea populations, as it apparently prefers a similar habitat and ecological conditions. As mentioned by Samain et al. (2014), at least in the case of H. steyermarkii, the quite ironic land use change of a native Hydrangea to an alien one is severely endangering the local species. It is currently unknown what the ecological impact of this might be on the longer term.
A final fact is that, to date, we are not able to distinguish between individuals and populations in most localities, unless plants of different sexes are present. This is due to the fact that what appear to be distinct plants growing on their respective host trees are interconnected by long stolons which may not be visible at first glance, as they are at least partially covered by leaf litter on the forest floor. Especially in primary habitat of a larger periphery, it is quite common that many trees carry a climbing Hydrangea, but this may concern a clonal colony rather than distinct indivuals. We have very rarely observed seedlings or juveniles in the wild, indicating that reproduction may be mostly or in some cases even entirely vegetative by stolons, even though sexual reproduction might still take place. Additional field observations and molecular studies are required to elucidate this situation.
Concluding, our recent and extensive field work across Mexico has reveiled an astonishing number of seven new endemic Hydrangea to science, six described here and one by Samain et al. (2014). One species had remained unnoticed, despite its size, and six were incorrectly identified. Of these six, three had only been collected once or twice before. Together with the two previously known species known from this country and the new record, all can truely be considered as “Lonesome Georges” among plants.

