











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Tropical montane cloud forest (TMCF) is a physiognomically heterogeneous floristic group composed of species with diverse biogeographic affinities. It is characterized by the persistent cloud cover that envelops its vegetation (Williams-Linera, 2007). This vegetation type stands out for its high levels of plant and fungal diversity (Gómez-Hernández et al., 2012). It occupies less than 1% of the national territory (Challenger and Dirzo, 2009), yet contributes 10% of plant diversity and exhibits a high degree of endemicity (Rzedowski, 1993). However, in Mexico, TMCF is also one of the most threatened ecosystems (Medel, 2013; Estrada-Contreras et al., 2015).
One conservation strategy for countering the current loss of TMCF is the establishment of areas for preserving the biodiversity and ecosystem services provided by these forests (Toledo-Aceves et al., 2011). Natural protected areas (NPAs) play a key role as reservoirs of the biodiversity associated with TMCF in human-dominated landscapes (MacGregor-Fors et al., 2016). However, the conservation value of NPAs may be spatially dependent on structural (e.g., habitat loss) and/or landscape factors (e.g., surrounding land uses/covers) (Gove et al., 2005; Escobar et al., 2008). Until now, little is known about whether the management activities of NPAs temporally impact their biodiversity. Thus, long-term studies are necessary to understand the conservation value of NPAs under anthropogenic management regimes.
Biological communities associated with TMCF are highly dynamic in space and time as a result of seasonality, habitat characteristics, and the degree of anthropogenic pressures (Gómez-Hernández and Williams-Linera, 2011; García-Martínez et al., 2016; MacGregor-Fors et al., 2016). Particularly, in the present context, temporal shifts experienced by biological communities (including those of TMCF) may be a cumulative consequence of historical changes in anthropogenic management activities (Toledo-Aceves et al., 2011). In addition, temporal variation in diversity (i.e., species richness and abundance) and compositional similarity (i.e., presence/absence of species) of a particular species group depend on the response ability and the degree of taxonomic resolution of the group as well as how the community is defined (Rahel, 1990).
Among the impressive biodiversity harbored by Mexican TMCFs, macrofungi form one of the most diverse groups (Guzmán et al., 2003; Gómez-Hernández et al., 2012). Some studies suggest that Ascomycota and/or Basidiomycota assemblages rapidly respond to natural and anthropogenic disturbances and management regimes (Gómez-Hernández and Williams-Linera, 2011; McGregor-Fors et al., 2015). Among ascomycetes, the Xylariaceae family, composed of 85 genera worldwide, is large and relatively well known (Kirk et al., 2008). Most are saprobic wood degraders and actively decay the wood of living or dead angiosperms, while others are endophytic and symbiotically live within plant tissues. In fact, several endophytes are commonly associated with numerous tropical plants and even associated with termite nests (Rogers, 2000; Rogers et al., 2005).
The genus Xylaria Hill ex Schrank is characterized by stromata, usually, erect, cylindrical or clavate, sometimes subglobose, usually higher than wide, surrounded by perithecia containing ascospores; stipitate perithecia with hemispherical or umbilicate ostioles; cylindrical asci with amyloid apical rings, black, ascospores usually with a straight, oblique, or spiraling germ slit (San Martín and Rogers, 1995). This genus is one of the most diversified in the family and contains about 300 species (Kirk et al., 2008). In Mexico, 105 species have been recorded (San Martin and Rogers, 1989; 1995), yet only 18 species are found in TMCF (Medel, 2013). Medel et al. (2008) recorded 45 species of Xylaria for Veracruz, of which 24 species have been reported for TMCF. Evaluating changes in the richness, abundance, or turnover of Xylaria assemblages can potentially be used as a tool for establishing conservation priorities or determining the impact of management activities on endangered ecosystems, including TMCF.
This study integrates a historical Xylaria inventory coming from a series of samples collected over the last 36 years in a natural protected area of tropical mountain cloud forest and it is improved with an exhaustive review of additional specimens available in the Fungi Collection of the herbarium XAL of the Instituto de Ecología, A.C. (INECOL) in Xalapa, Veracruz, Mexico. The aim of this study was to assess the temporal shifts over the last 36 years in the alpha, beta, and gamma diversity of a Xylaria assemblage in a NPA of TMCF that has experienced several anthropogenic management activities. Specifically, we examined whether the richness and abundance of Xylaria species changed over time, in addition to temporal patterns in the compositional similarity of the Xylaria assemblage. The implementation of long-term ecological monitoring using Xylaria assemblages as a model group could provide novel insight into the impacts of management and conservation strategies on NPAs and TMCF, one of the most threatened tropical ecosystems worldwide.
Material and Methods
Study area
This study was conducted in a fragment of tropical mountain cloud forest in the central mountainous region of the state of Veracruz, Mexico (Fig. 1). The climate is humid temperate, and the average annual temperature is 18 °C. The annual precipitation varies from 1500 to 2000 mm. A hot-dry season spans from March to May, a warm-rainy season from June to October, and a cold-dry season from November to February. The soil is characterized as Andosol, with a volcanic origin and loamy texture (Williams-Linera et al., 2002; 2015).
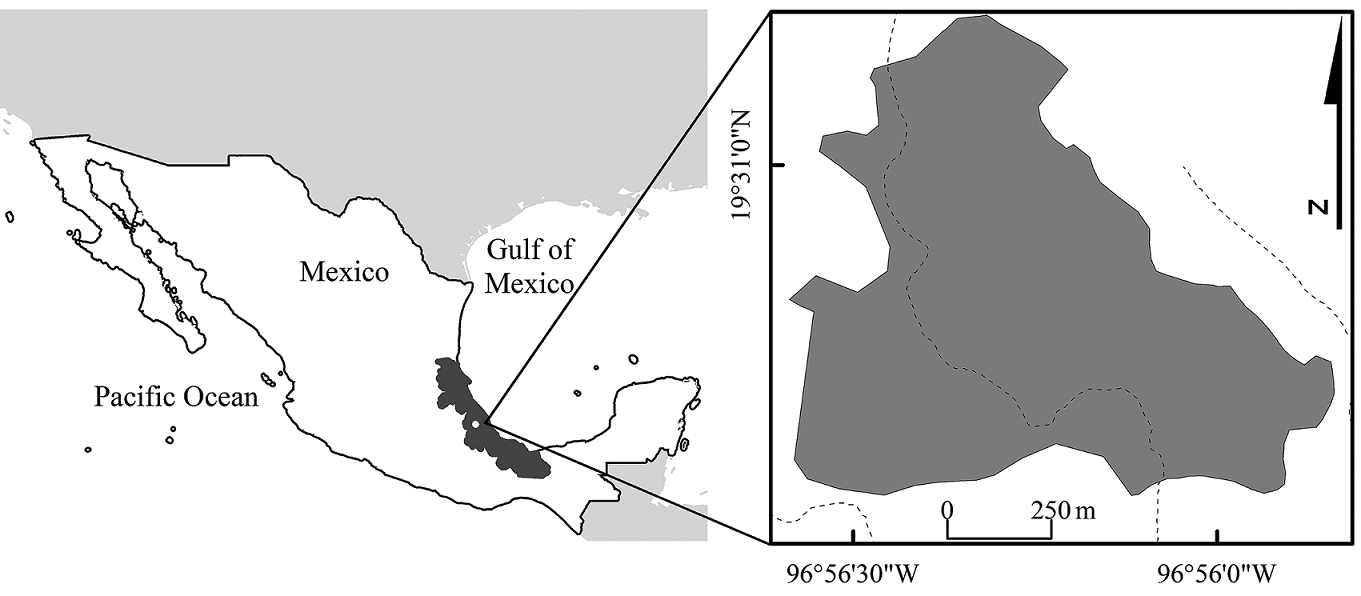
Figure 1: Location of a natural protected area of tropical montane cloud forest, Santuario del Bosque de Niebla, in central Veracruz, Mexico. The location of the study area (white circle) within the state of Veracruz (dark gray polygon) is shown. The dashed lines indicate the rivers and/or streams.
The studied forest is called Santuario del Bosque de Niebla and is located between the parallels 19°30'37''N; 19°31'06''N and the meridians 96°56'05''W; 96°56'27''W in the municipality of Xalapa, Veracruz, at an elevation ranging from 1250 to 1350 m a.s.l. (Fig. 1). This 31 ha sanctuary has been under state protection since February 17th, 1977, and currently belongs to the INECOL (Luna and García, 1998). The terrain of this forest has a slope of 52% and a soil compaction of 10 kg/m2. Ground cover is composed of leaf-litter (72%), grass (10%), and forbs and herbs (8%), and the remaining proportion is uncovered. Tree canopy cover is 79%, and canopy height is 11 m (García-Martínez et al., 2016). A total of 97 tree species are present (Lorea Hernández, pers. com.) at a density of 1140 individuals/ha (García-Martínez et al., 2016).
Significant events have occurred in the study area before the considered sampling periods that have changed its land use/land cover or management. Before 1980, the entire area was a shaded coffee plantation with some planted citrus trees, and a portion of it was modified to create a vacational/recreational center. Additionally, nine hectares were dedicated to creating a botanical garden, and an urban park with impressive infrastructure was constructed throughout the remaining area (Luna and García, 1998). From 1980 to 1989 the urban park was abandoned, and restoration activities began. The area came to be composed of a mosaic of habitats, including TMCF, secondary and riparian vegetation, as well as grasslands. The active restoration of the TMCF was later halted. Between 1990 and 1999 the area was converted to an ecological urban park dedicated to conserving, maintaining, and restoring native plant communities. The area contained relictual, disturbed, and heterogeneous fragments of TMCF. Natural succession continued, and the area was devoted to scientific research (Luna and García, 1998). Since 2000, the area was characterized as a young forest with a high canopy cover but low canopy height (Gómez-Hernández and Williams-Linera, 2011). Recently, the area may be categorized as a young forest remnant that, by now, is relatively undisturbed or as a secondary cloud forest undergoing maturation processes (MacGregor-Fors et al., 2016).
Collections and specimen revisions
Most Xylaria stromata are restricted to the rainy part of the year, so the study site was sampled between July and November from 2010 to 2016 following the sampling technique of Lodge et al. (2004). In every sampling event, the sporocarps of Xylaria were counted and collected in thin paper bags and each specimen was macro and micromorphologically described when fresh. Then, the specimens were dried in an oven at 50-60 °C for 48 h (Lodge et al., 2004). To improve our Xylaria inventory, we also examined all specimens of Xylaria collected in the same study area and deposited between 1981 and 2010 in the Fungi Collection of the herbarium XAL. All specimens collected and obtained during our explorations are available in XAL, and these were determined at the species level by identifying their micro and macromorphological characters according to several keys and specialized literature (Dennis, 1956, 1957; Rogers, 1983, 1984; Rogers and Callan, 1986; Rogers et al., 1988, 2002; San Martín and Rogers, 1989; San Martín, 1992; Medel et al., 2008; Hladki and Romero, 2010; Tapia et al., 2017).
Data analysis
As Xylaria species have a cespitose and gregarious growth form, two or more sporocarps of the same species growing in a diameter of <50 cm or in a fairy ring were recorded as one individual. For correcting the bias implicated in herbarium revision, we based the abundance on the incidence of Xylaria species (i.e., the number of times that a given species was collected during a sampling period). We grouped all Xylaria records (species and abundance records) in four sampling periods according to the year when specimens were collected: 1980-1989, 1990-1999, 2000-2009, and 2010-2016 (three periods of 10 years and one period of six).
We calculated the inventory reliability for each sampling period using the coverage estimator, which is a less biased estimator of sample completeness than non-parametric methods. The resulting value indicates the proportion of the statistical population that is represented by the collected species (Chao and Jost, 2012) and is expressed by the following equation:
where n is the sum of all individuals of all species in a given sampling period, and f 1 and f 2 are singletons (species each represented by only a single individual) and doubletons (species each represented by exactly two individuals), respectively. Sample completeness (Ĉn) indicates the proportion of the “total community” represented by the collected species and enables the diversity of assemblages sampled at the same coverage to be compared. When Ĉn≈100%, sampling is complete given the effort and utilized collection technique (Chao and Jost, 2012). Values of Ĉn were calculated using the iNEXT package for R (Hsieh et al., 2016).
As mentioned, the comparison of richness among sampling periods is only ecologically appropriate for those periods with a similar level of inventory completeness. Therefore, a comparison of our estimates of species richness per sampling period could be biased because of differences in inventory completeness. In particular, species richness is sensitive to variations in the number of singletons and doubletons (Chao and Jost, 2012). Thus, we estimated the species richness for each sampling period using coverage-based inter- and extrapolations in the iNEXT package for R (Hsieh et al., 2016). We considered 99.99% completeness as a reliable estimator of richness for all sampling periods (Chao and Jost, 2012). To compare species richness among sampling periods, we used 95% confidence intervals in which significant differences were indicated by non-overlapping confidence intervals (Cumming et al., 2007).
To evaluate differences in species’ dominance and rarity and in community evenness among sampling periods and, thus, to better interpret our results, Xylaria abundance was represented by rank-abundance species curves or Whittaker plots (Magurran, 2004). We plotted the proportional abundance of each species, ordered from the most to the least abundant, to show differences in species’ dominance and rarity and in assemblage evenness among sampling periods.
To analyze beta diversity, we determined the compositional similarity among sampling periods using the Jaccard index. This index has values ranging from 0 (minimal similarity) to 100 (maximum similarity) (Magurran, 2004). It only takes into account shared species and presence/absence of species between sites (Jost, 2006). The compositional similarity among sampling periods was represented by a cluster analysis using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) linkage technique. For the post hoc analysis, a similarity profile test (SIMPROF) was used as statistical test to compare similarity among assemblages in the PRIMER software version 6.1.16 (Clarke and Gorley, 2006). The SIMPROF test assumes that a real clustering of assemblages will be evidenced by an excess of smaller and/or larger similarities than expected under the null hypothesis that all assemblages are drawn from the same species assemblage (Clarke et al., 2008).
To quantify the contribution of each time period to total diversity (i.e., gamma diversity), we calculated the average number of species absent from each site (beta diversity), which is defined as β=γ-α, where γ corresponds to the number of species recorded historically in the study area (gamma diversity) and α to the average number of species present in a given time period (alpha diversity) (Lande, 1996). This approach allows for a direct comparison between alpha and beta diversities in terms of numbers (or percentages) of species (Crist et al., 2003).
Results
The Xylaria inventory
In total, we historically recorded 3480 individuals belonging to 30 species of the genus Xylaria (Table 1). The most abundant species was X. scruposa (Fr.) Berk. (903 individuals), followed by X. anisopleura (Mont.) Fr. (820), X. berteroi (Mont.) Cooke ex J.D. Rogers & Y.M. Ju (496), X. cubensis (Mont.) Fr. (435), X. feejeensis (Berk.) Fr. (231), X. albocinctoides Y.M. Ju, H.M. Hsieh & J.D. Rogers (153), and X. arbuscula Sacc. (136). The 23 remaining species were represented by less than 80 individuals. The average sampling completeness was 95.6%. Sampling completeness was 90.2% in the sampling period of 1980-1989, 98% in 1990-1999, 94.2% in 2000-2009, and 99.9% in 2010-2016. Considering all four decades, the sampling completeness was 99.9%.
Table 1 Historical inventory of the Xylaria Hill ex Schrank assemblages recorded over the last 36 years (four sampling periods) in a natural protected area of tropical montane cloud forest, Santuario del Bosque de Niebla, in central Veracruz, Mexico. *New records for Mexico, **New records for Veracruz, ***Possible new species for science.
| Species | 1980-1989 | 1990-1999 | 2000-2009 | 2010-2016 |
| 1. Xylaria albocinctoides Y.M. Ju, H.M. Hsieh & J.D. Rogers * | 0 | 0 | 1 | 152 |
| 2. Xylaria allantoidea (Berk.) Fr. | 6 | 0 | 0 | 0 |
| 3. Xylaria anisopleura (Mont.) Fr. | 3 | 0 | 3 | 814 |
| 4. Xylaria arbuscula Sacc. | 0 | 0 | 0 | 136 |
| 5. Xylaria berteroi (Mont.) Cooke ex J.D. Rogers & Y.M. Ju | 15 | 21 | 19 | 441 |
| 6. Xylaria cubensis (Mont.) Fr. 1851 | 10 | 18 | 0 | 407 |
| 7. Xylaria curta Fr. | 3 | 3 | 9 | 63 |
| 8. Xylaria feejeensis (Berk.) Fr. 1851 | 1 | 2 | 12 | 216 |
| 9. Xylaria discolor (Berk. & Broome) Y.M. Ju, H.M. Hsieh, J.D. Rogers & Jaklitsch * | 0 | 0 | 0 | 1 |
| 10. Xylaria grammica (Mont.) Mont. | 0 | 0 | 1 | 5 |
| 11. Xylaria heliscus (Mont.) J.D. Rogers & Y.M. Ju | 0 | 0 | 0 | 3 |
| 12. Xylaria hypoxylon (L.) Grev. | 0 | 1 | 2 | 3 |
| 13. Xylaria ianthinovelutina (Mont.) Fr. | 0 | 0 | 1 | 5 |
| 14. Xylaria liquidambaris J.D. Rogers, Y.M. Ju & F. San Martín | 0 | 0 | 0 | 15 |
| 15. Xylaria longipes Nitschke | 0 | 0 | 0 | 3 |
| 16. Xylaria microceras (Mont.) Berk. | 0 | 0 | 0 | 3 |
| 17. Xylaria multiplex (Kunze) Fr. | 1 | 0 | 0 | 65 |
| 18. Xylaria scruposa (Fr.) Fr. | 1 | 2 | 3 | 897 |
| 19. Xylaria telfairii (Berk.) Sacc. | 1 | 0 | 0 | 2 |
| 20. Xylaria xylarioides (Speg.) Hladki & A.I. Romero * | 0 | 0 | 0 | 6 |
| 21. Xylaria sp. 1 | 0 | 0 | 0 | 6 |
| 22. Xylaria sp. 2 | 0 | 0 | 0 | 21 |
| 23. Xylaria sp. 3 *** | 0 | 0 | 0 | 36 |
| 24. Xylaria sp. 4 ** | 0 | 0 | 0 | 6 |
| 25. Xylaria sp. 5 ** | 0 | 0 | 0 | 1 |
| 26. Xylaria sp. 6 *** | 0 | 0 | 0 | 21 |
| 27. Xylaria sp. 7 ** | 0 | 0 | 0 | 1 |
| 28. Xylaria sp. 8 | 0 | 0 | 0 | 6 |
| 29. Xylaria sp. 9 ** | 0 | 0 | 0 | 3 |
| 30. Xylaria sp. 10 ** | 0 | 0 | 0 | 3 |
Alpha diversity
With respect to species abundance, 41 individuals were recorded in the sampling period of 1980-1989, 47 in 1990-1999, 50 in 2000-2009, and 3341 in the final sampling period (2010-2016) (Table 1). We observed that the assemblage structure changed across the four sampling periods. Considering proportional abundance, the general pattern observed indicated an increase in species dominance during the sampling period of 1990-1999 (Fig. 2). The dominant species during the periods of 1980-1989, 1990-1999, and 2000-2009 was X. berteroi, with a relative abundance of greater than 35%. With respect to observed species richness (Fig. 3), nine species were recorded in the sampling period of 1980-1989, six in 1990-1999, nine in 2000-2009, and 28 in the final sampling period (2010-2016). Upon comparing estimated species richness, we observed that the number of species recorded in the final sampling period (2010-2016) significantly differed from that of the other sampling periods.

Figure 2: Rank-abundance curves of the Xylaria Hill ex Schrank assemblage associated with a natural protected area of tropical montane cloud forest, Santuario del Bosque de Niebla, in central Veracruz, Mexico. Only species with a relative abundance higher than 10% (above dashed line) in a given sampling period are shown.
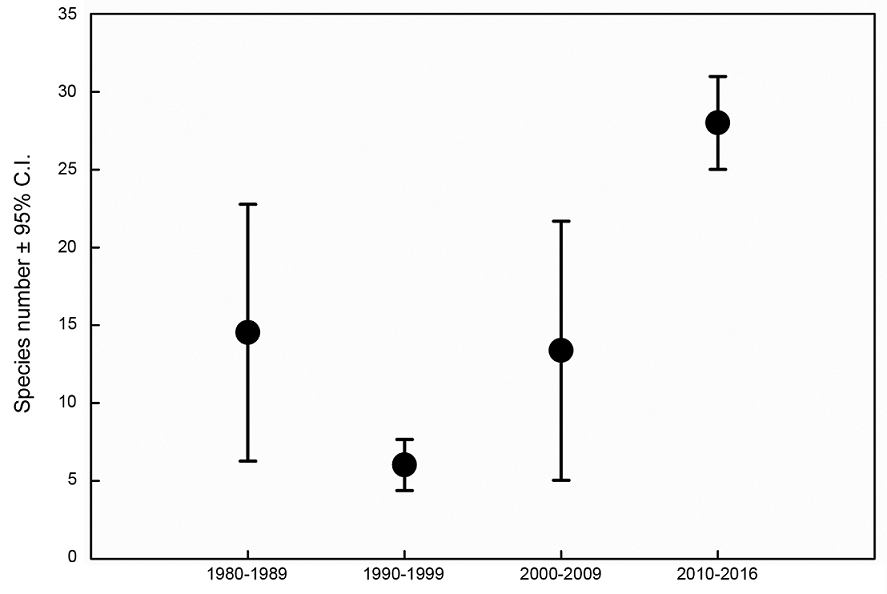
Figure 3: Comparison of species richness estimates for a rarefied and extrapolated sample with a size up to double the reference sample. Differences are considered to be statistically significant when the 95% confidence intervals (C.I.) do not overlap; for overlapping intervals, no differences are assumed (α=0.05).
Beta diversity
The evaluation of compositional similarity using a cluster analysis and SIMPROF test based on the Jaccard index showed the significant separation of two assemblage clusters at a similarity of 27% (π=4.65, p=0.002) (Fig. 4). The Xylaria assemblage collected in the 2010-2016 sampling period was significantly separated from the assemblages collected in the other periods. Regarding species turnover among sampling periods, 59% of total species were unique to a single sample period, and 14% were shared by all samplings (Fig. 5). During the 2010-2016 sampling period, 16 unique species were found; during the 1980-1989 period, only one unique species was found. During the other sampling periods, no unique species were found.
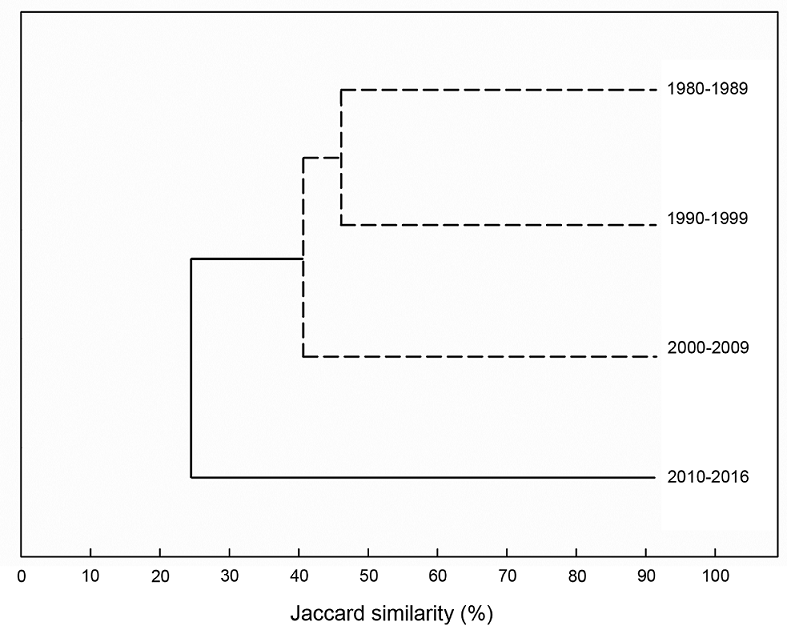
Figure 4: Dendrograms of the hierarchical clustering of Xylaria Hill ex Schrank assemblages recorded in four sampling periods based on the Jaccard similarity index. The dendrograms display as continuous lines the divisions for which the SIMPROF test rejects the null hypothesis (assemblages in these groups have no further structure to explore); the groups of assemblages not separated by the SIMPROF test (at p<0.05) are indicated by dashed lines.

Gamma diversity
The gamma diversity (γ) (i.e., historically recorded diversity) of the studied Xylaria assemblages was equal to 30 species. The estimated alpha diversity (α) varied from six to 12 species, with an estimated mean alpha of 10 species. Using an additive approach to evaluate the contribution of alpha (α) and beta diversity (β) to gamma diversity (γ), the observed mean alpha diversity accounted for 37% and beta diversity for 63% (Fig. 6).
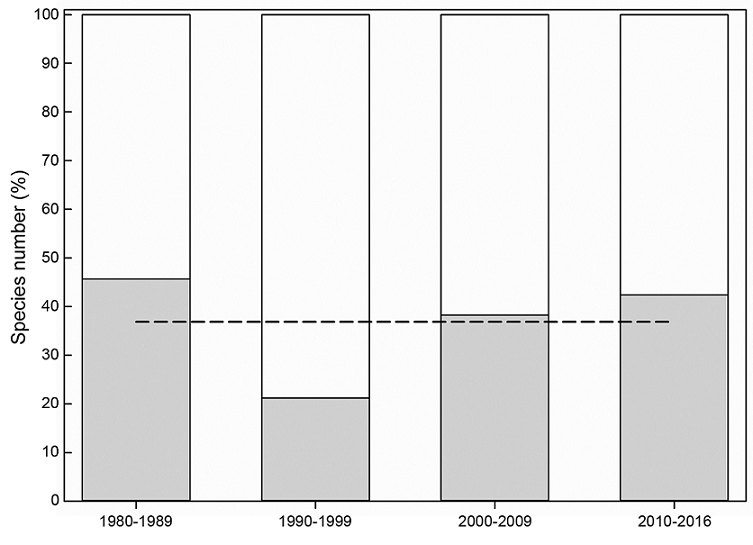
Figure 6: Contribution of the alpha and beta diversity of Xylaria Hill ex Schrank species recorded in four sampling periods to historical gamma diversity. The gray bars represent alpha diversity per sampling period, the white bars beta diversity, and the dotted line mean alpha diversity. All diversities are expressed as a percentage of total recorded species (n=30).
Discussion
This study provides the first historical inventory of Xylaria, mycobiota associated with a NPA of TMCF in Mexico (Fig. 1). Previous studies of macrofungi, including Xylaria species, have been carried out throughout the Americas, mainly in Argentina, Colombia, Mexico, and the United States of America (Lodge et al., 2008; Hladki and Romero, 2010; Gómez-Hernández and Williams-Linera, 2011; Soto-Medina and Bolaños-Rojas, 2013; MacGregor-Fors et al., 2015, 2016; Tapia et al., 2017; Ávalos-Lázaro et al., 2018). Such studies vary widely in sampling efforts, collection methods, objectives, and levels of identification, making meaningful comparisons difficult. However, these studies also report that TMCF maintains a high level of Ascomycota-Xylariaceae diversity, which differs in composition and structure among sites within a same region.
The total number of species and/or morphospecies collected in this study represents nearly 29% of all Xylaria species reported for Mexico (San Martín and Rogers, 1989) and 65% of all Xylaria species recorded for the state of Veracruz (Medel et al., 2008) (Table 1). In addition, the inventory presented in this study increased with 59% the current number of Xylaria species reported for TMCF by Medel (2013). Ten percent of the collected species are new records for Mexico and the state of Veracruz (Tapia et al., 2017), while six percent are new locality records in the state of Veracruz. Furthermore, two of the collected morphospecies may be new species because they are supported by parsimony phylogenetic analysis of morphological characters (Chacón Zapata, com. pers.). These results demonstrate that the NPA of TMCF serves as an important refuge for Xylaria diversity and likely for other Ascomycota species, some of which appear to restrict their distributions to this type of vegetation (Tapia et al., 2017).
The historically recorded number of Xylaria species in the NPA of TMCF is significantly high compared to the findings of other studies carried out in the region (Chacón and Medel, 1993; Medel et al., 2008; Medel, 2013). Although a large proportion of species was present in each sampling period, not all of them were recorded in each period (completeness deficits <10%) (Fig. 3). This result suggests that a consistently longer sampling period over several seasons would be necessary to record most species present in this NPA. Some studies have reported the significance of sampling macrofungi, including Xylaria, over periods of several years to record the majority of species present at a site (Gómez-Hernández and Williams-Linera, 2011; MacGregor-Fors et al., 2015, 2016;). Specifically, upon evaluating the species diversity of Xylaria during 36 years in a NPA of TMCF, we observed deficits of 9% (1980-1989), 5% (2000-2009), 2% (1990-1999), and <0.1% (2010-2016) in the inventory completeness of the different sampling periods. These results indicate the difficulty of determining the number of years of collection that are required to represent the majority of Xylaria species in a given habitat. This implication was previously shown for other macrofungal assemblages associated with TMCF fragments in the region near the studied NPA (Gómez-Hernández and Williams-Linera, 2011). Given the current scarcity of knowledge of the mycobiota of TMCF and the findings of the present study, our results significantly contribute to the data on Xylaria assemblages for Mexico and the state of Veracruz. Thus, the alpha diversity of Xylaria assemblages can potentially be used as an indicator parameter of conservation degree of the TMCF (Figs. 2, 3).
In the comparison of compositional similarity, we found two significant clusters or effective communities, sensuJost (2006)), of Xylaria assemblages (Fig. 4). This result indicates that the composition of Xylaria assemblages differentiated over time. This pattern of differentiation in composition may be explained by species turnover between time periods, which may be related to the relatively high fraction of species that are specific to each time period and the low fraction of numerically dominant species (Fig. 5). This result is consistent with other studies that have investigated the beta diversity of macrofungal assemblages in the region (Gómez-Hernández and Williams-Linera, 2011; MacGregor-Fors et al., 2015, 2016). Therefore, temporal shifts in species turnover suggest that the compositional similarity of Xylaria assemblages, and likely of other macrofungi, may be highly dynamic because of the addition and disappearance of new and existing species, respectively, over time.
Although the sampling efforts were not identical in each sampling period, we estimated a reliable inventory over time, reinforcing the verity of the observed changes in the Xylaria assemblages at the study area during the last 36 years. The alpha and beta diversity of Xylaria assemblages are undoubtedly influenced by local factors (e.g., human activity, management of NPA, and natural forest regeneration) and landscape effects (e.g., composition and structure of land use/land cover types) (see Study Area, section at Material and Methods). In fact, we consider that changes in the Xylaria assemblages (i.e., gains in diversity and compositional shifts) are likely related to changes in land use/land cover or management of the study area (see Study Area section at Material and Methods). Given that such changes impact habitat structure or microenvironmental conditions, they may limit or, in this case, favor the ability of Xylaria species to colonize or increase their abundance inside the TMCF.
The human impacts resulting from the management of the NPA have been historically significant (see Study Area section in Material and Methods). Recently, since 2010, a maturation or regeneration process has likely been occurring in the protected TMCF, which is supported by several studies carried out in the area. For example, the same NPA was found to harbor an impressive array of macrofungi assemblages that are shaped by resources and conditions at the habitat level (Gómez-Hernández and Williams-Linera, 2011; MacGregor-Fors et al., 2016). Likely, the natural succession and the associated subsequent changes in substrate availability and microclimate, in the studied NPA, may be currently responsible for improving the quality of habitats available to Xylaria assemblages (see Study Area section at Material and Methods).
In conclusion, this study highlights the value of using Xylaria assemblages as a bioindicator group and as a means of monitoring the effectiveness of conservation strategies and the impact of management activities over time. Overall, our results suggest that the temporal dynamics of the assemblages partly depend on the conservation and management strategies that have been historically implemented in the studied NPA. The historical and structural heterogeneity recorded in the NPA may have induced shifts in the alpha and beta diversity of Xylaria assemblages, which, in turn, may have also influenced the ecological and ecosystem processes of the TMCF. Therefore, the present NPA is demonstrated to be an important habitat for Xylaria, an environmentally sensitive group of macrofungi. Effective conservation outcomes for TMCF will only be achieved if current management strategies are maintained and integrated into the long-term management framework of NPAs containing TMCF.

