











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los macrolíquenes son formas de crecimiento terrestres exitosas y diversas, están ampliamente distribuidas desde los polos al ecuador (Barreno Rodríguez y Pérez-Ortega, 2003; Santoni y Lijteroff, 2006; Fuga et al., 2008). Se estima que de 8 a 10% de los ecosistemas terrestres están dominados por líquenes, contribuyendo de manera importante a la biomasa, la biodiversidad, los ciclos de nutrimentos y de minerales, y al flujo energético (Seaward, 2008). Promueven la formación de suelo, incrementan la disponibilidad de nitrógeno en el sustrato y acumulan varios elementos esenciales, como K, P y S (Seaward, 2008). Son capaces de fijar nitrógeno atmosférico aportando hasta 25% en el ciclo local de este elemento; alteran las características de la corteza de los árboles e interfieren en las interacciones entre parásitos y plantas hospederas (Forman, 1975; Bermúdez de Castro et al., 1990).
Debido a su sensibilidad a cambios abruptos en sus hábitats así como a la contaminación atmosférica, los líquenes son usados en el monitoreo de contaminación ambiental (Conti y Cecchetti, 2001). Los factores ecológicos, como el sustrato (composición química y pH) y el clima, condicionan la naturaleza del complejo liquénico al igual que su distribución. Las rocas o los suelos silíceos albergan floras muy distintas de aquellos sustratos carbonatados o evaporíticos, debido al color, textura, porosidad, permeabilidad y a la disponibilidad del agua. Los sustratos oscuros se calientan mucho más que los claros y ello incide sobre la temperatura, la retención de agua de los talos liquénicos y en el cociente de fotosíntesis/respiración (Brodo et al., 2001; Herrera-Campos y Nash III, 2001; Gómez-Peralta, 2003; Barreno Rodríguez y Pérez-Ortega, 2003; Herrera-Campos et al., 2014).
Los líquenes consisten en una asociación mutualista formada por organismos de dos o tres diferentes reinos biológicos. Están constituidos por un hongo (micobionte) y uno o dos autótrofos algales y/o cianobacteriales (fotobiontes) (Scott, 1973; Hale, 1983; Honegger, 1998; Honegger y Scherrer, 2008; Lijteroff et al., 2009). El número de especies conocidas de líquenes es de 19,387, perteneciendo la mayoría al phylum Ascomycota (19,215) y 0.9% a Basidiomycota (172) (Barreno Rodríguez y Pérez-Ortega, 2003; Oksanen, 2006; Riquelme, 2008; Lücking et al., 2016). En los líquenes u hongos liquenizados, el micobionte constituye de 80 a 90% de la biomasa del talo, mientras que, en algunos géneros de líquenes tropicales, el fotobionte es dominante (Nash III et al., 2002). Se clasifican en el reino Eumycota, phylum Ascomycota, subphylum Pezizomycotina, clases Arthoniomycetes, Coniocybomycetes, Dothideomycetes, Eurotiomycetes, Lecanoromycetes, Lichinomycetes, Sordariomycetes y en el phylum Basidiomycota, subphylum Agaricomycotina, clase Agaricomycetes (Lücking et al., 2016).
El estudio de la diversidad liquénica de México es relativamente reciente (Herrera-Campos y Nash III, 2001). En México, se han registrado 2833 taxones de líquenes, pertenecientes a 2722 especies y 111 categorías infra-específicas y se estima que representan aproximadamente 50% total de especies esperadas para el país. La región centro-norte (Aguascalientes, Guanajuato, Querétaro, San Luis Potosí y Zacatecas) tiene el menor número de especies reportadas (216) (Herrera-Campos et al., 2014).
Son pocos los trabajos florísticos acerca de los líquenes del estado de Guanajuato, entre los que se encuentran el realizado por Puy-Alquiza et al. (2015), quienes describen el papel de las comunidades de líquenes en el deterioro superficial de su sustrato rocoso en los edificios históricos de la ciudad de Guanajuato, y el de Gaytán-Esparza y León-Álvarez (2013) en donde realizan una caracterización genética y molecular de líquenes tolerantes a contaminación ambiental en muestras de la Sierra de Lobos, Sierra de Santa Rosa, Irapuato, comunidad Joya del Calvillo y cerro de Arandas.
Con base en la literatura revisada, se puede decir que es escasa la información que existe acerca de la flora liquénica de las zonas áridas de la altiplanicie mexicana y más aún sobre las especies saxícolas. Por lo anterior, el objetivo de la presente contribución fue la elaboración de la lista florística de los macrolíquenes saxícolas de cuatro localidades del municipio de Guanajuato cercanas a la ciudad del mismo nombre: cerro “La Bufa” (CB), parque ecológico “El Orito” (PEO), carretera panorámica “El Pípila” (CPP) y Sierra de Santa Rosa de Lima (SRL).
Materiales y Métodos
Área de estudio
El distrito minero de Guanajuato es una de las regiones mineras más importantes del país, pertenece a una entidad geológica definida como Sierra de Guanajuato, de características geológicas singulares. Su riqueza minera se debe a la presencia de tres zonas productoras de plata y oro denominadas: Vetas de la Sierra, Veta Madre y Vetas de la Luz. La Veta Madre es la zona más importante y se encuentra en la ciudad de Guanajuato. Esta última ciudad dio nombre al distrito y se convirtió en la capital del estado, así como un importante centro turístico internacional, gracias a sus riquezas mineras y su relevancia histórica. La producción minera del distrito supera 37,000 toneladas métricas de plata refinada y 135 de oro. Con ello, el distrito es el segundo en importancia mundial, sólo después del de Real del Monte-Pachuca (Antúnez, 1964).
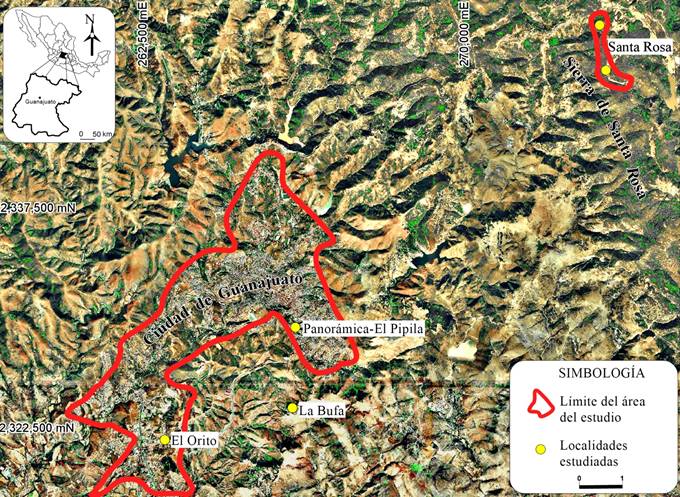
En las inmediaciones del distrito minero se estudiaron cuatro localidades: 1) cerro “La Bufa”, coordenadas 14Q0265971, 2323270, a 2600 m de altitud. Caracterizada por la presencia de líquenes en depósitos ignimbríticos de edad Oligoceno (IFB), en rocas sedimentarias, específicamente areniscas de la Formación Losero (AFL) y en el Conglomerado Guanajuato (CG); 2) parque ecológico “El Orito”, coordenadas 14Q0263071, 2322610, a 2021 m de altitud. Los líquenes se encuentran en rocas sedimentarias principalmente en el Conglomerado Guanajuato (CG); 3) carretera panorámica “El Pípila”, coordenadas 14Q0265829, 2325073, a 2106 m de altitud. Los líquenes se encuentran igual que en la localidad 2, en el Conglomerado Guanajuato (CG); 4) Sierra de Santa Rosa de Lima, ubicada en las coordenadas 14Q0272808, 2332042, a una altitud de 2113 m. Los líquenes se encuentran en roca volcánica (toba) de composición riolítica (TR) (Fig. 1).
Clima
En el distrito minero de Guanajuato, en donde se ubican las muestras de la localidad 1, 2 y 3, predomina un clima templado-subhúmedo de acuerdo con la clasificación de Köppen modificada por García (1964). Alcanza una temperatura máxima de 36 °C en verano y una mínima de 3 °C en invierno, dándose una temperatura media anual de 18.5 °C. A su vez, la precipitación pluvial varía de 600 a 840 mm anuales, siendo el promedio anual de 697 mm. En los meses de julio y agosto se observa una mayor incidencia de lluvias, alcanzando un valor de 217 mm cada uno, distinguiéndose como el mes más seco febrero, con un valor menor de 10 mm. La irregularidad del relieve determina que en las partes elevadas la temperatura disminuya a menos de 18 °C y en las partes bajas llegue a 22 °C. Los vientos dominantes provienen del oeste en invierno, en primavera del sureste y suroeste, y durante verano-otoño del este y noreste.
El clima en la Sierra de Santa Rosa de Lima, localidad 4, es cálido a templado. Los veranos son lluviosos con una precipitación anual de 810 mm, con una temperatura promedio de 14.9 °C (García, 1964).
Recolección de líquenes saxícolas
Se seleccionaron cuatro localidades de muestreo correspondientes a las áreas del distrito minero de Guanajuato y la Sierra de Santa Rosa de Lima (Cuadro). En cada localidad se caracterizaron los diferentes tipos de roca, en donde los líquenes fueron recolectados. La mayoría de los ejemplares (42 en total), con excepción de los fruticosos fueron recolectados con ayuda de un martillo de geólogo y cincel; durante los meses de noviembre y diciembre de 2014. Los ejemplares fueron envueltos en sobres de papel, recolectándose preferentemente los líquenes que presentaron estructuras reproductoras visibles (apotecios, peritecios, picnidios, soredios e isidios, principalmente). Se tomaron fotografías de los ejemplares en campo con una cámara Canon SX420 (Modelo 1068C001, Los Ángeles, EUA).
Cuadro Listado de especies, sustrato y localidad. Simbología de sustrato: AFL (Areniscas de la Formación Losero), CG (Conglomerado Guanajuato), IFB (Ignimbrita Formación “La Bufa”), TR (Toba riolítica). Simbología de localidad: CPP (carretera panorámica “El Pípila”), CB (cerro La Bufa), PEO (parque ecológico “El Orito”), SRL (Sierra de Santa Rosa de Lima). *Nuevo registro para Guanajuato, ***Nuevo registro para México, ▲ Especies que se fijan al sustrato por un ombligo (Umbilicados). Todas las colectas fueron realizadas por Violeta Cortés-Hernández.
| Forma biológica | Taxa | Sustrato | Localidad |
|---|---|---|---|
| Foliosa | Lecanorales | ||
| Parmeliaceae | |||
| Canoparmelia texana (Tuck.) Elix & Hale 298 (EBUM 27495), 349 (EBUM 27496) | IFB | CB | |
| Flavoparmelia caperata (L.) Hale 366 (EBUM 27513) | TR | SRL | |
| Flavopunctelia flaventior (Stirt.) Hale 290 (EBUM 27514), 300 (EBUM 27515), 348 (EBUM 27516), 410 (EBUM 27517) | IFB | CB | |
| F. praesignis (Nyl.) Hale 365 (EBUM 27518) | TR | SRL | |
| *Hypotrachyna subsaxatilis (B. de Lesd.) Hale 308 (EBUM 27522) | IFB | CB | |
| Parmotrema cetratum (Ach.) Hale 361 (EBUM 27551), 407 (EBUM 27552) | TR | SRL | |
| P. crinitum (Ach.) M. Choisy 371 (EBUM 27527) | TR | SRL | |
| P. eurysacum (Hue) Hale 302 (EBUM 27528) | IFB, TR | CB, SRL | |
| P. reticulatum (Taylor) M. Choisy 402 (EBUM 27553), 406 (EBUM 27554), 430 (EBUM 27555) | CG, TR | PEO, SRL | |
| *Punctelia punctilla (Hale) Krog 327 (EBUM 27547), 341 (EBUM 27548), 385 (EBUM 27549) | IFB, CG | CB, SRL | |
| Xanthoparmelia coloradoensis (Gyeln.) Hale 305 (EBUM 27569) | IFB | CB | |
| *X. conspersa (Ehrh. ex Ach.) Hale 425 (EBUM 27570) | IFB | CB | |
| X. lavicola (Gyeln.) Hale 426 (EBUM 27571) | CG | CPP | |
| X. mexicana (Gyeln.) Hale 96 (EBUM 27581), 451 (EBUM 27582) | IFB | CB | |
| *X. nigropsoromifera (T.H. Nash) Egan 303 (EBUM 27583) | IFB | CB | |
| * , ***X. paradoxa Hale 292 (EBUM 27584) | IFB | CB | |
| X. rimalis (Kurok.) Elix, A. Thell & Søchting 294 (EBUM 27572), 299a (EBUM 27573), 301 (EBUM 27574), 343 (EBUM 27575), 369 (EBUM 27576), 409(EBUM 27577), 442 (EBUM 27578), 445 (EBUM 27579), 455 (EBUM 27580) | IFB, CG, TR | CB, PEO, SRL | |
| X. subtasmanica Elix & T.H. Nash 408 (EBUM 27585) | IFB | CB | |
| X. tinctina (Maheu & A. Gillet) Hale 427 (EBUM 27565), 439 (EBUM 27566), 446 (EBUM 275667), 450 (EBUM 27568) | IFB, AFL, CG | CB, PEO, CPP | |
| Caliciales | |||
| Physciaceae | |||
| Heterodermia albicans (Pers.) Swinscow & Krog 434 (EBUM 27520), 438 (EBUM 27521) | CG, DP | PEO | |
| *Phaeophyscia adiastola (Essl.) Essl. 353 (EBUM 27533) | CG | CPP | |
| *P. ciliata (Hoffm.) Moberg 437 (EBUM 27534) | CG, DP | PEO | |
| Physcia aipolia (Ehrh. ex Humb.) Fürnr. 448 (EBUM 27535) | CG | CB | |
| *P. convexa Müll. Arg. 329 (EBUM 27536) | AFL | CB | |
| *P. halei J.W. Thomson 390 (EBUM 27538), 404 (EBUM 27539) | TR | SRL | |
| *P. millegrana Degel. 318 (EBUM 27540), 386 (EBUM 27541), 423 (EBUM 27542) | IFB, TR | CB, SRL | |
| *P. tribacia (Ach.) Nyl. var. tribacia Nyl. 453 (EBUM 27537) | CG | CPP | |
| Peltigerales | |||
| Collemataceae | |||
| *Physconia thomsonii Essl. 363 (EBUM 27543), 432 (EBUM 27544), 436 (EBUM 27545) | CG | CPP, PEO | |
| *Collema tenax (Sw.) Ach. var. tenax 468 (EBUM 27507) | CG | CPP | |
| *Enchylium coccophorum (Tuck.) Otálora, P.M. Jørg. & Wedin 428 (EBUM 27503), 466 (EBUM 27504), 467 (EBUM 27505) | CG IFB, DP | CPP, PEO | |
| *E. polycarpon (Hoffm.) Otálora, P.M. Jørg. & Wedin 322 (EBUM 27506) | CG | CB | |
| *Leptogium wilsonii Zahlbr. 469 (EBUM 27526) | CG, DP | PEO | |
| Peltigerales | |||
| Coccocarpiaceae | |||
| Coccocarpia palmicola (Spreng.) Arv. & D.J. Galloway 324 (EBUM 27500), 370 (EBUM 27501), 447 (EBUM 27502) | IFB, CG, TR | CB, SRL | |
| Umbilicariales | |||
| Umbilicariaceae | |||
| *▲ Lasallia papulosa (Ach.) Llano 368a (EBUM 27523) | TR | SRL | |
| *▲ L. pustulata (L.) Mérat 444 (EBUM 27524) | TR | SRL | |
| Verrucariales | |||
| Verrucariaceae | |||
| *▲ Dermatocarpon miniatum (L.) W. Mann var. miniatum 319 (EBUM 27508), 378 (EBUM 27509), 431 (EBUM 27510), 441 (EBUM 27511) | IFB, TR, CG, DP | CB, SRL, PEO | |
| Fruticosa | Lecanorales | ||
| Cladoniaceae | |||
| *Cladonia asahinae J.W. Thomson 463 (EBUM 27497) | TR | SRL | |
| *C. scabriuscula (Delise) Leight. 464 (EBUM 27498) | TR | SRL | |
| Stereocaulaceae | |||
| *Lepraria albicans (Th. Fr.) Lendemer & B.P. Hodk. 384 (EBUM 27525) | TR | SRL | |
| Ramalinaceae | |||
| *Ramalina intermedia (Delise ex Nyl.) Nyl. 449 (EBUM 27550) | TR | SRL | |
| Parmeliaceae | |||
| Usnea amblyoclada Müll. Arg. 338 (EBUM 27556), 388 (EBUM 27557), 393 (EBUM 27558), 394 (EBUM 27559), 395 (EBUM 27560), 397(EBUM 27561) | IFB, TR | CB, SRL | |
| *U. halei P. Clerc 461 (EBUM 27562), 462 (EBUM 27563) | TR | SRL |
Determinación de especies
Las especies fueron determinadas considerando sus características morfológicas, anatómicas y químicas. Para ello se aplicaron varias técnicas: microscopio estereoscópico (marca Zeiss Stemi DV4, Nueva York, EUA), microscopio compuesto (marca Zeiss Stemi DV4, Nueva York, EUA), y en algunos casos cromatografía y/o a la respuesta bajo luz ultravioleta λ 254 nm.
Para cada especie, en un microscopio estereoscópico, se observó: 1) la presencia o ausencia de estructuras reproductoras (sexuales y asexuales), 2) ornamentación de la superficie superior e inferior y estructuras de fijación, 3) la respuesta al contacto con los reactivos químicos: hidróxido de potasio al 10% en agua (K), hipoclorito de sodio (C), parafenilendiamina en alcohol etílico (P), y 4) a la combinación de los reactivos K y C para algunas estructuras del talo (corteza superior y médula) así como del apotecio.
Para la observación en el microscopio compuesto se realizaron cortes transversales semifinos de las estructuras reproductoras sexuales y del talo y en algunos casos también de picnidios los cuales fueron montados en agua. Para observar las características de los tejidos del apotecio, ascas, esporas y reacciones, fue necesario agregar una gota de hidróxido de potasio al 10% en agua (K) o solución de lugol al 3% (KI). Se consideraron las medidas de más de cinco ascas y más de diez esporas (100×).
Para corroborar la determinación de algunas especies se realizó la cromatografía de capa fina o TLC (Thin Layer Chromatography), siguiendo la técnica propuesta por Lumbsch (2002) y utilizando los solventes A, B y C. Para confirmar la identidad de los ácidos liquénicos se utilizó el catálogo de datos cromatográficos estandarizados de Elix (2014), que se basa en valores de RF (relación entre las distancias recorridas por el soluto y por el eluyente desde el origen de la placa), en los sistemas de solventes utilizados, el color de la mancha visible de manera natural o bajo luz ultravioleta, color visible y bajo luz ultravioleta después del rociado con H2SO4, entre otros.
Se consultaron claves taxonómicas para identificar formas biológicas (Thomson, 1963; Brodo et al., 2001; Nash III et al., 2002; Nash III et al., 2004a; Nash III et al., 2007, y Lendemer y Hodkinson, 2010). Finalmente, se revisaron descripciones de las mismas y se cotejaron los ejemplares determinados (42 en total) con aquellos existentes en el herbario EBUM de la Universidad Michoacana de San Nicolás de Hidalgo, donde fue depositado el material revisado.
Se realizó una revisión de la literatura especializada, así como de los nuevos registros, tanto para la localidad estudiada como para el país, con la finalidad de elaborar una lista de especies de acuerdo al arreglo sistemático propuesto por Lücking et al. (2016). Para la distribución de las especies se siguió lo indicado en el Consortium of North American Lichen Herbaria (CNALH) y se consultó información adicional para algunas especies en el Global Information System for Lichenized and Non-Lichenized Ascomycetes (LIAS, 2017) y de diversos trabajos publicados. La nomenclatura y abreviatura de los autores de las especies fue corroborada con Index Fungorum (2017).
Resultados
En el Cuadro se resumen los hallazgos de este estudio sobre la diversidad de especies de macrolíquenes saxícolas, el tipo de sustrato rocoso, las localidades estudiadas, el número de recolecta y entre paréntesis, el folio asignado en el herbario EBUM (las recolectas fueron realizadas por Violeta Cortés-Hernández, la última autora de este trabajo, por lo cual su nombre será obviado en este apartado).
Se identificaron 42 especies en nueve familias, cuatro de estas (Parmeliaceae, Cladoniaceae, Ramalinaceae y Sterocaulaceae) corresponden al orden Lecanorales; dos (Collemataceae y Coccocarpiaceae) a Peltigerales, mientras que una (Physciaceae) a Caliciales, y Umbilicariaceae y Verrucariaceae a los órdenes Umbilicariales y Verriucariales, respectivamente, de acuerdo con el arreglo de Lücking et al. (2016).
Se determinaron para los líquenes foliosos 36 especies en seis familias (Coccocarpiaceae, Collemataceae, Parmeliaceae, Umbilicariaceae, Verrucariaceae y Physciaceae), y para los fruticosos seis especies en cuatro familias (Cladoniaceae, Parmeliaceae, Ramalinaceae y Stereocaulaceae).
Todas las especies son nuevos registros para las localidades estudiadas y 23 lo son para Guanajuato; una es nuevo registro para México: Xanthoparmelia paradoxa Hale, registrada para África, en la provincia del Cabo (Hale, 1990); su distribución incluye Madagascar (LIAS, 2017).
De la familia Parmeliaceae, los nuevos registros para el estado (Cuadro y Fig. 2) son: Hypotrachyna subsaxatilis (B. de Lesd.) Hale, registrada para Michoacán (Hale, 1975), Chihuahua y Sinaloa (Nash III y Elix, 2002), Durango, Estado de México, Guerrero, Hidalgo, Jalisco, Oaxaca y Sonora (Nash III et al., 2016b); Punctelia punctilla (Hale) Krog, para Baja California Sur (Egan y Aptroot, 2004) y Baja California (Egan y Lendemer, 2016); Xanthoparmelia conspersa (Ehrh. ex Ach.) Hale, para Baja California, Chihuahua, Estado de México, Durango, Hidalgo, Jalisco, Michoacán, Sonora, Oaxaca, Veracruz (Nash III y Elix, 2004; Nash III et al., 2004a) y Puebla (Nash III et al., 2016a); X. nigropsoromifera (T.H. Nash) Egan, para Chihuahua, Durango, Sonora, Zacatecas (Nash III y Elix, 2004; Nash III et al., 2004a, 2004b; Nash III et al., 2016a) y Usnea halei P. Clerc, para Chihuahua y Sinaloa (Clerc, 2007), Chihuahua, Durango (Clerc y Herrera-Campos, 1997), Hidalgo y Tamaulipas (Herrera-Campos, 2016). De la familia Physciaceae (Fig. 3): Phaeophyscia adiastola (Essl.) Essl., registrada para Baja California Sur (CNALH, 2017); P. ciliata (Hoffm.) Moberg, para Sonora (Esslinger, 2004), Baja California, Puebla y Sonora (CNALH, 2017); Physcia convexa Müll. Arg., para Baja California Sur, Chihuahua y Sonora (Moberg, 2002); P. halei J.W. Thomson, para Baja California, Baja California Sur, Chihuahua y Sonora (Moberg, 2002); P. millegrana Degel; P. tribacia (Ach.) Nyl. var. tribacia, para Baja California, Baja California Sur, Chihuahua y Sonora (Moberg, 2002) y Physconia thomsonii Essl., solo para Michoacán (Gregorio-Cipriano et al., 2016). De la familia Collemataceae (Fig. 3): Enchylium coccophorum (Tuck.) Otálora, P.M. Jørg. & Wedin, registrada para Baja California, Baja California Sur, Chihuahua y Sonora (Schultz et al., 2004), Coahuila, Durango y Nuevo León (CNALH, 2017); E. polycarpon (Hoffm.) Otálora, P.M. Jørg. & Wedin, para Chihuahua (Schultz et al., 2004), además de Coahuila y Sonora (CNALH, 2017); Collema tenax (Sw.) Ach. var. tenax, para Baja California, Baja California Sur, Chihuahua, Coahuila y Sonora (Schultz et al., 2004); Leptogium wilsonii Zahlbr, para Baja California Sur, Chihuahua, Sinaloa y Sonora (Jørgensen y Nash, 2004), además de Colima, Durango, Hidalgo y Michoacán (CNALH, 2017). Del resto de las familias (Fig. 3): Cladonia asahinae J.W. Thomson, cuya distribución incluye México, sin indicar la región (LIAS, 2017); C. scabriuscula (Delise) Leight, registrada para Baja California (Ahti y Hammer, 2002); Ramalina intermedia (Delise ex Nyl.) Nyl., para Chihuahua(Kashiwadani y Nash III, 2004); Lepraria albicans (Th. Fr.) Lendemer & B.P. Hodk., para Chihuahua (Ryan, 2002) e Hidalgo (CNALH, 2017), (Cuadro). Lasallia papulosa (Ach.) Llano, para Jalisco y Michoacán (Gómez-Peralta, 1992; 2003), Chihuahua (Sancho, 2004), Baja California, Baja California Sur, Jalisco, Oaxaca, Sinaloa (CNALH, 2017) y Puebla (Brizuela y Guzmán, 1971); L. pustulata (L.) Mérat, para Michoacán (Bouly de Lesdain, 1914; 1922); Dermatocarpon miniatum (L.) W. Mann var. miniatum, para Michoacán (Bouly de Lesdain, 1929), Baja California, Baja California Sur, Chihuahua, Sonora, Oaxaca, Tlaxcala y Veracruz (CNALH, 2017).

Figura 2: Macrolíquenes de la familia Parmeliaceae: A. Canoparmelia texana (Tuck.) Elix & Hale (barra=3 mm); B. Flavoparmelia caperata (L.) Hale (barra=3 mm); C. Flavopunctelia flaventior (Stirt.) Hale (barra=2 mm); D. F. praesignis (Nyl.) Hale (barra=2 mm); E. detalle de apotecios en F. praesignis (barra=0.5 mm); F. Hypotrachyna subsaxatilis (B. de Lesd.) Hale (barra=2 mm); G. Parmotrema cetratum (Ach.) Hale (barra=1 mm); H. detalle de apotecio en P. cetratum, (barra=1mm); I. P. crinitum (Ach.) M. Choisy (barra=2 mm); J. P. eurysacum (Hue) Hale (barra=2 mm); K. detalle de apotecio en P. eurysacum (barra=0.5 mm); L. P. reticulatum (Taylor) M. Choisy (barra=3 mm); M. Punctelia punctilla (Hale) Krog (barra=2 mm); N. Usnea amblyoclada (Müll. Arg.) Zahlbr. (barra=5 mm); Ñ. detalle de isidiomorfos con la punta pigmentada de negro (barra=2 mm); O. U. halei P. Clerc (barra=5 mm); P. Xanthoparmelia coloradoensis (Gyeln.) Hale (barra=3 mm); Q. X. conspersa (Ehrh. Ex Ach.) Hale (barra=2 mm); R. X. lavicola (Gyeln.) Hale (barra=2 mm); S. X. mexicana (Gyeln.) Hale (barra=2 mm); T. X. nigropsoromifera (T.H. Nash) Egan (barra=3 mm); U. X. paradoxa Hale (barra=3mm); V. X. rimalis (Kurok.) Elix, A. Thell & Søchting (barra=4 mm); W. X. subtasmanica Elix & T.H. Nash (barra=3 mm); X. X. tinctina (Maheu & A. Gillet) Hale (barra=4 mm).

Figura 3: Macrolíquenes de otras familias: A. Cladonia asahinae J.W. Thomson (barra=3 mm); B. C. scabriuscula (Delise) Leight (barra=3 mm); C. Coccocarpia palmicola (Spreng.) Arv. & D.J. Galloway (barra=2 mm); D. Collema tenax (Sw.) Ach. var. tenax (barra=2 mm); E. Dermatocarpon miniatum (L.) W. Mannvar. miniatum (barra=4 mm); F. Enchylium coccophorum (Tuck.) Otálora, P.M. Jørg. & Wedin (barra=2 mm); G. E. polycarpon (Hoffm.) Otálora, P.M. Jørg. & Wedin (barra=3 mm); H. Heterodermia albicans (Pers.) Swinscow & Krog (barra=3 mm); I. Lasallia papulosa (Ach.) Llano (barra=5 mm); J. L. pustulata (L.) Mérat, (barra=5 mm); K. Lepraria albicans (Th. Fr.) Lendemer & B.P. Hodk (barra=2 mm); L. Leptogium wilsonii Zahlbr. (barra=3 mm); M. Phaeophyscia adiastola (Essl.) Essl. (barra=2 mm); N. detalle de soredia granular e isidioide de P. adiastola (barra=2 mm); Ñ. P. ciliata (Hoffm.) Moberg (barra=2 mm); O. detalle de cilios en P. ciliata (barra=1 mm); P. Physcia aipolia (Ehrh. Ex Humb.) Fürnr. (barra=3 mm); Q. P. convexa Müll. Arg. (barra=3 mm); R. detalle de puntas de los lóbulos y superficie maculada en P. convexa (barra=3 mm); S. P. halei J.W. Thomson (barra=2 mm); T. P. millegrana Degel. (barra=2 mm); U. Physconia thomsonii Essl. (barra=2 mm); V. detalle de soralia y pruina en P. thomsonii (barra=2 mm); W. P. tribacia (Ach.) Nyl. var. tribacia (barra=2 mm); X. Ramalina intermedia (Delise ex Nyl.) Nyl. (barra=3 mm).
Las especies Canoparmelia texana (Tuck.) Elix & Hale, Flavoparmelia caperata (L.) Hale, Flavopunctelia flaventior (Stirt.) Hale, F. praesignis (Nyl.) Hale, Parmotrema cetratum (Ach.) Hale, P. crinitum (Ach.) M. Choisy, P. eurysacum (Hue) Hale, P. reticulatum (Taylor) M. Choisy, Usnea amblyoclada (Müll. Arg.) Zahlbr., Xanthoparmelia coloradoensis (Gyeln.) Hale, X. lavicola (Gyeln.) Hale, X. mexicana (Gyeln.) Hale,X. paradoxa, X. rimalis (Kurok.) Elix, A. Thell & Søchting, X. subtasmanica Elix & T.H. Nash, X. tinctina (Maheu & A. Gillet) Hale, Coccocarpia palmicola (Spreng.) Arv. & D.J. Galloway, Heterodermia albicans (Pers.) Swinscow & Krog y Physcia aipolia (Ehrh. Ex Humb.) Fürnr. ya estaban registrados para el estado de Guanajuato (Figs. 2, 3).
Discusión
Se registran por primera vez para el distrito minero de Guanajuato 42 especies de macrolíquenes saxícolas, los cuales se encuentran representados por las familias Parmeliaceae (21 especies), Physciaceae (nueve), Collemataceae (cuatro), Cladoniaceae y Umbilicariaceae (dos cada una), Coccocarpiaceae, Ramalinaceae, Stereocaulaceae y Verrucariaceae (una cada una). De estas especies 57.14% son nuevos registros para el estado, lo que constituye una importante contribución al conocimiento de la diversidad liquénica de la región centro-norte del país, la cual de acuerdo con Herrera-Campos et al. (2014) es una de las zonas con menor número de especies de líquenes registradas.
De acuerdo con los sustratos rocosos presentes en el área estudiada, la toba riolítica presenta una mayor diversidad de especies (19), siendo las más representativas las correspondientes a Parmeliaceae. Las tobas tienen como peculiaridad su origen volcánico, color claro, grano fino, composición ácida, con alta porosidad y bajo escurrimiento, facilitando el almacenamiento de agua. En cuanto al sustrato de origen ignimbrítico se contabilizaron 18 especies, de las cuales 14 corresponden a Parmeliaceae, destacando en ello la presencia de ocho de las nueve especies de Xanthoparmelia del área de estudio. Al igual que la toba riolítica son rocas volcánicas cuya diferencia con esta última es el tamaño de grano. El conglomerado Guanajuato presenta una menor diversidad de macrolíquenes (17) en donde Collemataceae, Physciaceae y Verrucariaceae dominan en dicho sustrato, caracterizado éste por ser una roca de origen sedimentario de grano grueso a fino, color claro, poca dureza, composición alcalina, alta porosidad y escurrimiento bajo, lo que favorece la colonización de dichos especímenes. De los cinco sustratos rocosos, la toba riolítica, la ignimbrita y el conglomerado Guanajuato presentan la mayor diversidad de especies. Con base en las localidades estudiadas la zona que presenta mayor número de especies es el cerro “La Bufa”, debido a la presencia de una mayor cantidad de sustratos rocosos (tres de los cinco que se presentan en el área), tanto de origen sedimentario como volcánico.
Es importante mencionar que esta investigación contribuye a ampliar el conocimiento de la flora liquénica para la región centro-norte del país. De igual manera dicha contribución apoyará a investigaciones futuras encaminadas al monitoreo de contaminación ambiental utilizando para ello las especies que se desarrollan en la zona, de igual manera el conocimiento de la existencia de dichas especies permitirá conocer cómo éstas intervienen en el deterioro de los monumentos considerados como patrimonio cultural.
Conclusión
De los 42 macrolíquenes que se presentan en este trabajo, Parmeliaceae es la más representativa del área de estudio, especialmente el género Xanthoparmelia con nueve especies, de las cuales dos (X. conspersa y X. nigropsoromifera) son nuevos registros para el estado de Guanajuato, mientras que Xanthoparmelia paradoxa lo es para México. La zona de La Bufa con clima semicálido y la ignimbrita como roca dominante reúne la mayoría de los macrolíquenes registrados. Cladoniaceae, Umbilicariaceae, Ramalinaceae y Stereocaulaceae son representativas de la zona de Santa Rosa de Lima en donde domina un clima húmedo y la toba riolítica como roca dominante. La diversidad de macrolíquenes encontrados en estas localidades se debe a las características texturales del sustrato rocoso que colonizan (color, forma, tamaño de grano, selección) y porosidad.
Contribución de autores: MJP, MGP, MGC y VRZ concibieron y diseñaron el estudio. MGP y MJP realizaron los análisis. MJP, MGP, RMA, DNR y VCH contribuyeron a la adquisición de datos y la interpretación. MJP escribió el manuscrito con la ayuda de MGP y MGC. Todos los autores contribuyeron a la discusión, revisión y aprobación del manuscrito final.
Financiamiento: Este estudio fue apoyado por la DINPO-Universidad de Guanajuato, Proyecto 358/2014.

