











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El género Malpighia L. (Malpighiaceae) comprende ca. 50 especies de árboles y arbustos nativos de América tropical y subtropical (Davis y Anderson, 2010). En México, se distribuyen 19 especies, 12 de ellas son endémicas (Anderson, 2013; Villaseñor, 2016). Entre éstas se encuentra Malpighia mexicana A. Juss., comúnmente conocida como nanche rojo, guejocote o guachocote (Maldonado-Peralta et al., 2016). Esta especie es una fuente de alimentación para los pobladores locales, por lo que se cultiva en los huertos familiares de zonas rurales, aunque sin ningún manejo agronómico (Morton, 1987; Castañeda-Vildózola et al., 2014). Así mismo, M. mexicana es fuente de recursos económicos, maderables, ornamentales y medicinales (Maldonado-Peralta et al., 2016). Cabe señalar que como especie arbórea brinda protección y sombra a otras especies, tanto vegetales como animales, desempeñando un papel importante dentro del ecosistema como productor de oxígeno (Arambarri et al., 2011). Además, es una fuente de fijación de bióxido de carbono y, por ende, es parte de los reguladores del clima (Villaseñor, 2010).
Dado que el ambiente es un factor que puede influir en las características anatómicas de las especies vegetales, se han realizado diversos estudios enfocados en las hojas (Fank-de-Carvalho et al., 2010; Zamora-Natera y Terrazas, 2012; Jiménez-Noriega et al., 2015), tallos (Rebollar et al., 1994; Herrera-Cárdenas et al., 2000) y raíces (Laskowski y Bautista, 1999; Espinoza-Velázquez et al., 2012; Martín-Montiel et al., 2012). Independientemente del órgano estudiado el análisis indica que las especies tienen estrategias de adaptación a nivel morfológico, fisiológico y anatómico.
Uno de los órganos vegetales más importantes es la hoja ya que en ella se efectúan funciones vitales para la planta; por ello, se ha estudiado la estructura foliar con enfoques taxonómicos y ecológicos. Al respecto, Milan et al. (2006) compararon la anatomía de la hoja de especies de la familia Asteraceae. Todas ellas mostraron diferencias; por ejemplo, Mikania glomerata Spreng. presentó estomas en ambas superficies, tricomas glandulares uniseriados y conductos secretores; por su parte, Porophyllum ruderale (Jacq.) Cass. tiene hidátodos y cavidades secretoras; mientras que Vernonia condensata Baker mostró idioblastos y tricomas glandulares uni y biseriados. Cabe señalar que las estructuras secretoras son caracteres con relevancia taxonómica para esta familia (Castro et al., 1997).
Para el caso de la familia Leguminosae se ha descrito la anatomía foliar de cuatro especies de Lupinus L. (L. aschenbornii S. Schauer, L. exaltatus Zucc., L. montanus Kunth y L. reflexus Rose), las cuales comparten epidermis papilosa con paredes anticlinales, estomas anomocíticos, tricomas simples y mesófilo bifacial (Zamora-Natera y Terrazas, 2012). Se observó que estas especies difieren en la disposición, tamaño y número de haces vasculares, los cuales permiten su identificación. Por ejemplo, Lupinus exaltatus tiene haces dispuestos en un cilindro vascular continuo y compacto, mientras que en L. aschenbornii forman un anillo y en L. reflexus y L. montanus se presentan de forma discontinua (Zamora-Natera y Terrazas, 2012).
Otra familia estudiada es Solanaceae; Liscovsky y Cosa (2005) describen la anatomía foliar de 13 especies de los géneros Cestrum L. y Sessea Ruiz & Pav., donde reportan que el tipo de estomas y tricomas, así como la arquitectura de la vena secundaria, son los caracteres anatómicos más importantes en la taxonomía de estos géneros. Además, observaron que Cestrum se caracteriza por tener una venación broquidódroma; en cambio en Sessea es semicraspedódroma.
En la familia Malpighiaceae, mediante la descripción de la anatomía de la hoja, se ha reportado la presencia de laticíferos en los géneros Galphimia Cav. y Verrucularia A. Juss. (Vega et al., 2002). En el mismo sentido, se han estudiado 16 especies de los siguientes géneros: Banisteriopsis C.B. Rob. (p. ej., B. anisandra (A. Juss.) B. Gates, B. argyrophylla (A. Juss.) B. Gates. y B. campestris (A. Juss.) Little), Byrsonima Rich. ex Kunth. (p. ej., B. basiloba A. Juss., B. crassifolia (L.) Kunth y B. lancifolia A. Juss.) y Heteropterys Kunth (p. ej. H. anoptera A. Juss., H. byrsonimifolia A. Juss. y H. tomentosa A. Juss.), las cuales comparten la disposición del haz vascular en forma de arco, estomas paracíticos y tricomas glandulares y difieren en el tipo de mesófilo, floema, tipo de glándulas y disposición de los tricomas (Araújo-Silva et al., 2010).
Cabe resaltar que los estudios anteriores, además de proveer información taxonómica, también aportan conocimiento ecoanatómico, el cual toma relevancia ante los actuales escenarios de cambio climático, ya que su entendimiento permite la toma de decisiones sobre restauración y conservación de especies in situ o ex situ (Torres y Sanabria, 2011; Montaño-Arias et al., 2013).
Chapolagh-Paridari et al. (2013) sugieren que caracteres como la densidad estomática, el tamaño de la planta y el espesor de la lámina, están relacionados con factores ambientales, tales como la altitud, temperatura y precipitación. En este sentido, en Ribes ciliatum Humb. & Bonpl. ex Roem. & Schult., Lupinus meridanus Moritz ex C.P. Sm. y L. eremonomus C.P. Sm., se observó una correlación negativa entre la altura de la planta y el tamaño de la hoja con respecto a la altitud, y una positiva con la densidad estomática (Briceño et al., 2000; Jiménez-Noriega et al., 2015).
Ely et al. (2005) estudiaron la estructura foliar de Monochaetum meridense Naudin con relación a su hábitat, observando hojas glabras, con cutícula delgada y parénquima esponjoso, atributos que la caracterizan como una especie mesomórfica (Laskoswki, 2000). Por otra parte, Torres y Sanabria (2011) reportan cambios en el desarrollo de Cattleya jenmanii Rolfe y C. lueddemanniana Rchb. f. originados por el ambiente como es la reducción en el espesor de la lámina, epidermis y cutícula, y un mesófilo con ausencia de fibras y haces vasculares. Dichos cambios son estrategias para distribuirse sobre gradientes altitudinales y condiciones climáticas adversas (Molina-Montenegro y Cavieres, 2010).
A pesar de que existen estudios anatómicos foliares para la familia Malpighiaceae, éstos son escasos, en específico para el género Malpighia, ya que solo se registra un estudio en M. emarginata DC., indicando que tiene adaptaciones xeromórficas, por presentar epidermis uniestratificada, hojas hipoestomáticas y estomas paracíticos (Siddique y Ahmad, 1977). En el caso de M. mexicana no existen estudios sobre la anatomía de la hoja; esta especie ha sido reportada como sinónimo de M. glabra L. y de M. punicifolia L., antes llamada M. emarginata (Maldonado-Peralta et al., 2016). Sin embargo, existen caracteres morfológicos como altura del tronco, forma y tamaño de hojas, color de los pétalos, así como la disposición del estilo, tamaño del peciolo y tamaño del fruto, que deben estudiarse para definir dicha sinonimia. Por lo anterior, los objetivos de este estudio son: 1) describir las características anatómicas de la hoja de Malpighia mexicana y 2) determinar los caracteres anatómicos que pueden estar influenciados por el ambiente, como una forma de contribuir al conocimiento taxonómico y ecoanatómico del género Malpighia en México.
Materiales y Métodos
El estudio se realizó en la Faja Volcánica Transmexicana. Se seleccionaron tres sitios, dos en el Estado de México (Malinalco (S1) y Zumpahuacán (S2)), y el tercero en el estado de Morelos (Coatlán del Río (S3)). Para cada sitio de muestreo se revisó la temperatura, la precipitación (CONAGUA, 2017), el tipo de vegetación (Rzedowski, 2006) y clima (García, 1988) (Cuadro 1).
Cuadro 1: Estado, sitio de recolecta, tipo de vegetación, clima, coordenadas, altitud, recolector y número de recolecta, altura, temperatura y precipitación de los ejemplares de Malpighia mexicana A. Juss. utilizados para este estudio. LYBL: Lorena Yeraldi Bárcenas-López.
| Estado | Sitio de recolecta | Tipo de vegetación (Rzedowski, 2006) | Clima (García, 1988) | Coordenadas/ Altitud (m) | Recolector No. de recolecta | Altura (m) | Temperatura media anual (°C) | Precipitación anual (mm) |
| Estado de México | S1*Malinalco (Barrio de San Sebastián) | Bosque de pino-encino | Semi-cálido subhúmedo con lluvias en verano | 18°58.23'N 99°29.43'O, 2450 | LYBL 2 (CODAGEM), 4 (CODAGEM), 5 (CODAGEM) | 3.5, 4, 6 | 20 | 1375 |
| Estado de México | S2*Zumpahuacán (San Pedro Guadalupe) | Bosque tropical subcaducifolio | Semi-cálido subhúmedo con lluvias en verano | 18°50.1'N 99°34.43'O, 1706 | LYBL 5 (CODAGEM), 6 (CODAGEM), 8 (CODAGEM) | 3.5, 4.2, 5 | 18.2 | 1000 |
| Morelos | S3*Morelos (Coatlán del río) | Bosque tropical caducifolio | Cálido con lluvias en verano | 18º45.5'N 99º26.8'O, 1073 | LYBL 10 (CODAGEM), 18 (CODAGEM), 20 (CODAGEM) | 5.1, 4.3, 4.5 | 22.2 | 964 |
En cada sitio se seleccionaron tres individuos con alturas (3-4 m) y coberturas (4.70 y 6.20 m) similares. La copa de los individuos se dividió en cuadrantes (Montaño-Arias et al., 2018), se recolectaron cinco hojas por cuadrante; éstas se mezclaron para obtener una muestra compuesta (20 hojas/individuo). Las hojas se fijaron con FAA (1:1:1) y se mantuvieron a temperatura ambiente (18 °C). Los ejemplares de referencia correspondientes están depositados en el Herbario Eizi Matuda (CODAGEM), de la Universidad Autónoma del Estado de México (Cuadro 1).
En el laboratorio, las hojas se lavaron con agua corriente y se colocaron en dietilamina concentrada durante 24 h para su ablandamiento. Enseguida se deshidrataron en series ascendentes de alcoholes (30-100%), alcohol y xileno 50:50 y xileno (100%) por 6 h en cada concentración, terminando con una inclusión en parafina (50% de histoparafina y 50% de aceite vegetal) con punto de fusión de 60-65 ºC (Ruzin, 1999). Se realizaron cortes paradermales y transversales (15-20 µm) en un micrótomo de deslizamiento (American Optical 860, Nueva York, EUA). Se tiñeron con safranina-verde rápido y se montaron en resina sintética (Johansen, 1940).
La descripción anatómica de la hoja se realizó siguiendo la nomenclatura de Metcalfe y Chalk (1979). Las observaciones se efectuaron en un fotomicroscopio Leica DM 300 (Leica Microsystems, Heerbrugg, Suiza); se tomaron fotografías y se registraron mediciones lineales y de área con el analizador de imágenes Leica LV 40 (Leica Microsystems, Heerbrugg, Suiza). Se evaluaron 18 caracteres; para cada variable se tomaron 20 mediciones. Las variables evaluadas fueron: espesor de la lámina (µm, de cutícula adaxial a cutícula abaxial), espesor de la cutícula adaxial y abaxial (µm), largo y ancho de las células epidérmicas adaxiales y abaxiales (µm), espesor del parénquima en empalizada y del esponjoso (µm), tamaño de las células oclusivas (µm, largo y ancho) y la densidad de cristales (número de cristales/mm2).
En la vena media se registró el ancho y largo de las células epidérmicas (µm), el diámetro tangencial de los elementos de vaso (µm); la orientación de la medición de los vasos fue hacia la epidermis adaxial; así como el espesor del floema. De forma adicional, se calculó la densidad estomática (número de estomas/mm2) e índice estomático (IE). El índice estomático se calculó mediante la fórmula: Índice Estomático=(número de estomas/número de estomas+número de células epidérmicas) × 100; se realizaron cinco repeticiones de 1 mm2/individuo (Cañizares et al., 2003).
Por último, los datos obtenidos se examinaron con un análisis de varianza (ANOVA, p≤0.05), seguido por la prueba de Tukey-Kramer (p≤0.05). Los análisis estadísticos se llevaron a cabo mediante el paquete estadístico INFOSTAT (2017).
Resultados
Las hojas de Malpighia mexicana midieron 3 a 16 cm de largo y 2 a 8 cm de ancho, son ovadas, gruesas, pubescentes, de color verde brillante, con vena media sobresaliente. Anatómicamente las hojas de los tres sitios presentan características similares. En corte paradermal, se observó epidermis en ambas caras de la lámina, ésta consta de células poligonales y ovadas-rectangulares con paredes anticlinales lineales y gruesas (Figs. 1A-F), sin arreglo definido (Figs. 1C-D) y ambas superficies con apéndices epidérmicos con abundantes tricomas unicelulares caedizos de tipo malpighiáceo.

Figura 1: Epidermis en vista superficial en Malpighia mexicana A. Juss.: A. y B. epidermis adaxial; C. y D. epidermis abaxial. Barra=100 μm, E y F. tricomas. Barra=50 μm.
El espesor de la lámina fue mayor en hojas del sitio S2 (205.12 µm) en tanto que en las de S3 fue menor con 155.07 µm. En cuanto al espesor de la cutícula, éste mostró diferencias en ambas superficies. La cutícula adaxial más gruesa la presentan las hojas de S1, mientras que las hojas de S2 presentan la cutícula delgada. Sin embargo, la cutícula abaxial en las hojas de S2 fue la más gruesa y en las de S1 la más delgada (Cuadro 2).
Cuadro 2: Caracteres anatómicos mensurables de la hoja de Malpighia mexicana A. Juss. provenientes de tres sitios diferentes: S1: Malinalco, Estado de México, México; S2: Zumpahuacán, Estado de México, México y S3: Coatlán del Río, Morelos, México. Para cada carácter, los valores seguidos por la misma letra no difieren significativamente; mientras que los valores seguidos por letras diferentes, muestran diferencias significativas (p<0.05). F=significancia estadística y DE=desviación estándar.
| Variable | S1 | S2 | S3 | F |
| Media ± DE | ||||
| Espesor de la lámina (µm) | 156.09±0.77a | 205.12±1.45b | 155.07±9.02a | 1754.50 |
| Espesor de la cutícula adaxial (µm) | 5.36±0.23c | 3.53±0.27a | 4.49±0.25b | 793.74 |
| Espesor de la cutícula abaxial (µm) | 2.51±0.28a | 4.45±0.28c | 3.51±0.19b | 855.87 |
| Largo de la célula epidérmica adaxial (µm) | 27.79±9.65a | 35.62±14.73b | 38.36±13.35b | 11.09 |
| Ancho de la célula epidérmica adaxial (µm) | 20.27±2.68a | 24.74±5.92b | 22.92±9.15ab | 7.23 |
| Largo de la célula epidérmica abaxial (µm) | 22.12±5.95a | 23.26±9.16a | 22.83±11.86a | 0.23 |
| Ancho de la célula epidérmica abaxial (µm) | 13.67±2.24b | 17.56±2.74c | 10.23±1.64a | 158.77 |
| Espesor del parénquima en empalizada (µm) | 55.68±0.82a | 56.25±2.60a | 78.37±6.07b | 679.15 |
| Espesor del parénquima esponjoso (µm) | 29.59±3.55a | 76.90±1.47c | 46.53±1.55b | 6034.72 |
| Largo de las células oclusivas (µm) | 13.21±2.9b | 16.42±0.05c | 11.32±5.0a | 426397.06 |
| Ancho de las células oclusivas (µm) | 15.12±3.4b | 28.95±0.01c | 12.16±4.9a | 232349025 |
| Densidad de cristales (µm) | 6-10 | 8-10 | 10 | 0.24 |
| Largo de las células epidérmicas vena media (µm) | 31.88±0.54c | 24.48±7.73b | 21.36±4.57a | 64.95 |
| Ancho de las células epidérmicas vena media (µm) | 23.17±0.49b | 23.66±7.77b | 18.56±3.66c | 19.26 |
| Diámetro tangencial de los vasos (µm) | 20.72±7.35b | 21.56±0.23b | 18.58±0.20a | 7.86 |
| Espesor del floema (µm) | 105.49±0.23a | 106.36±0.24b | 133.40±0.24c | 269743.04 |
| Densidad estomática (µm) | 76a | 92b | 143c | 738.56 |
| Índice estomático (µm) | 18.14a | 18b | 18.32c | 0.13 |
Las hojas de los tres sitios presentaron apéndices epidérmicos en ambas superficies de la lámina, éstos fueron simples y ramificados (Figs. 1E, F). Con relación al tamaño de las células epidérmicas adaxiales, las hojas del sitio S1 presentaron células pequeñas y las células epidérmicas abaxiales fueron más grandes en el S2. Cabe señalar que en S3 las células son más angostas, en tanto que el largo de las células se mantuvo constante en las hojas de los tres sitios (Cuadro 2).
Con relación a las células oclusivas de los complejos estomáticos se observó que las hojas provenientes de S2 tienen en promedio el mayor tamaño (16.42 × 28.95 µm) y las de S3 son las más pequeñas (11.32 × 12.16 µm) (Cuadro 2).
En corte transversal, la epidermis es uniestratificada con cutícula lisa y gruesa y células rectangulares ocasionalmente introducidas en el mesófilo, éste presentó un arreglo bifacial (Figs. 2A-C). Además, el mesófilo de la superficie adaxial tiene cristales clasificados como drusas, con una densidad de 6-10 drusas/mm2; los cuales se observaron tanto solitarios como agrupados. El mesófilo está compuesto de 2 a 4 estratos de parénquima en empalizada con cloroplastos. El parénquima esponjoso estuvo constituido de 6 a 8 estratos de células con presencia de espacios intercelulares. El parénquima en empalizada de las hojas del sitio S3 registró un mayor espesor (78.37 μm), mientras que en las de S1 y S2 el tamaño fue similar (Cuadro 2). Respecto al espesor del parénquima esponjoso, en las hojas de S2 tuvo mayor espesor (76.90 μm), mientras que el de menor se registró en las hojas de S1.
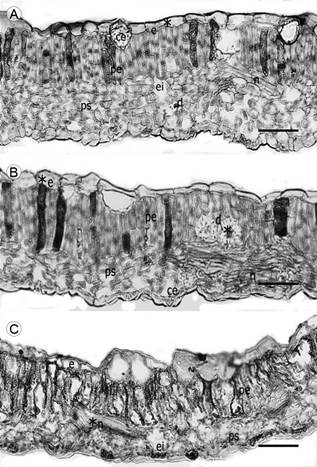
Figura 2: Corte transversal en hoja de Malpighia mexicana A. Juss.: A. Malinalco (S1); B. Zumpahuacán (S2); C. Morelos (S3): e=epidermis, d=drusas, pe=parénquima de empalizada, ps=parénquima esponjoso, ce=cámara estomática, ei=espacios intercelulares, n=nervadura. Escala=100 μm.
La vena media está conformada por un haz colateral en forma de arco hacia la superficie adaxial. La epidermis tiene una capa de células rectangulares a ovadas de tamaño variable con cutícula lisa (Figs. 3A-C). Cabe resaltar que, independientemente del sitio de recolecta, no se observó parénquima en empalizada ni parénquima esponjoso. En las hojas de S3, por debajo de la epidermis, se observaron de 3 a 5 estratos de células de colénquima angular (Fig. 3C). El xilema se encontró dispuesto radialmente formado de 3 a 8 estratos de células, alternando una hilera de vasos con una de parénquima xilemático. Los sitios S1 y S2 presentaron los vasos con el mayor diámetro tangencial (20.72 y 21.56 μm, respectivamente) (Cuadro 2). El floema se encuentra en forma de arco, hacia la superficie adaxial, conformado por 10 a 14 estratos de células, cabe resaltar que en las hojas de S3 se registró el mayor espesor.
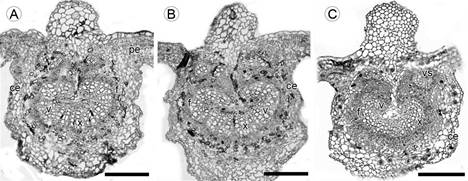
Figura 3: Corte transversal de la vena media de la hoja de Malpighia mexicana A. Juss: A. Malinalco (S1); B. Zumpahuacán (S2); C. Morelos (S3): x=xilema, ce=células epidérmicas, f=floema, pe=parénquima, v=vasos, vs=vena secundaria. Escala=200 μm.
De forma adicional, se calculó el índice estomático, éste no presentó diferencias significativas entre sitios; sin embargo, la densidad estomática fue diferente en los tres sitios. Las hojas del sitio S3 exhibieron la mayor cantidad de estomas con 143 mm2, mientras que las de los sitios S1 (76 mm2) y S2 (92 mm2) tienen la menor densidad, aunque éstos son más grandes (Cuadro 2).
Discusión
Interpretación taxonómica
La familia Malpighiaceae resulta difícil de estudiar debido a la variabilidad morfológica y sinonimia de sus especies (Gates, 1982; Anderson, 2001 a, b). Por ejemplo, Malpighia emarginata tiene ocho sinónimos y M. mexicana tiene tres subespecies y una forma (TROPICOS, 2017). Los caracteres como la forma, longitud, ancho y pubescencia de la hoja se utilizan para identificar taxonómicamente las especies de la familia Malpighiaceae (Anderson, 1979).
De acuerdo con los resultados obtenidos, M. mexicana presenta hojas hipoestomáticas con abundancia de tricomas en ambas superficies de la lámina, células epidérmicas alargadas y grandes, vena media en forma de arco y el mesófilo con un arreglo bifacial. Los caracteres con relevancia taxonómica son la presencia de tricomas unicelulares simples y ramificados en ambas superficies de la lámina, aunque la presencia de tricomas unicelulares simples se ha reportado en M. emarginata, pero éstos solo están presentes en la parte abaxial (Laskowski, 2000).
La epidermis uniestratificada presente en M. mexicana también ha sido observada en especies de los géneros Banisteriopsis, Byrsonima y Heteropterys (Araújo-Silva et al., 2010), por lo que ésta no presenta relevancia taxonómica. En cuanto al mesófilo dorsiventral, éste es propio de las eudicotiledóneas (Arambarri et al., 2011), dato que concuerda con lo reportado por Attala (1997); Laskoswki (2000) y Pereira (2002) para la familia Malpighiaceae.
En este estudio se encontraron complejos estomáticos de tipo paracítico, al igual que lo reportado por Araújo-Silva et al. (2010), en 16 especies de la familia Malpighiaceae, lo que coincide con Metcalfe y Chalk (1979), quienes señalan que estos predominan en las malpigiáceas. Sin embargo, éstos no son exclusivos de las malpigiáceas, ya que han sido reportados en integrantes de otras familias, como Cactaceae (Eggli, 1984), Gesneriaceae (Ramírez-Roa y Varela-Hernández (2011) y Fabaceae (Montaño-Arias et al., 2018), entre otras.
El índice estomático obtenido en M. emarginata es de 16% y los valores encontrados en este estudio para M. mexicana son de 18%. Schoch et al. (1980) señalan que el índice estomático es una función del total de la radiación recibida y de las variaciones que ésta experimenta durante el desarrollo de la hoja. De acuerdo con Leegod (1993), existe un rango de 40 a 300 estomas/mm2 en la parte abaxial de las hojas de plantas C3; en consecuencia, esta característica no puede ser considerada con valor taxonómico por la amplitud del rango mencionado.
El haz vascular en forma de arco, el xilema dispuesto radialmente y la posición del floema en la vena media presentes en M. mexicana son caracteres considerados constantes en el género Malpighia por Metcalfe y Chalk (1979) y Laskoswki (2000). Estos caracteres han sido reportados también en especies de Lupinus de la familia Leguminosae (Zamora-Natera y Terrazas, 2012), por su parte; Araújo-Silva et al. (2010) registraron la presencia de tejido vascular en forma de arco y presencia de floema en la médula de Byrsonima, excepto B. coccolobifolia Kunth. En este estudio, el xilema y el floema se encuentran en forma de arco pero no se observó floema en la región medular de la vena media.
Interpretación ecoanatómica
Las hojas de los tres sitios presentaron características que corresponden con la estructura de tipo de hoja mesomórfica como mesófilo dorsiventral, abundancia de espacios intercelulares y estomas restringidos a la superficie abaxial. Además, se observaron caracteres xeromorfos como células del parénquima en empalizada largas y estrechas, presencia de cutícula gruesa en epidermis superior y estomas restringidos a la superficie abaxial (Lindorf, 1994; Araque et al., 2009).
El análisis de los resultados muestra que M. mexicana conserva su estructura anatómica en las hojas de los tres sitios. No obstante, se observaron diferencias en algunos caracteres cuantitativos, como en el espesor de la lámina, cutícula y largo y ancho de las células epidérmicas; estas diferencias podrían deberse al sitio donde se desarrolla la especie, a un déficit hídrico y a altas temperaturas (Laskowski, 2000).
Las hojas de S3 presentaron mayor abundancia de tricomas; el sitio presenta la temperatura más alta, 22.2 ºC (CONAGUA, 2017), lo que podría sugerir que la pubescencia surge como estrategia para mitigar las ondas de calor (Molina-Montenegro y Cavieres, 2010). En los S1 y S2 que presentan menor temperatura (20 °C y 18.2 °C, respectivamente), la densidad de tricomas de las hojas disminuyó, dato que concuerda con lo reportado por Jiménez-Noriega et al. (2015), quienes observaron un incremento en la densidad de tricomas en hojas de Ribes ciliatum conforme aumenta la temperatura del sitio donde se desarrollan.
De acuerdo con Granada-Chacón y Benítez de Rojas (2004), la presencia de una cutícula gruesa y células grandes con posible función de almacenamiento de agua son caracteres propios de especies caducifolias. Malpighia mexicana concuerda con estas características y es una especie caducifolia. Sin embargo, existe variación en el espesor de la cutícula de la hoja entre sitios, lo cual podría ser el resultado de la diferencia de humedad entre ellos. Por ejemplo, el S1 presenta una precipitación de 1375 mm y cutícula gruesa, mientras que en los S2 y S3 es más delgada con 1000 mm y 964 mm respectivamente (CONAGUA, 2017), lo que podría explicar el engrosamiento de la cutícula. Datos similares fueron reportados por Araújo-Silva et al. (2010), en especies que habitan climas secos (Banisteriopsis anisandra, B. campestris, Byrsonima coccolobifolia, B. basiloba, Heteropterys byrsonimifolia y H. anoptera), donde observaron variación en el grosor de la cutícula. Según Yeats y Rose (2013), esta característica ejerce algún efecto sobre la resistencia estructural de las plantas; además, la cutícula es la línea de defensa de los vegetales ante los ataques de patógenos (Agrios, 2010).
La conformación del mesófilo bifacial y la combinación de características meso y xeromórficas es típica de plantas adaptadas a altas condiciones de luz y alta disponibilidad de agua (Smith et al., 1997), lo que indica que M. mexicana puede estar adaptada a esas condiciones. Las hojas de los sitios S1 y S2 mostraron una disminución en el espesor de la lámina a diferencia del otro sitio, esto puede deberse a que se localizan a mayor altitud y cuenta con una precipitación anual mayor (Cuadro 1), lo que coincide con lo señalado por Zamora-Natera y Terrazas (2012), quienes reportan que las especies de Lupinus que conviven y crecen a mayor altitud mostraron menor espesor de la lámina y del mesófilo, que las especies que crecen a menor altitud.
La presencia de parénquima esponjoso laxo observado en especies de las familias Asteraceae, Solanaceae y Malpighiaceae (Ragonese, 1990; Milan et al., 2006; Araújo-Silva et al., 2010) se ha sugerido como un amortiguador en cambios bruscos de temperatura en el ciclo diario, además de proporcionar rigidez a la lámina foliar (Ely et al., 2005). Malpighia mexicana se caracteriza por tener varios estratos de células parenquimáticas en empalizada, condición que favorece la mayor producción de fotoasimilados y polisacáridos, que se puede reflejar en un mejor crecimiento vegetativo y reproductivo que ayude a la producción de un mayor número de frutos y de mayor calidad (Terashima et al., 2011; Rodríguez et al., 2016).
Por otro lado, Sack y Scoffoni (2013) reportan que el diámetro de los vasos del xilema podría ser crucial para aumentar la eficiencia hidráulica con relación a la altura del árbol y, por lo tanto, permitir un equilibrio en el suministro de agua. Las hojas provenientes del S3 fueron las que presentaron diámetros menores; además, fue el sitio con temperatura más baja lo que podría significar menores tasas de transpiración en la planta.
De igual forma, Engelbrecht et al. (2000) mencionan que las especies maderables que habitan regiones más húmedas registran diámetros y longitudes de los vasos mayores, en comparación con las que habitan ambientes secos. Esto coincide con lo observado en las hojas de Malpighia mexicana donde los individuos provenientes de sitios con mayor precipitación presentan vasos con diámetros mayores en la hoja, lo cual facilita un adecuado flujo de agua y nutrientes (Roth et al., 1986).
La presencia de cristales tipo drusas en los individuos de los tres sitios favorece el desarrollo de la planta. Según Nitta et al. (2006) las drusas ayudan a la reabsorción de calcio en condiciones de deficiencia, previenen toxicidad y dan soporte estructural. Finley (1999) señaló que estas sustancias son tóxicas y su acumulación es una forma de excreción que es aprovechada por la planta para prevenir el consumo por parte de los herbívoros, pues los cristales provocan irritación de las mucosas bucales en estos animales.
Conclusiones
La hoja de Malpighia mexicana conserva su estructura anatómica independientemente del sitio de colecta. Los caracteres taxonómicos que permiten identificarla son la disposición bifacial de los tricomas de tipo simple y ramificado y la posición de floema en el nervio medio. Se registraron diferencias cuantitativas con respecto al espesor de la lámina y el mesófilo entre los sitios estudiados, diferencias que se deben a la altitud, precipitación y temperatura de los sitios.
Malpighia mexicana presenta una epidermis con células grandes y una mayor concentración de estomas en la superficie abaxial que la definen como una especie xeromórfica; sin embargo, muestra hojas dorsiventrales, espacios intercelulares en el mesófilo y ausencia de esclerénquima, que la caracterizan como mesomórfica. Por lo anterior, se puede concluir que M. mexicana es una especie mesomórfica, con caracteres xeromórficos que le ayudan adaptarse a variaciones en la precipitación y temperatura, por lo que podría ser una especie que tolere el cambio climático.

