











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
La familia Cactaceae del orden Caryophyllales es un grupo de plantas suculentas representativo de las zonas áridas y semiáridas del hemisferio Oeste, excepto Rhipsalis baccifera (Sol.) Stearn, la cual se encuentra en África y Asia (Hunt, 2006).
Su división en cuatro subfamilias (Pereskioideae, Opuntioideae, Maihuenioideae y Cactoideae) ha sido sustentada por caracteres morfológicos y moleculares (Hunt, 2006; Bárcenas et al., 2011). La subfamilia Cactoideae comprende el mayor número de especies y es la más diversa en términos de hábitos de crecimiento y formas de vida; se divide en nueve tribus: Browningeae, Cacteae, Calymmantheae, Cereeae, Hylocereeae, Notocacteae, Pachycereeae, Rhipsalidae, Trichocereeae (Ortega-Baes et al., 2010; dos Santos et al., 2012; Vázquez-Sánchez et al., 2012). En la tribu Cacteae, por ejemplo, predominan géneros como Aztekium Boed. y Mammillaria Haw., que incluyen representantes con tallos cortos, mientras que Echinocactus Link & Otto y Ferocactus Britton & Rose tienen integrantes con tallos globosos que miden 1 m o más de altura, así como Escobaría Britton & Rose, Neolloydia Britton & Rose y Thelocactus (K. Schum.) Britton & Rose, con especies que forman tallos de entre 20 y 30 cm de alto (Vázquez-Sánchez et al., 2012).
Por otro lado, para sobrevivir y prosperar en ambientes hostiles, la mayoría de las cactáceas cuentan con la capacidad de sintetizar compuestos químicos variados lo que las hace productos naturales promisorios en la búsqueda de nuevas fuentes de compuestos con actividad biológica. La presencia de fitoquímicos, como esteroles, alcaloides y compuestos fenólicos (Garza-Padrón et al., 2010) que muestran actividad biológica de tipo antioxidante benéfica para el ser humano, ha sido demostrada en diversas especies, tal es el caso de Astrophytum myriostigma Lem. (Garza-Padrón et al., 2010) y diferentes especies de Opuntia Mill. (Matías et al., 2014).
Para las especies de Mammillaria, aunque es un género que incluye un gran número de especies importantes en diversos aspectos, existen pocos estudios con relación a estudios anatómicos (Janu y Raghuvanshi, 2011; Mosco, 2012; Herrera-Martínez et al., 2015), y a su perfil fitoquímico. La información existente se enfoca al estudio de los pigmentos presentes en el tallo (Wybraniec y Nowak-Wydra, 2007; Mosco, 2012), el perfil fitoquímico y componentes nutracéuticos del fruto (Aparicio-Fernández et al., 2013), así como la inducción de metabolitos secundarios en cultivos de células en suspensión (Robles-Zepeda et al., 2009).
Por lo anterior, los objetivos del presente trabajo incluyeron la caracterización anatómica-histoquímica y el perfil fitoquímico del tallo y la raíz de Mammillaria uncinata Zuccarini & Pfeiffer, con la finalidad de contribuir al establecimiento de las bases para el aprovechamiento de esta especie como fuente de metabolitos secundarios con actividad biológica.
Materiales y métodos
Se llevó a cabo la recolecta de ejemplares de Mammillaria uncinata en sus áreas de distribución en municipios de Jalisco, México (Cuadro 1). Se recolectaron en promedio cinco individuos de cada localidad; dos de ellos se utilizaron para realizar el estudio morfo-anatómico. Para ello, se tomaron muestras de tejido de los tubérculos y parte interna del tallo, así como muestras de la raíz. Fueron procesados dos ejemplares por localidad para el estudio de perfil fitoquímico y un ejemplar completo de cada región de muestreo se depositó en el herbario IBUG de la Universidad de Guadalajara.
Cuadro 1: Localización geográfica de localidades de recolecta de Mammillaria uncinata Zuccarini & Pfeiffer en el estado de Jalisco, México.
| Localidad | Latitud (N) | Longitud (O) | Altitud (m s.n.m.) | Colector/No. colecta |
| Lagos de Moreno, Jal. | 21°31' | 101°41' | 1930 | S. Loza-Cornejo 226-230 (IBUG) |
| Ojuelos de Jalisco, Jal. | 21°33' | 103°09' | 2100 | S. Loza-Cornejo 231-236 (IBUG) |
| Unión de San Antonio, Jal. | 21°02' | 101°27' | 1920 | S. Loza-Cornejo 237-241 (IBUG) |
| Encarnación de Díaz, Jal. | 21°24' | 102°25' | 1851 | S. Loza-Cornejo 242-246 (IBUG) |
Anatomía e histoquímica
Las muestras recolectadas de tallo y raíz de los individuos de M. uncinata fueron fijados con solución FAA (formaldehido-alcohol-ácido acético glacial-agua destilada, 1: 5: 0.5: 3.5) por 24-72 horas y procesados para el estudio anatómico, siguiendo la microtécnica convencional de inclusión en parafina. Se realizaron cortes transversales y longitudinales de un grosor de 12-14 μm con un microtomo rotatorio (RMT-30 Radical Instruments, India), los cuales se pegaron en portaobjetos con adhesivo de Haupt, dejando secar por 12 horas. Se continuó con inmersiones en diferentes concentraciones de xileno y alcohol etílico (100% a 50%) para eliminar la parafina. La tinción de los tejidos se realizó con safranina 0.05% con 2% de cloruro de sodio, disueltos en agua y verde rápido disuelto en alcohol etílico absoluto, aceite de clavo y metilcelosolve (López-Curto et al., 1998). Posteriormente se montaron con resina sintética; una vez secas y limpias las preparaciones, se procedió a su observación con un analizador de imágenes Image Manager (Imagic Bildverarbeitung AG, 1992-2005, Berlin, Alemania), adaptado a un microscopio óptico (Leica DM-1000, Wetzlar y Mannheim, Alemania). Para el análisis con microscopía electrónica de barrido, fueron fijadas muestras del tallo en FAA, lavadas y deshidratadas en serie de alcoholes y preparadas para su observación en un microscopio electrónico (JSM-6390, Jeol, Japón) operando a 30 Kv. Para la caracterización histoquímica del tallo y de la raíz, se hicieron cortes en fresco a mano alzada, aplicando diferentes pruebas de tinción (lugol, sudán IV, Ácido Peryódico-Reactivo de Schiff (APS) y vainillina-HCl) para la identificación de almidón, lípidos, polisacáridos insolubles y taninos, respectivamente (López-Curto et al., 1998).
Perfil fitoquímico
El tallo y raíz de M. uncinata de las plantas limpias y sin espinas se cortaron en cubos de aproximadamente 1 cm de lado, los cuales posteriormente fueron deshidratados bajo condiciones ambientales (14-28 °C; 44-58% HR) durante siete días hasta la pérdida de 85 a 95% de humedad, la cual se determinó mediante el registro de la pérdida diaria de peso. Las muestras secas se trituraron manualmente en un mortero y el polvo obtenido se sometió a extracciones secuenciales usando reflujo durante 30 minutos para la obtención de cuatro extractos de polaridad creciente (hexano, acetato de etilo, 80% etanol, agua). Los extractos se almacenaron en refrigeración (4 °C) y se protegieron de la luz hasta su análisis. Las determinaciones se llevaron a cabo por triplicado empleando técnicas cualitativas que indican la presencia de los diferentes metabolitos mediante cambios de coloración y/o formación de precipitado, y de acuerdo a las metodologías propuestas por Domínguez (1973). La presencia de alcaloides se detectó utilizando reactivos de Dragendorff, Mayer, Warner y Hager. Los reactivos de Libermann-Burchard y Salkowski se emplearon para revelar la presencia de esteroles. La existencia de flavonoides se descubrió utilizando la prueba de Shinoda y la de las saponinas mediante la formación de espuma y con el reactivo de Rosenthaler. Se empleó FeCl3 para la detección presuntiva de taninos y soluciones acuosas de gelatina, sal y gelatina-sal, como prueba confirmatoria. La presencia de quinonas se observó utilizando NaOH y tolueno en solución acuosa, después de tratar los extractos con H2O2 y H2SO4. Todas las técnicas empleadas fueron descritas por Sánchez-Herrera et al. (2011).
Resultados
Anatomía e histoquímica
La cutícula del tallo es relativamente delgada, lisa, inicia desde el nivel apical y continúa hasta las regiones cercanas a la base. La epidermis está presente en todas las regiones verdes del tallo y es uniestratificada (Fig. 1A). Las células de la epidermis son de forma cuadrada a rectangular con los bordes curvos en vista longitudinal, y de forma cuadrada en corte transversal; en vista superficial tienen paredes onduladas (Fig. 1C, D). Estas células no presentan algún tipo de contenido. Los estomas son paracíticos. La hipodermis es multiestratificada, colenquimatosa, compuesta por dos a cuatro estratos celulares (Fig. 1A, B). La peridermis es unidireccional, inicia en forma de parches en algunas partes del tallo y tiene un mayor desarrollo en la región media y basal. El felógeno presenta dos estratos de células con paredes delgadas, de forma rectangular; el felema está compuesto por cuatro o cinco estratos de células con pared delgada, las cuales aparecen colapsadas y alternan con estratos de células, en donde las paredes externas y radiales son gruesas y lignificadas (Fig. 1E, F). El córtex está constituido por tejido fundamental con dos tipos de células, las del clorénquima, de forma rectangular en sentido radial y cloroplastos abundantes (Fig. 2A, B) y granos de almidón (Fig. 3A, B), y las células del parénquima de reserva, las cuales son isodiamétricas, tienen paredes delgadas y también almacenan almidón. Células de mucílago y haces vasculares corticales formados por xilema y floema primarios están presentes en el tallo. En la región basal, los haces vasculares corticales ocasionalmente se presentan en grupos de hasta cuatro haces, con elementos de vaso estrechos que presentan pared secundaria helicoidal y floema con elementos de tubo criboso y células acompañantes (Fig. 2C-G). Una red de laticíferos se extiende en el córtex reservante (Fig. 4A) y se tiñe fuertemente con lugol, lo que indica la presencia de almidón. Cristales prismáticos o romboidales individuales o en conglomerados dispersos en el córtex (Fig. 4B-F), se encuentran uno o dos por célula y son relativamente grandes. En las regiones del córtex los granos de almidón son abundantes (Fig. 3G). La presencia de polisacáridos insolubles, cuerpos lipídicos y taninos oxidados en las células del córtex reservante fue también una característica distintiva del tallo (Fig. 3C-F). Los cuerpos lipídicos se concentran en la periferia de la célula, adyacentes a las paredes celulares (Fig. 3I).
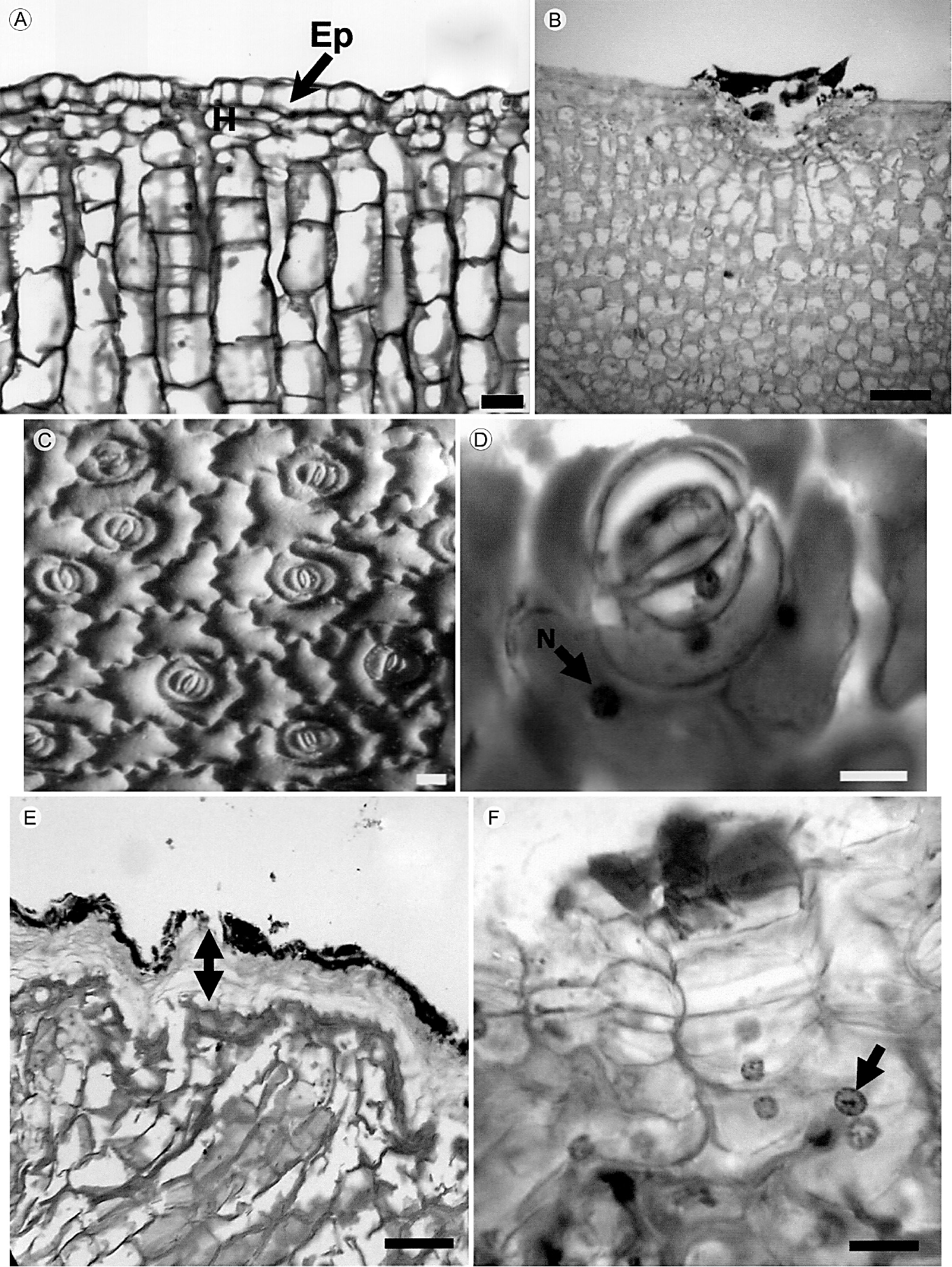
Figura 1 Cortes transversales y superficiales del tallo de Mammillaria uncinata Zuccarini & Pfeiffer; A. epidermis uniestratificada (flecha), hipodermis y clorénquima; B. inicio de formación de peridermis; C y D. vista superficial de células epidérmicas, núcleo (flecha) y estomas; E. corte transversal del tallo mostrando células de paredes delgadas (flecha doble punta) y células de pared gruesa lignificada más externas; F. detalle de células en división en el felógeno, con núcleo conspicuo (flecha). Ep, epidermis; H, hipodermis; N, núcleo. Escala: A-C, 30 μm; D-F, 100 μm.
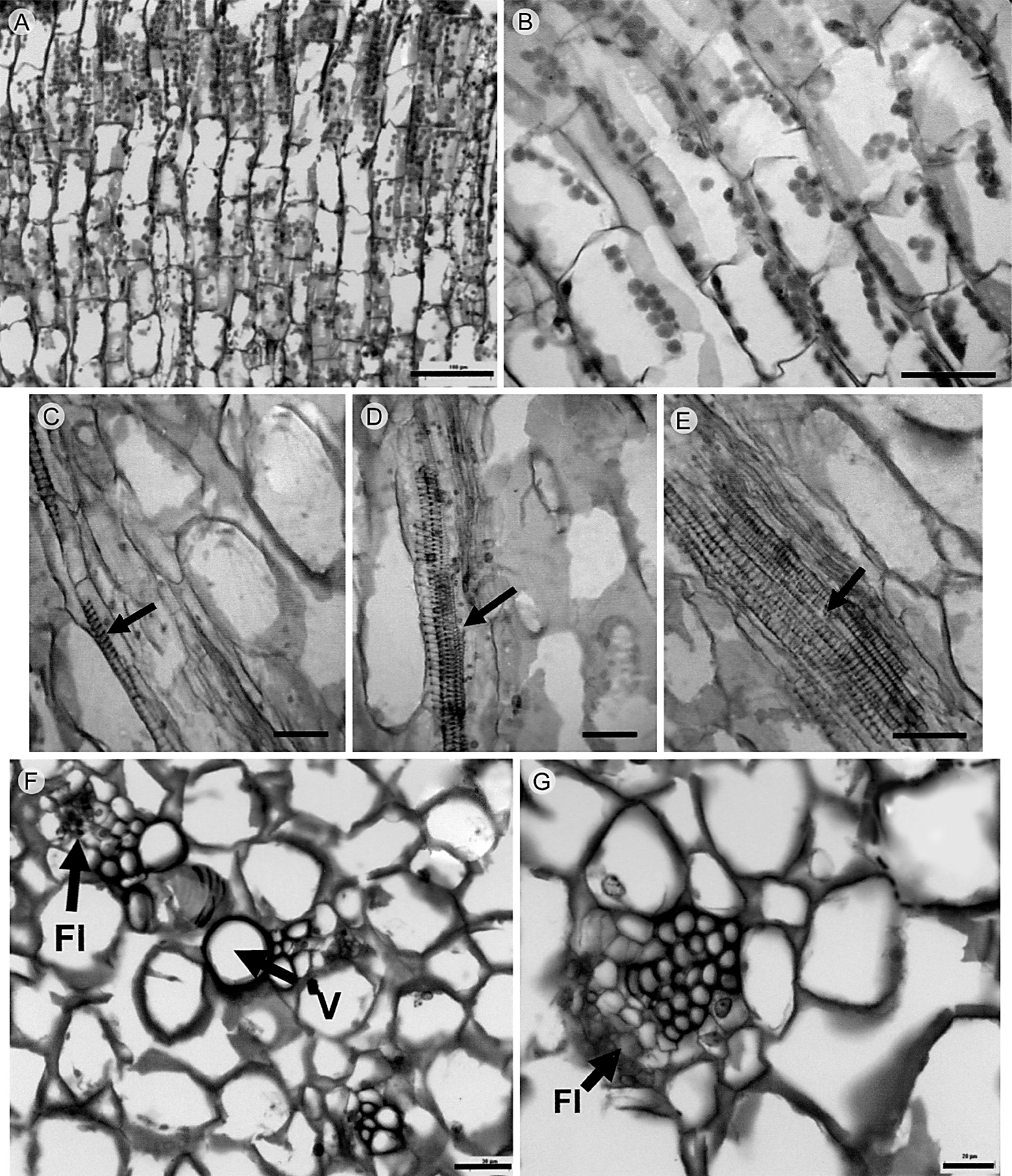
Figura 2 Cortes transversales y longitudinales del tallo de M. uncinata Zuccarini & Pfeiffer. A. y B. córtex fotosintético con abundantes cloroplastos; haces vasculares corticales de parte apical, media y basal del tallo; C, D y E. haces vasculares corticales de parte media y basal del tallo (flechas), note la mayor acumulación de xilema en la parte basal; F y G. haces vasculares con xilema y floema (flechas). Floema (Fl), vaso (V). Escala: A, F, G, 30 μm; B-E, 100 μm.
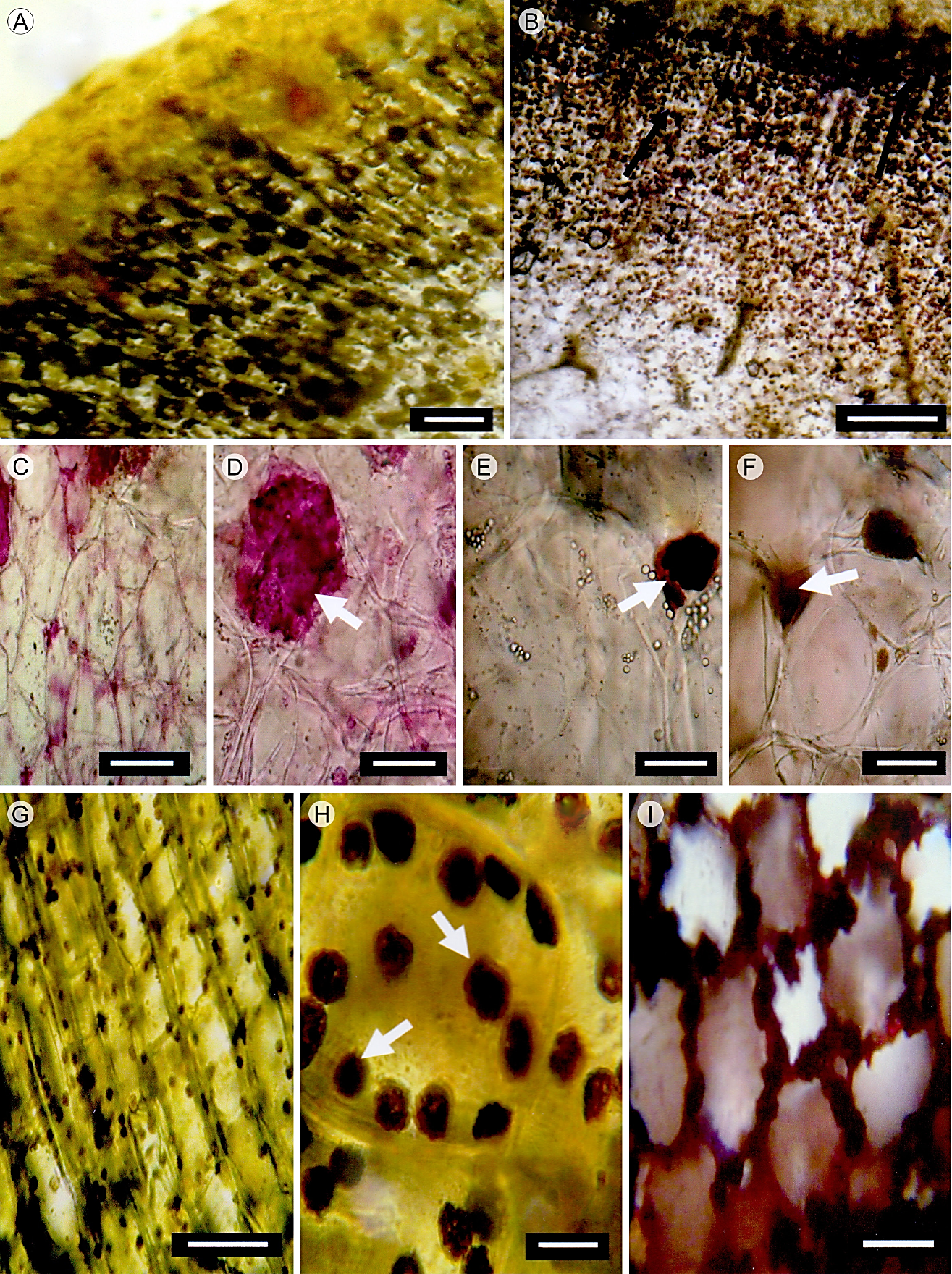
Figura 3 Cortes transversales del tallo de M. uncinata Zuccarini & Pfeiffer. A y B. abundantes granos de almidón en el córtex fotosintético y de reserva; C y D. polisacáridos estructurales en parénquima cortical de reserva (flecha); E y F. taninos oxidados (flecha); G. detalle de granos de almidón presentes en el clorénquima; H. granos de almidón (flecha) en el parénquima de reserva; I. cuerpos lipídicos en el parénquima de reserva. Escala: A y B 300 μm; C-F, 20 μm; G, 100 μm; H, I, 30 μm.
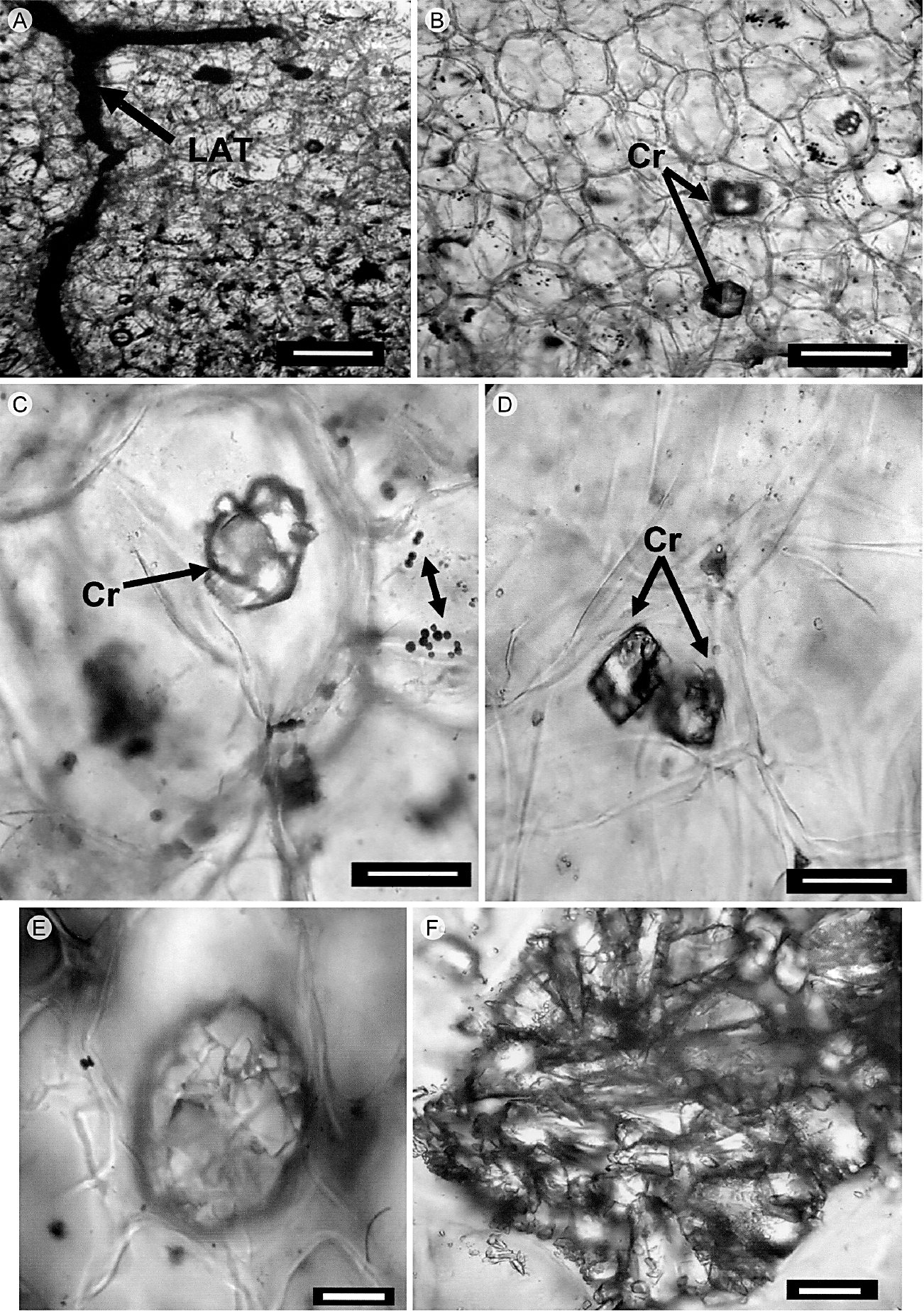
Figura 4 Cortes transversales del tallo de Mammillaria uncinata Zuccarini & Pfeiffer. A. laticífero (flecha); B-D. granos de almidón (flecha doble punta), cristales prismáticos (flechas) en parénquima de reserva; E y F. cristales agregados. LAT, laticífero; Cr, cristales. Escala: A y B, 100 μm; C-E, 30 μm; F, 28 μm.
El tejido vascular del xilema está constituido por una matriz de traqueidas de banda ancha, vasos escasos y células de parénquima de forma irregular (Figs. 5, 6). Los radios son no lignificados y son principalmente secundarios. El floema se compone por elementos de tubo criboso, células acompañantes y parénquima. Granos de almidón y cristales están presentes de manera abundante en el parénquima radial (Figs. 5C, 6C-D). Los haces vasculares se presentan en número variable (Fig. 5D) y tienden a formar un cilindro sólido de xilema en la parte basal del tallo. La médula es parenquimatosa y presenta haces vasculares dispersos formados por xilema y floema primarios (Fig. 6F, G).
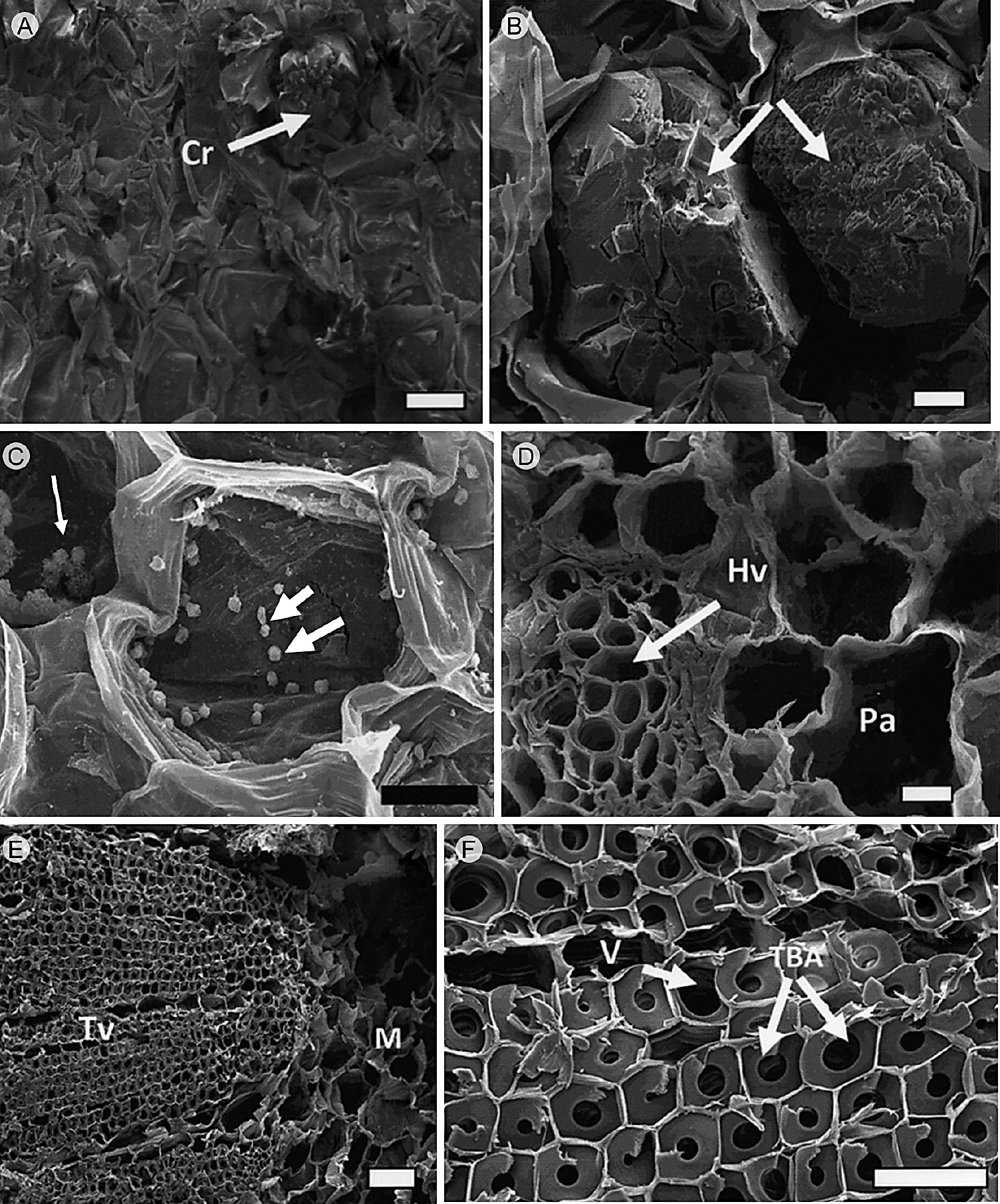
Figura 5 Cortes transversales del tallo de M. uncinata Zuccarini & Pfeiffer observados con microscopía electrónica de barrido. A, B. cristales agregados (flechas) en parénquima cortical del tallo; C. granos de almidón en parénquima cortical (flechas); D. detalle de haz vascular cortical (flecha) y parénquima; E. tejido vascular del tallo, y médula; F. detalle de tejido vascular del tallo mostrando una matriz de traqueidas de banda ancha (flechas), y vasos escasos. Cr, cristales; Hv, haz vascular; M, médula; Pa, parénquima; TBA, traqueidas de banda ancha; Tv, tejido vascular; V, vasos. Escala: A y E, 100 μm; B, 20 μm; C y F, 50 μm; D, 10 μm.
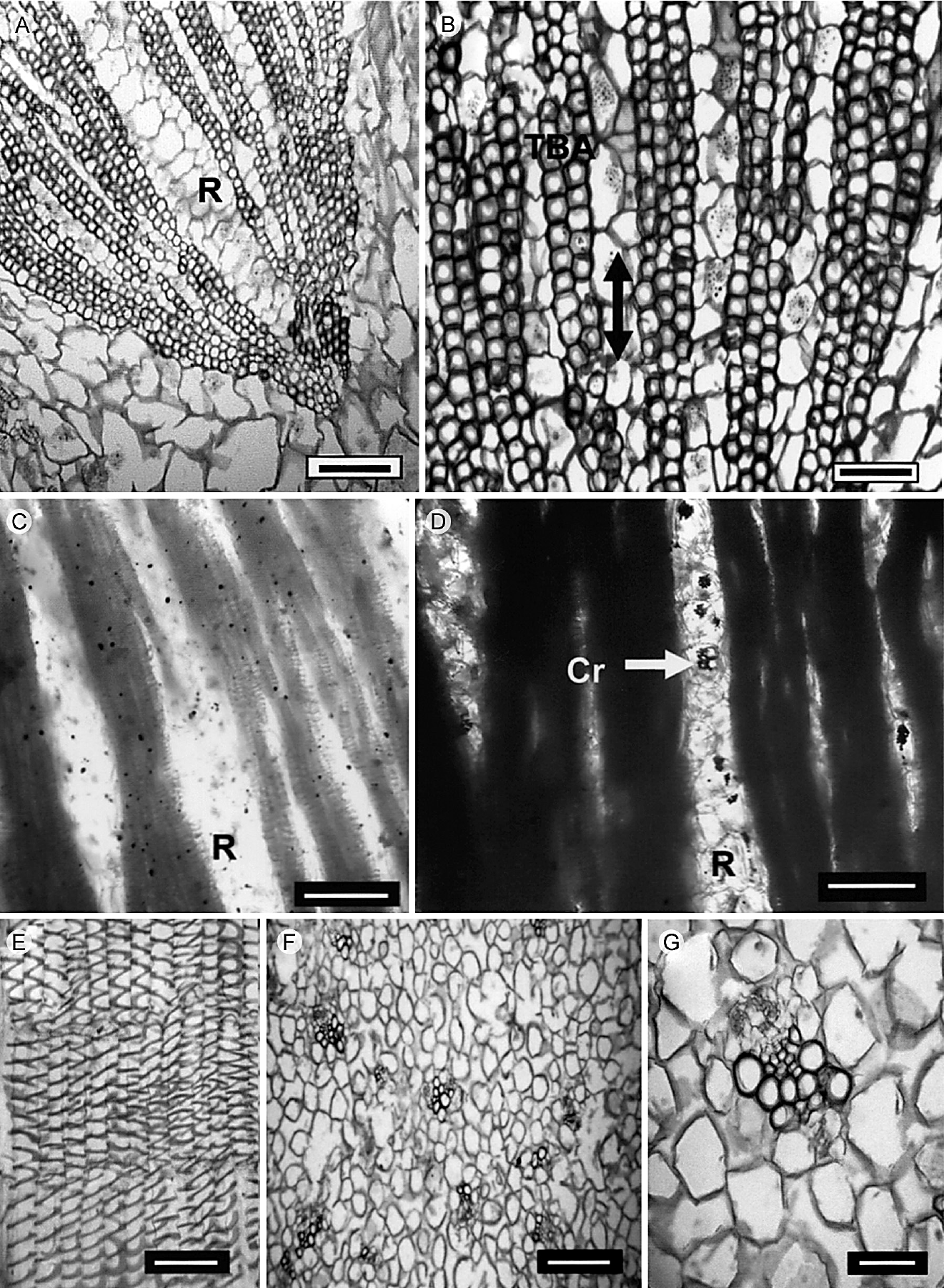
Figura 6 Cortes transversales y longitudinales del tallo de M. uncinata Zuccarini & Pfeiffer. A, B. radios no lignificados (flecha doble punta) y traqueidas de banda ancha en el xilema; C y D. células de los radios con granos de almidón y cristales (flecha); E. traqueidas de banda ancha estratificadas; F. haces vasculares en la médula; G. detalle de haz vascular medular. Cr, cristales; R, radios; TBA, traqueidas de banda ancha. Escala: A y B, 100 μm; C y D. 50 μm; E-G, 150 μm.
Se observaron raíces con crecimiento secundario. En corte transversal se observa una peridermis, parénquima, tejido vascular y una región central. La peridermis está formada por varios estratos de células colapsadas de paredes delgadas (Fig. 7A, B). En las raíces con mayor diámetro, estas células alternan con estratos de células de pared gruesa y lignificada. Las células de parénquima tienen forma isodiamétrica y se colapsan radialmente cerca de la peridermis conforme se avanza a la base de la raíz. El tejido vascular forma un anillo completo formado por el xilema secundario constituido por vasos, traqueidas de banda ancha y radios xilemáticos uniseriados, con células cuadradas a rectangulares (Fig. 7C-E).

Figura 7 Cortes transversales de la raíz de Mammillaria uncinata Zuccarini & Pfeiffer. A. desarrollo de la peridermis (flecha doble punta); B. radios uniseriados en el xilema (flecha); C-E. detalle de radios uniseriados y traqueidas de banda ancha (flechas) en el xilema. R, radios. Escala: A-C, 50 μm; D y E, 30 μm.
Perfil fitoquímico
El Cuadro 2 muestra los resultados del análisis de perfil fitoquímico realizado en cuatro extractos de diferente polaridad obtenidos a partir del tallo y la raíz de M. uncinata. Como se puede observar los metabolitos secundarios son más abundantes en el tallo, donde se detecta la presencia de cuatro grupos, mientras que en la raíz únicamente se detectó la presencia de dos de ellos. Los alcaloides son abundantes en los extractos de mayor polaridad (etanólico y acuoso) del tallo de M. uncinata, mientras que no se detectó su presencia en los extractos obtenidos de la raíz. Los esteroles se presentan en cantidades bajas a moderadas en extractos de ambas partes de la planta. La presencia en cantidades bajas y moderadas de este tipo de metabolitos en los extractos obtenidos con acetato de etilo y hexano, tanto del tallo como de la raíz, sugiere una menor polaridad de dichos compuestos. Los flavonoides fueron abundantes en los extractos de la parte aérea de M. uncinata, mientras que en la raíz no se detectaron. Fue abundante la presencia de saponinas en el extracto acuoso del tallo de M. uncinata, y escasa en el extracto etanólico del tallo y en el extracto acuoso de la raíz. La presencia de taninos y quinonas no pudo detectarse con las técnicas empleadas, sugiriendo que podrían encontrarse en cantidades muy pequeñas o en forma insoluble, formando parte de estructuras celulares.
Cuadro 2: Perfil fitoquímico de tallo y raíz de Mammillaria uncinata Zuccarini & Pfeiffer analizado en cuatro extractos de diferente polaridad. Extractos: H: hexano, AE: acetato de etilo, E: 80% etanol, A: agua (+)=Ensayo positivo. Contenido: + bajo, ++ moderado, +++ abundante. (-)=Ensayo negativo, ausencia.
| Tallo | Raíz | |||||||
| Extractos | ||||||||
| H | AE | E | A | H | AE | E | A | |
| Alcaloides | - | - | +++ | +++ | - | - | - | - |
| Esteroles | - | ++ | + | - | ++ | + | + | - |
| Flavonoides | ++ | ++ | +++ | +++ | - | - | - | - |
| Saponinas | - | - | + | +++ | - | - | - | + |
Discusión
Anatomía e histoquímica
Las características anatómicas observadas en Mammillaria uncinata son similares a las descritas para otras especies de Cactoideae (Terrazas y Arias, 2003; Salinas-Revilla y Álvarez-Moscoso, 2006; dos Santos García et al., 2012). El tallo de Mammillaria uncinata presenta una cutícula lisa y delgada que inicia a nivel apical y se conserva hasta las regiones cercanas a la base del tallo, similar a otros géneros de Cactoideae. La cutícula tiene una función importante al constituir una barrera primaria entre el cuerpo de la planta y las condiciones ambientales (Salinas-Revilla y Álvarez-Moscoso, 2006). La epidermis uniestratificada de células con pared delgada y lisa coincide con observaciones realizadas para especies de otras cactáceas (Muruaga y Guantay, 2011; dos Santos García et al., 2012; Gonçalves da Silva et al., 2013). En otras especies de Mammillaria (M. polythele Mart. y M. perbella Hildm. ex K. Schum.) la superficie de las células epidérmicas es lisa, excepto en algunas zonas donde es ligeramente rugosa (Gasson, 1981). En corte transversal, las células de la epidermis de M. uncinata muestran formas cuadradas o rectangulares, similar a otras cactáceas, por ejemplo, Corryocactus erectus (Backeb.) F. Ritter (Salinas-Revilla y Álvarez-Moscoso, 2006). La peridermis en el tallo se desarrolla principalmente en la región media y basal, es unidireccional y de origen epidérmico como se presenta comúnmente en Cactaceae (Terrazas y Arias, 2003; dos Santos-García et al., 2012; Gonçalves da Silva et al., 2013). Mammillaria uncinata presenta hipodermis formada por varios estratos de células de colénquima con paredes primarias. De acuerdo con Mauseth (2006), la existencia de este tipo de células en los tallos proporciona flexibilidad, permitiendo la contracción o expansión de éste, acorde con la disponibilidad de agua. Representaría, además, un mecanismo eficiente para proteger al tallo de depredadores (Soares da Silva et al., 2010; dos Santos-García et al., 2012).
El tejido cortical constituye una gran proporción del tejido suculento del tallo de M. uncinata. Su organización es típica de cactáceas ya que se diferencia en dos porciones: 1) las células exteriores del córtex que son ricas en cloroplastos y ocurren en columnas y 2) las partes centrales de tubérculos y tallo que consisten de células de parénquima isodiamétricas en un arreglo irregular. La disposición radial del clorénquima permite la difusión de CO2 a través del aumento en espacios intercelulares. Fue notoria la acumulación de granos de almidón en el clorénquima. La presencia de almidón en el córtex es de gran importancia, ya que éste es usualmente el principal carbohidrato insoluble en la mayor parte del tejido fundamental y constituye la forma más significativa de reserva de carbono en plantas en términos de su cantidad, distribución e importancia para el funcionamiento del metabolismo fotosintético (Kötting et al., 2009). En el córtex del tallo de M. uncinata es notoria la presencia de granos de almidón tanto en el clorénquima como en el parénquima reservante. Adicionalmente, en el parénquima de los radios del xilema fueron también abundantes los granos de almidón. Lindeboom et al. (2004) mencionan que las condiciones ambientales temperatura, sequía y disponibilidad de nutrientes influyen en la cantidad de almidón presente. Éste podría ser el caso de M. uncinata, ya que la especie se distribuye en localidades con escasa precipitación pluvial, por lo que mantener una reserva suficiente de carbohidratos es vital para la supervivencia. Los cuerpos lipídicos o cuerpos de aceite están presentes en el parénquima cortical. Los lípidos son constituyentes esenciales de todas las células vegetales. Su contenido varía entre 5-10% del peso seco. Semillas, granos de polen, flores, raíces y tallos almacenan lípidos como material de reserva en partículas subcelulares denominadas cuerpos lipídicos, cuerpos de aceite, oleosomas o esferosomas (Ho et al., 2014). Dichas reservas constituidas principalmente por triacilglicéridos (TAGs) y proteínas oleosinas pueden ser movilizadas durante periodos de metabolismo activo (Chen et al., 1999; van der Schoot et al., 2011; Chapman et al., 2012). La presencia de lípidos o compuestos lipofílicos, como estructuras subcelulares en el tallo de M. uncinata, además de formar parte de las reservas de carbono en el tallo, podría representar una barrera hidrofóbica para prevenir la pérdida de agua o formar parte de una cubierta protectora contra patógenos, ataques de herbívoros y otros factores ambientales, como mencionan Rodrigues et al. (2011).
Cristales y células de mucílago están presentes en el córtex reservante del tallo de M. uncinata. Los cristales presentan una morfología similar a los de otras especies de Cactaceae (Contreras-Padilla et al., 2011). De acuerdo a Frausto-Reyes et al. (2014), la composición de dichos cristales puede ser de oxalato de calcio mono o dihidratado. La presencia de células de mucílago y cristales en el córtex son ejemplos de adaptación a hábitats xéricos de M. uncinata. Los cristales pueden reflejar la excesiva irradiación solar, mientras que la acumulación de mucílago hidrofílico facilitaría el almacenamiento de agua. La formación de cristales, además, está generalmente asociada con mecanismos de regulación de calcio en tejidos y órganos cuando los niveles de éste se incrementan en el suelo (Hudgins et al., 2003), con la defensa contra herbívoros (Ruiz et al., 2002), así como con la regulación osmótica y detoxificación de metales (Nakata, 2003). La presencia de laticíferos es una característica distintiva del tallo de Mammillaria uncinata, similar a otras especies del mismo género (M. heyderi Muehlenpf., M. guerreronis (Bravo) Boed.) (Witler y Mauseth, 1984a, b). De acuerdo a estos autores, los laticíferos articulados presentes en Mammillaria constituyen un sistema secretor complejo que se forma por lisis de masas celulares cilíndricas que luego son rodeadas por un epitelio multicelular. El látex contenido en estos conductos contiene varias sustancias en solución y suspensión coloidal, entre las que se incluyen carbohidratos, ácidos orgánicos, sales, esteroles, grasas y mucílagos. Son comunes también los terpenoides (cis-1, 4 poliisoprenos) y gomas, aunque otras sustancias pueden estar presentes, tales como glicósidos cardíacos, alcaloides, azúcares, proteínas y taninos (Konno, 2011), algunos de los cuales se detectaron en el perfil fitoquímico. Los laticíferos contienen una gama amplia de metabolitos secundarios; sin embargo, ninguno es movilizado o participa en el metabolismo celular. De acuerdo a Konno (2011) y Roshchina y Roshchina (2012), los laticíferos sirven como sistemas para "secuestrar" metabolitos secundarios tóxicos, los cuales pueden tener una función de protección contra herbívoros; este podría ser el caso de Mammillaria uncinata. Los taninos son los metabolitos secundarios más abundantes producidos por las plantas y contribuyen por su toxicidad a la defensa contra insectos herbívoros al inducir en éstos un estrés oxidativo (Barbehenn y Constabe, 2011; Salminen y Karonen, 2011). La presencia de taninos en el tallo de M. uncinata fue escasa, solamente algunas células del córtex reservante presentaron este contenido, motivo por el cual no fueron detectados en el análisis del perfil fitoquímico.
El xilema secundario está constituido por una matriz de traqueidas de banda ancha, vasos escasos y células de parénquima. Los haces vasculares se presentan en número variable y tienden a formar un cilindro sólido de xilema secundario con radios no lignificados. El floema se compone por elementos de tubo criboso, células acompañantes y parénquima como se presenta también en otras cactáceas (Terrazas y Arias, 2003).
La raíz de Mammillaria uncinata está constituida por peridermis, parénquima, xilema y floema secundarios, además de las células de parénquima en la región central. Las células de parénquima subyacentes a la peridermis con disposición radial que ocupan una mayor proporción de tejido en la raíz. La peridermis se caracteriza por presentar varios estratos de células esclerificadas, semejante a otras especies de Cactaceae (Terrazas y Arias, 2003).
Perfil fitoquímico
Los resultados del estudio fitoquímico realizados en M. uncinata complementan el estudio histoquímico, mostrando un panorama más amplio sobre la composición química de la especie. Los diferentes tipos de metabolitos secundarios detectados en el análisis del perfil fitoquímico de M. uncinata son sintetizados en la planta para ayudar a su protección y supervivencia (Konno, 2011; Roshchina y Roshchina, 2012); sin embargo, todos ellos pueden presentar algún tipo de actividad biológica en el organismo humano. Por lo tanto, M. uncinata podría considerarse como una especie potencial como fuente de fitoquímicos.
La abundante presencia de alcaloides en los extractos polares obtenidos del tallo sugiere que estos compuestos se encuentran en forma de sales (Arango, 2002). La presencia de alcaloides ha sido reportada en el tallo de otras especies de cactáceas globosas; por ejemplo, Sánchez-Herrera et al. (2011) registraron mayor abundancia de alcaloides en el tallo de Coryphantha clavata (Scheidw.) Backeb., que en su raíz. Un comportamiento similar encontramos en la presente investigación, ya que solo se detecta la presencia de alcaloides en el tallo, mientras que en la raíz están ausentes. A la fecha no existen evidencias sobre las propiedades de los alcaloides presentes en M. uncinata, probablemente por el hecho de ser una especie no utilizada en la medicina tradicional, por lo que resulta de interés el continuar con el estudio en cuanto a la concentración y el tipo de alcaloides, ya que éstos podrían presentar algún tipo de actividad biológica.
Los esteroles, también llamados triterpenos, forman parte de estructuras importantes en las membranas de células vegetales, y cumplen diversas funciones que van desde protección (ceras) hasta regulación del crecimiento como son las fitoalexinas (Céspedes et al., 2005). Diferentes especies vegetales sintetizan esteroles, aunque en una variedad de formas y concentraciones, dependiendo de cada planta y de las condiciones ambientales (Piironen et al., 2000). De acuerdo a Domínguez (1973), los esteroles pueden encontrarse en diferentes formas químicas (libres, como ésteres, o glicósidos). En el presente trabajo se detectó la presencia de esteroles, presumiblemente en dos formas químicas diferentes, ya que los detectados en el tallo presentaron mayor polaridad que los de la raíz. La presencia de esteroles en especies de cactáceas ha sido estudiada y se ha mencionado que algunos de ellos presentan efectos biológicos. Por ejemplo, Jiang et al. (2006), concluyeron sobre el descubrimiento de dos nuevos esteroles en tallos de Opuntia dillenii (Ker Gawl.) Haw. Por otro lado, Sri Nurestri et al. (2008) mostraron resultados sobre la actividad citotóxica de extractos obtenidos de hojas de Pereskia bleo (Kunth) DC., los cuales son ricos en esteroles. La cactácea columnar Myrtillocactus geometrizans (Mart. ex Pfeiff.) Console presenta compuestos de tipo esteroles tanto en la parte aérea, como en la raíz y éstos tienen un efecto insecticida (Céspedes et al., 2005). El estudio de dichas moléculas fue retomado por Salazar et al. (2011) quienes demostraron que dichas sustancias tienen, además, actividad antiinflamatoria y citotóxica.
Los flavonoides son un grupo de compuestos fenólicos de amplia distribución en plantas, incluyendo diferentes especies de Cactaceae. Su comprobada capacidad como antioxidantes y otras formas de actividad biológica los ha hecho blanco de diferentes estudios (Alimia et al., 2010; Angulo-Bejarano et al., 2014; Gutiérrez-Grijalva et al., 2016). Los extractos obtenidos del tallo de M. uncinata muestran contenidos importantes de estos compuestos, mientras que en la raíz no se evidenció su existencia. Los resultados sugieren la presencia de diferentes tipos de flavonoides con distintas características de solubilidad, así como la importancia de la luz para su síntesis en las plantas, ya que únicamente se detectaron en el tallo. La presencia de flavonoides en los frutos de M. uncinata (Aparicio-Fernández et al., 2013), así como en especies de Opuntia (Matías et al., 2014; Hahm et al., 2015) ha sido establecida con anterioridad incluyendo su capacidad antioxidante.
Las saponinas son glucósidos solubles en agua con capacidad para disminuir la tensión superficial y formar espuma al agitar sus soluciones. Al hidrolizar las saponinas se forman carbohidratos y una aglicona llamada sapogenina que puede presentar estructuras de tipo esteroidal, triterpénico o tetracíclico, entre otras (Domínguez, 1973). La presencia de saponinas se detecta en los extractos más polares del tallo y raíz de M. uncinata sugiriendo que se encuentran en forma glicosilada. En investigaciones previas del equipo de trabajo (Aparicio-Fernández et al., 2013) se encontró la presencia de saponinas en los frutos de la misma cactácea. La presencia de estos compuestos ha sido descrita en otras especies de cactáceas (Okazaki et al., 2011; Kakuta et al., 2012), y algunas de ellas han demostrado actividad biológica, destacando la actividad antialérgica de tipo I.
La presencia de taninos no se detecta mediante las técnicas cualitativas empleadas en el análisis del perfil fitoquímico. Sin embargo, sí fueron detectados mediante pruebas histoquímicas, lo que sugiere que estos componentes se encuentran en pequeñas cantidades firmemente unidos a las paredes celulares y de forma insoluble. Por su importante efecto biológico como antioxidantes fuertes, los taninos podrían considerarse parte de la fibra antioxidante y a M. uncinata una fuente de ésta (Barbehenn y Constabe, 2011; Salminen y Karonen, 2011).
Conclusiones
Este estudio muestra la presencia de caracteres anatómicos en Mammillaria uncinata similares a otras especies de Cactaceae, destacando la presencia de cristales de morfología variable en el tallo, así como laticíferos. Dada la diversidad de compuestos fitoquímicos (alcaloides, esteroles, flavonoides y saponinas) detectados en el tallo y raíz de M. uncinata, se considera de trascendencia su descubrimiento, así como de relevancia el continuar con su estudio en la búsqueda de moléculas con actividad biológica que puedan ser aplicados en beneficio de la salud humana. Lo anterior sugiere la potencialidad de M. uncinata como fuente de fitoquímicos con actividad biológica.

