











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Tradicionalmente, la familia Leguminosae se ha dividido en tres subfamilias: Caesalpinioideae, Mimosoideae y Papilionoideae (Lewis et al., 2005). Una nueva clasificación de esta familia, basada en datos moleculares y morfológicos (LPWG, 2017) propone seis subfamilias: Duparquetioideae, Cercidoideae, Detarioideae, Dialioideae, Caesalpinoideae (incluyendo a la subfamilia Mimosoideae como sinónimo y formando un clado mimosoide anidado entre las tribus Caesalpinieae y Cassieae), y Papilionoideae; por lo que, actualmente, el género Mimosa L. podría ubicarse en la subfamilia Caesalpinoideae. Sin embargo, esta nueva clasificación no afecta la clasificación infragenérica de Mimosa ni los resultados de este trabajo.
En México, Mimosa cuenta con ca. 104 especies, de las cuales 54% es endémico (Grether et al., 2015). De acuerdo con Barneby (1991) y Grether (1997) los taxa se distinguen, a nivel infragenérico, utilizando caracteres morfológicos como la forma de vida, tipo de inflorescencia, morfología de la flor (i.e. número de estambres; presencia o ausencia de estaminodios), hoja (i.e. número de pinnas, número de folíolos), fruto (i.e. valvas enteras o en artejos, número de semillas por fruto), semilla (i.e. color, tamaño del pleurograma) y aguijones (i.e. posición y número), entre otros. No obstante, la delimitación de secciones y series aún es compleja, por lo que se ha implementado la búsqueda de otros caracteres que aporten información adicional para el reconocimiento entre taxa.
Con esta visión, se han realizado estudios palinológicos (Caccavari, 1985, 1986, 1987; Chehaibar, 1988; Flores-Cruz et al., 2006) y de la anatomía de la madera (Marchiori, 1996; Montaño-Arias, 2010; Montaño-Arias et al., 2016, 2017). Sin embargo, dado el número de taxa comprendidos en el género Mimosa, los estudios con enfoque taxonómico se consideran escasos.
Igualmente, es relevante mencionar que los caracteres de las células epidérmicas, y los estomas y tricomas han sido usados en la taxonomía de las angiospermas (Stuessy, 1990); así como la anatomía de los complejos estomáticos (Rosati, 1991). En particular, el estudio anatómico de los complejos estomáticos comenzó en la década de 1960 con la observación de una tendencia de las células subsidiarias a formar patrones constantes en un mismo taxon (Fahn, 1967; Roth y Clausnitzer, 1969; Percy y Santa, 1976). Desde entonces, se han realizado estudios enfocados a la determinación de estos patrones en diferentes taxa.
El tipo de complejo estomático ha permitido diferenciar los taxa de las familias Smilacaceae (Baruah et al., 2012), Cactaceae (Eggli, 1984) y Gesneriaceae (Ramírez-Roa y Varela-Hernández, 2011), entre otras.
En el caso de las leguminosas, Percy y Santa (1976) reportaron el tipo de complejo estomático de 300 especies pertenecientes a diversos géneros como Inga Mill., Swartzia Schreb., Trifolium L., Hymenaea L., Canavalia Adans., Dioclea Kunth y Tamarindus L., entre otros. Asimismo, Araujo (2005) reporta que seis especies del género Erythrina L. han podido diferenciarse por la ubicación del complejo estomático presente.
Por su parte, Grosso et al. (1994) estudiaron el tipo de complejo estomático en 102 especies de Acacia Mill., reconociendo dos tipos de complejos, el anomocítico y el anisocítico. En el caso particular del género Mimosa, se conoce que sus taxa pueden presentar folíolos hipostomáticos o anfistomáticos, que la presencia de complejos estomáticos de tipo paracítico es común, pero pueden llegar a presentar más de un tipo de complejo (Bässler, 1985; Chiamolera et al., 2011; Begum y Borthakur, 2013; Rothe y Bathe, 2014). Sin embargo, la anatomía foliar es un campo poco explorado. Con base en lo anterior, el objetivo de este estudio fue determinar, con fines taxonómicos, el tipo de complejo estomático de 24 taxa del género Mimosa (Leguminosae) presentes en México, bajo el supuesto de que, al aumentar el número de taxa analizados, habrá variación del tipo de complejos estomáticos, lo que permitirá la distinción de los taxa.
Material y Métodos
Material vegetal
Se seleccionaron 24 taxa de Mimosa presentes en México, los cuales presentaron forma de vida arbórea (un solo tronco leñoso, con ramificación a partir del metro de altura) o arbustiva (al menos dos troncos leñosos, con ramificación desde la base); dichos taxa pertenecen a diferentes secciones y series del género (Cuadro 1).
Cuadro 1: Clasificación de los taxa de Mimosa L. (Leguminosae) estudiados según Barneby (1991) y Grether (2000).
| Sección | Serie | Subserie | Especie | Variedad |
| Batocaulon | Acanthocarpae | M. aculeaticarpa Ortega | aculeaticarpa | |
| M. biuncifera Benth. | ||||
| M. galeottii Benth. | ||||
| M. grahamii A. Gray | grahamii | |||
| M. lacerata Rose | ||||
| M. purpusii Brandegee | ||||
| Distachyae | M. benthamii J.F. Macbr. | benthamii | ||
| M. luisana Brandegee | ||||
| M. polyantha Benth. | ||||
| Boreales | M. calcicola B.L. Rob. | |||
| M. depauperata Benth. | ||||
| M. monancistra Benth. | ||||
| M. pringlei S. Watson | pringlei | |||
| M. similis Britton & Rose | ||||
| M. texana (A. Gray) Small | filipes (Britton & Rose) Barneby | |||
| M. texana (A. Gray) Small | texana | |||
| Habbasia | Habbasia | M. pigra L. | pigra | |
| Mimosa | Mimosa | Mimosa | M. albida Humb. & Bonpl. ex Willd. | albida |
| M. albida Humb. & Bonpl. ex Willd. | strigosa (Willd.) B.L. Rob. | |||
| Lactifluae | M. caerulea Rose | |||
| M. goldmanii B.L. Rob. | ||||
| M. lactiflua Delile ex Benth. | ||||
| M. tricephala Schltdl. & Cham. | ||||
| Mimosa sp. |
Selección y tratamiento de los folíolos
Considerando la variabilidad de cada taxon, se seleccionaron tres individuos de cada uno. La copa de cada individuo se dividió en cuatro cuadrantes para la selección de pinnas de las hojas. De cada cuadrante, se recolectó el mayor número de pinnas; éstas se mezclaron para obtener una muestra compuesta por individuo. Las pinnas se depositaron en frascos, se fijaron con FAA (1:1:1) y se mantuvieron a temperatura ambiente en el laboratorio de Biosistemática de Leguminosas, UAM Iztapalapa, en la Ciudad de México. Los ejemplares de referencia están depositados en el Herbario Metropolitano (UAMIZ), de la Universidad Autónoma Metropolitana, Unidad Iztapalapa.
En el laboratorio, se eligieron los folíolos de la parte media de cada pinna y se lavaron con agua destilada. Posteriormente, se colocaron en cloro comercial durante 4 a 72 h a temperatura ambiente (Sandoval, 2005). Una vez aclarados, se deshidrataron en alcoholes graduales de 50 a 100%, y se elaboraron preparaciones permanentes con resina sintética (10 preparaciones/taxon). Por preparación, se observaron 20 folíolos, haciendo un total de 200 folíolos por taxon. De los 24 taxa, se observaron 4800 folíolos en total. Se utilizó un microscopio óptico (Zeiss, Primo-Star, Alemania). Las preparaciones permanentes están bajo resguardo del laboratorio de Biosistemática de Leguminosas.
Resultados
La sección Batocaulon fue la mejor representada; en ella, los taxa estudiados están ubicados principalmente en las series Acanthocarpae y Boreales (Cuadro 1).
Los folíolos anfistomáticos se encontraron en 22 de los taxa. En el caso de Mimosa benthamii J.F. Macbr. var. benthamii y M. caerulea Rose, las hojas son de tipo hipostomático. Asimismo, los 24 taxa estudiados presentan complejos estomáticos de tipo paracítico, caracterizados por presentar dos células acompañantes paralelas al eje longitudinal de las células oclusivas (Cuadro 2, Fig. 1).
Cuadro 2: Estado, coordenadas y altitud (msnm), forma de vida, tipo de vegetación, tipo de folíolo, complejo estomático y números de colecta de cada taxon de Mimosa L. estudiado, presente en México. Los ejemplares de referencia están depositados en el Herbario Metropolitano (UAMIZ), de la Universidad Autónoma Metropolitana, Unidad Iztapalapa.
| Taxa | Estado | Coordenadas /Altitud | Forma de vida | Tipo de vegetación | Tipo de folíolo | Complejo estomático | Números de colecta |
| M. aculeaticarpa Ortega var. aculeaticarpa | Querétaro | 20°48.406'N 100°30.686'O 2060 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 152, SAMA 153, SAMA 154 |
| M. albida Humb. & Bonpl. ex Willd. var. albida | Morelos | 18°57'50.1"N 99°16'39.2"O 1269 | Arbusto | Bosque de pino | Anfistomático | Paracítico | SAMA 227, SAMA 228, SAMA 229 |
| M. albida var. strigosa (Willd.) B.L. Rob. | Veracruz | 18°49.627'N 96°48.082'O 475 | Arbusto | Pastizal | Anfistomático | Paracítico | SAMA 142, SAMA 143, SAMA 144 |
| M. benthamii J.F. Macbr. var. benthamii | Oaxaca | 18°07.733'N 97°41.023'O 1720 | Árbol | Bosque tropical caducifolio | Hipostomático | Paracítico y Anomocítico | SAMA 131, SAMA 132, SAMA 133 |
| M. biuncifera Benth. | Querétaro | 20°48'24.2"N 100°30'36.5"O 2040 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 148, SAMA 157, SAMA 158 |
| M. caerulea Rose | Morelos | 18°58' 3.5" N 99°16'36.9"O 1955 | Arbusto | Bosque de pino | Hipostomático | Paracítico | SAMA 209, SAMA 210, SAMA 211 |
| M. calcicola B.L. Rob. | Puebla | 18°24'09.8"N 97°26'19.2"O 1720 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 108, SAMA 109, SAMA 110 |
| M. depauperata Benth. | Querétaro | 20°55.78'N 99°90.45'O 1820 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 160, SAMA 161, SAMA 162 |
| M. galeottii Benth. | Morelos | 18°59'16.5"N 99°14'07.9"O 1952 | Árbol | Bosque de pino | Anfistomático | Paracítico | SAMA 203, SAMA 204, SAMA 205 |
| M. goldmanii B.L. Rob. | Morelos | 18°33.867'N 99°06.250'O 897 | Árbol | Bosque tropical caducifolio | Anfistomático | Paracítico y Anisocítico | SAMA 213, SAMA 214, SAMA 215 |
| M. grahamii A. Gray var. grahamii | Querétaro | 20°48'27.6"N 100°30'48.7"O 2080 | Arbusto | Bosque tropical caducifolio | Anfistomático | Paracítico | SAMA 254, SAMA 255, SAMA 256 |
| M. lacerata Rose | Puebla | 18°41'31"N 97°24'01.3"O 2232 | Árbol | Matorral xerófilo | Anfistomático | Paracítico | SAMA 78, SAMA 79, SAMA 80 |
| M. lactiflua Delile ex Benth. | Oaxaca | 17°44.634'N 97°41.653'O 1830 | Arbusto | Bosque tropical caducifolio | Anfistomático | Paracítico | SAMA 128, SAMA 129, SAMA 130 |
| M. luisana Brandegee | Puebla | 18°18.732'N 97°30.023'O 1430 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico y Anisocítico | SAMA 88, SAMA 89, SAMA 90 |
| M. monancistra Benth. | Querétaro | 20°48'24.7"N 100°30'41.8"O 1070 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico y Anisocítico | SAMA 147, SAMA 156, SAMA 159 |
| M. pigra L. var. pigra | Veracruz | 17°41'13.2"N 94°56'16.8"O 250 | Arbusto | Pastizal | Anfistomático | Paracítico | SAMA 145, SAMA 146, SAMA 146b |
| M. polyantha Benth. | Puebla | 18°18.732'N 97°30.023'O 1430 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 98, SAMA 99, SAMA 100 |
| M. purpusii Brandegee | Puebla | 18°41'31"N 97°24'01.3"O 2232 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 83, SAMA 84, SAMA 85 |
| M. pringlei S. Watson var. pringlei | Querétaro | 20°53.518'N 99°39.679'O 2070 | Arbusto | Bosque de Pino | Anfistomático | Paracítico y Anisocítico | SAMA 166, SAMA 167, SAMA 168 |
| M. similis Britton & Rose | Querétaro | 20°92.30'N 99°75.70'O 1490 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico y Anisocítico | SAMA 170, SAMA 171, SAMA 172 |
| M. texana (A. Gray) Small var. filipesBritton & Rose) Barneby | Puebla | 18°12'0.46"N 97°31'28.6"O 2050 | Árbol | Bosque tropical caducifolio | Anfistomático | Paracítico | SAMA 103, SAMA 104, SAMA 105 |
| M. texana (A. Gray) Small var. texana | Querétaro | 20°42.388'N 99°40.766'O 1960 | Arbusto | Matorral xerófilo | Anfistomático | Paracítico | SAMA 163, SAMA 164, SAMA 165 |
| M. tricephala Schltdl. & Cham. | Morelos | 18°38.710'N 99°02.346'O 1070 | Arbusto | Bosque tropical caducifolio | Anfistomático | Paracítico, Anisocítico | SAMA 224, SAMA 225, SAMA 226 |
| Mimosa sp. | Morelos | 18°34.671'N 99°02.249'O 931 | Árbol | Bosque tropical caducifolio | Anfistomático | Paracítico | SAMA 217, SAMA 218, SAMA 219 |
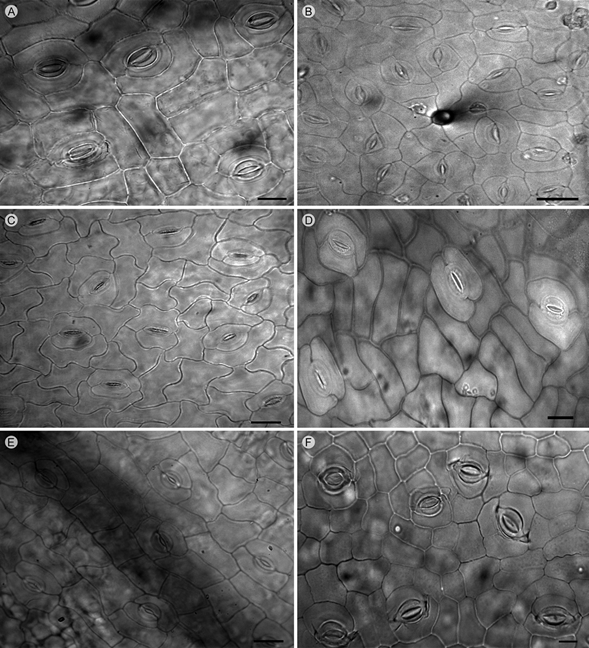
Figura 1: Complejos estomáticos paracíticos. A. Mimosa aculeaticarpa Ortega var. aculeaticarpa; B. M. albida Humb. & Bonpl. ex Willd. var. strigosa (Willd.) B.L. Rob.; C. M. caerulea Rose; D. M. lactiflua Delile ex Benth.; E. M. polyantha Benth.; F. M. texana (A. Gray) Small var. texana. Escala: 20 µm.
En el caso de M. goldmanii B.L. Rob., M. luisana Brandegee, M. monancistra Benth., M. pringlei var. pringlei S. Watson, M. similis Britton & Rose y M. tricephala Schltdl. & Cham.,además de presentar complejos estomáticos de tipo paracítico, tienen complejos anisocíticos, los que se caracterizan por tener tres células acompañantes de distintos tamaños que rodean a las células oclusivas. Cabe señalar que éstos se encuentran en menor proporción, pero en la misma superficie epidérmica (Fig. 2). Mimosa benthamii var. benthamii también tiene complejos estomáticos paracíticos y presenta otros anomocíticos.
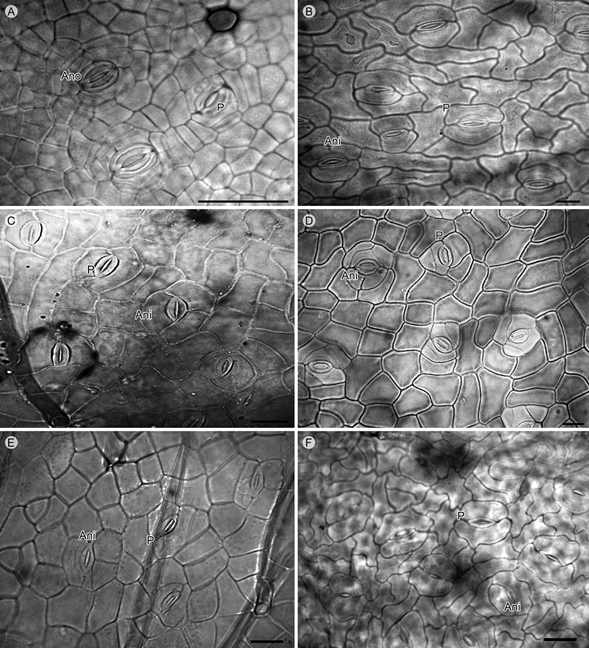
Figura 2: Complejos estomáticos anomocíticos (Ano), anisocíticos (Ani) y paracíticos (P). A. Mimosa benthamii J.F. Macbr. var. benthamii; B. M. goldmanii B.L. Rob.; C. M. monancistra Benth.; D. M. pringleiS. Watson var. pringlei; E. M. similis Britton & Rose; F. M. tricephala Schltdl. & Cham. Escala: 20 µm
De manera adicional, se observó que los folíolos de M. benthamii var. benthamii, M. calcicola B.L. Rob., M. luisana, M. purpusii Brandegee y M. tricephala presentan abundantes tricomas unicelulares. En el caso de M. purpusii y M. tricephala, la presencia de dichos tricomas dificultó la observación de sus complejos estomáticos.
Discusión
Los resultados obtenidos permiten separar a los taxa de Mimosa estudiados en dos grupos: i) los que presentan folíolos hipostomáticos, y ii) los que presentan folíolos anfistomáticos.
Mimosa benthamii var. benthamii y M. caerulea son los únicos taxa que tienen folíolos hipostomáticos. Asimismo, Bässler (1985) observó este tipo de folíolos en M. fagaracantha Griseb., M. pulverulenta Urb., M. ekmanii Urb. y M. apleura Urb. Chiamolera et al. (2011) también los observaron en M. scabrella Benth. Por otra parte, los 22 taxa restantes presentan folíolos de tipo anfistomático, al igual que M. distachya Cav., M. berlandieri A. Gray (=M. pigra L. var. berlandieri (A. Gray ex Torr.) B.L. Turner), M. invisa Mart. (=M. diplotricha C. Wright ex Sauvalle var. diplotricha), M. viva Vell. (Bässler, 1985), M. diplotricha var. diplotricha, M. himalayana Gamble y M. pudica L. (Begum y Borthakur, 2013). Por lo expuesto, son 29 taxa con esta característica, de las ca. 530 especies conocidas a nivel mundial (Särkinen et al., 2011; Simon et al., 2011). En forma adicional, Percy y Santa (1976) señalan el tipo de complejo estomático paracítico, presente en M. colombiana Britton & Killip y M. sensitiva L. Sin embargo, no hacen referencia al tipo de folíolo presente (hipostomático o anfistomático), al igual que Chiamolera et al. (2011), quienes no lo indican en M. scabrella. En la actualidad, se contabilizan ca. 38 taxa estudiados, lo que representan ca. 7% de los taxa de Mimosa a nivel mundial, siendo todavía prematuro el afirmar que el tipo de complejo estomático y su presencia en una o en ambas superficies del folíolo pudiese ser considerado un carácter con valor taxonómico a nivel genérico o infragenérico.
De la misma forma, los taxa de Mimosa estudiados presentan complejos estomáticos de tipo paracítico, los que fueron considerados por Metcalfe y Chalk (1979) como distintivos de las leguminosas. Esto fue confirmado por Percy y Santa (1976) al observar el mismo tipo de complejo estomático en 300 especies de esta familia.
Estudios posteriores reportan la presencia del complejo estomático paracítico en folíolos de taxa pertenecientes a dos géneros de mimosoideas: Acacia (Duarte y Wolf, 2005; Hernández y Arambarri, 2010) y Prosopis L. (Vilela, 1993); así como de los géneros Astragalus L. (Fahmy, 1997) e Indigofera L. (Martínez, 1997), pertenecientes a la subfamilia Papilionoideae.
En el caso de Mimosa, los complejos estomáticos paracíticos ya habían sido observados en M. albida Humb. & Bonpl. ex Willd., M. colombiana, M. pigra L., M. pudica, M. pulverulenta y M. sensitiva (Shah et al., 1972; Percy y Santa, 1976; Bässler, 1985; Rothe y Bathe, 2014). Sin embargo, no se especifica la variedad estudiada en estas especies, con excepción de M. colombiana y M. pulverulenta, ya que no se reconoce ninguna variedad (Barneby, 1991).
Por otra parte, Begum y Borthakur (2013) reportaron complejos estomáticos paracíticos en M. diplotricha var. diplotricha, M. himalayana y M. pudica; aunque señalan que M. himalayana, además del complejo estomático paracítico, presenta complejos anisocíticos. Stenglein et al. (2005) consideran que la presencia de más de un tipo de complejo estomático es común en leguminosas. Sin embargo, hasta ahora era distintivo de M. himalayana, pero en este estudio también se observó en M. benthamii var. benthamii, M. goldmanii, M. luisana, M. monancistra,M. pringlei var. pringlei y M. similis.
Por lo tanto, la presencia de dos tipos de complejos estomáticos en los folíolos de un mismo taxon permite separarlos en dos grupos: i) taxa con un sólo tipo de complejo estomático, y ii) taxa con dos tipos de complejo estomático, lo que permite distinguir por la combinación de tipos de complejos estomáticos presentes. Por ejemplo, M. benthamii var. benthamii es el único taxon con estomas paracíticos y anomocíticos; mientras que M. goldmanii, M. luisana, M. monancistra, M. pringlei y M. similis presentan complejos estomáticos paracíticos y anisocíticos; por lo que, a nivel específico, la presencia de dos tipos de complejos estomáticos en un mismo folíolo puede considerarse como un carácter útil para diferenciar entre los taxa. Sin embargo, la presencia de complejos estomáticos paracíticos, o de dos tipos de complejos en la misma superficie foliar, no es exclusivo de una serie. Asimismo, a nivel de las secciones del género, es prematuro considerarlo un carácter útil, ya que, aunque se tienen representadas todas las secciones de Mimosa, únicamente se han estudiado los estomas de un taxon de las secciones Calothamnos, Habbasia y Mimadenia.
Conclusiones
Los taxa estudiados se agruparon considerando tres caracteres con valor taxonómico: i) folíolos hipostomáticos vs. folíolos anfistomáticos, ii) presencia de un tipo de complejo estomático vs. dos tipos de complejos estomáticos, y iii) complejos estomáticos paracíticos y anisocíticos vs. complejos estomáticos paracíticos y anomocíticos.
Hasta este momento, se conoce el tipo de complejo estomático de ca. 7% de los taxa de Mimosa a nivel mundial y el 23% de los taxa presentes en México, por lo que todavía se requiere estudiar el 93 y 77% de los taxa de Mimosa presentes en el mundo y en México, respectivamente.
Contribuciones de autores
SAMA concibió y diseñó el estudio, trabajo de campo y desarrollo experimental. SAMA, SLCR y RG realizaron el análisis e interpretación de los datos. SAMA redactó el manuscrito con ayuda de SLCR y RG. Todos los autores contribuyeron con la discusión, revisión y aprobación del manuscrito final.

