










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkActa botánica mexicana
versão On-line ISSN 2448-7589versão impressa ISSN 0187-7151
Act. Bot. Mex no.110 Pátzcuaro Jan. 2015
Composición y diversidad del matorral desértico rosetófilo en dos tipos de suelo en el noreste de México
Composition and diversity of the rosetophilous desert scrub in two soil types at northeastern Mexico
Eduardo Alanís-Rodríguez1,3, Arturo Mora-Olivo2, Javier Jiménez-Pérez1, Marco Aurelio González-Tagle1, José Israel Yerena Yamallel1, José Guadalupe Martínez-Ávalos2, Laura Elena González-Rodríguez2
1 Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales, carretera Linares-Cd. Victoria km 145, Apdo. postal 41, 67700 Linares, Nuevo León, México.
2 Universidad Autónoma de Tamaulipas, Instituto de Ecología Aplicada, División del Golfo 356, 87019 Ciudad Victoria, Tamaulipas, México.
3 Autor para la correspondencia: eduardo.alanisrd@uanl.edu.mx
Recibido en noviembre de 2012.
Aceptado en enero de 2014.
RESUMEN
Se estudiaron aspectos de la flora y la ecología del matorral desértico rosetófilo en dos áreas que difieren en el tipo de suelo en el centro de Nuevo León, México (Rendzina + Litosol, textura media, lítico y Regosol calcárico + Litosol, textura media). Utilizando 44 parcelas de 20 × 10 m se muestrearon las especies suculentas, arbustivas y arbóreas. Se calcularon los atributos de la vegetación tales como densidad (N/ha), cobertura de copa (m2/ha), frecuencia, índice de valor de importancia (IVI), además de la diversidad α y β. Mediante un ANOVA no se encontraron diferencias significativas en la riqueza, la diversidad y la cobertura foliar. Los resultados indican que la composición florística presentó disimilitud entre los dos tipos de suelos, siendo además la densidad significativamente mayor en el Regosol (8909 N/ha) que en la Rendzina (4790 N/ha). La predominancia de cactáceas es favorecida por la baja retención de humedad de la asociación Regosol calcárico + Litosol.
Palabras clave: composición florística, matorral xerófilo, Nuevo León, tipos de suelos.
ABSTRACT
The floristic and ecological characteristics of a rosette desert scrub in two areas differing in soil type (Rendzina + Lithosol, medium texture, lithic, and Regosol calcareous + Lithosol, medium texture) in northeastern Mexico were studied. Using 44 plots of 20 × 10 m we sampled succulents, shrubs and tree species. We estimated the following vegetation variables: density (N/ha), crown cover (m2/ha), frequency, importance value index (IVI), in addition to α- and β-diversity. By ANOVA, no significant differences were found in richness, diversity and crown cover. The results indicate that the floristic composition was different between the two soil types, the density being significantly higher in the Regosol (8909 N/ha) than in the Rendzina (4790 N/ha). The predominance of cacti is favored by low moisture retention in the calcaric + Litosol Regosol association.
Key words: floristic composition, Nuevo León, soils types, xerophilous scrub.
INTRODUCCIÓN
El matorral xerófilo constituye el tipo de vegetación mejor representado en el norte de México (Rzedowski, 1978). De hecho, en algunas entidades del norte ocupa las mayores coberturas vegetales, como es el caso de Nuevo León donde esta comunidad se presenta en 54.6% de toda su superficie (Palacio et al., 2000). En el noreste de México y otras regiones del país, esta formación vegetal está representada por comunidades como matorral submontano, matorral espinoso tamaulipeco, matorral desértico micrófilo y matorral desértico rosetófilo (González-Medrano, 2004).
Aunque algunos matorrales han sido mejor estudiados en el noreste de México (Briones y Villarreal, 2001; Alanís et al., 2008; Ugalde et al., 2008; Canizales et al., 2009; Jiménez-Pérez et al., 2009; Jiménez-Pérez et al., 2013), el matorral desértico rosetófilo ha recibido poca atención, no solo en esta región sino también a nivel nacional. Entre los pocos trabajos que han evaluado esta comunidad en México se encuentran el de Treviño-Carreón y Hernández-Sandoval (2000), el de Martorell y Ezcurra (2002) y el de Encina-Domínguez et al. (2013). En algunos contribuciones se destaca la relación de la vegetación con factores ambientales (González-Medrano, 1996, 2004; Huerta-Martínez y García-Moya, 2004; Rankin et al., 2007).
Algunos factores bióticos y abióticos influyen en la diversidad de plantas, como la fauna microbiana (Zak et al., 2003), el clima (González-Medrano, 1996, 2004) y muy especialmente del suelo (Abd, 2000; Gallego, 2003; Rankin et al., 2007). De hecho, la relación suelo-vegetación ha sido documentada tomando en cuenta que las plantas juegan un papel primordial en el proceso de la edafogénesis, pues el substrato deriva tanto de la fragmentación o meteorización de la roca como de la influencia de las raíces vegetales y su aporte de materia orgánica (Porta et al., 1999; Treviño-Carreón, 2004).
Existen pocos estudios para el noreste de México que relacionen los matorrales con variables edafológicas (Estrada-Castillón et al., 2012). Tomando en cuenta que los regosoles y las rendzinas son suelos comunes en las zonas áridas de México (Anónimo, 2005; Bautista et al., 2009), se consideró interesante comparar el matorral desértico rosetófilo presente en estos dos tipos de suelos, los cuales poseen características distintas en cuanto a su composición y capacidad de retención de humedad (Porta et al., 1999). Así, se planteó como objetivo conocer: (1) las variables de densidad (N/ha), cobertura de copa (m2/ha), frecuencia, índice de valor de importancia (IVI), y (2) las diversidades α y β de ambas comunidades vegetales.
MÉTODOS
Áreas de estudio
El estudio se realizó en dos áreas situadas en el noreste de México, pertenecientes al municipio de Mina, Nuevo León (Fig. 1). El clima es seco semicálido (BWhw), con una temperatura media anual que varía entre 21 y 22°C. Los suelos predominantes son rendzinas, litosoles y regosoles. La vegetación está representada por matorrales desérticos con una alta incidencia de los de tipo rosetófilo.

En verano de 2012 se seleccionaron dos áreas de matorral desértico rosetófilo con características muy similares (Cuadro 1), pero difiriendo en el tipo de suelo. De acuerdo con la cartografía oficial del área (Anónimo, 2005) y con su verificación en campo, los tipos de suelo encontrados fueron E+I/2 L (Rendzina + Litosol, textura media, lítico) y Rc+I/2 (Regosol calcárico + Litosol, textura media).

Evaluación de la vegetación
En cada área se establecieron 22 parcelas de muestreo rectangulares de 200 m2 (20 × 10 m) distribuidos al azar. En total fueron 44 sitios (8800 m2) en las dos áreas evaluadas, registrando las especies vegetales (suculentas, arbustivas y arbóreas) a las cuales se les midió la cobertura de copa. Se recolectó un ejemplar botánico de todas las especies cuantificadas y fueron llevados al herbario UNL de la Facultad de Ciencias Biológicas de la Universidad Autónoma de Nuevo León para su identificación y depósito.
Análisis de la información
Para cada especie se determinó su abundancia, cobertura de copa, y frecuencia. Los resultados se utilizaron para obtener el Índice de Valor de Importancia (Mueller-Dombois y Ellenberg, 1974).
Para estimar la riqueza de especies y la diversidad α de las comunidades se utilizó el índice de Margalef (DMg) y el índice de Shannon-Wiener (H´) (Magurran, 2004) mediante las ecuaciones:
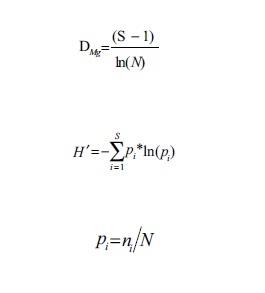
donde S es el número de especies presentes, N es el número total de individuos y ni es el número de individuos de la especie i.
Para determinar si había diferencias significativas en las variables de riqueza (índice de Margalef), diversidad (índice de Shannon-Wiener), densidad (N/ha) y cobertura de la copa (m2/ha) entre las dos áreas evaluadas, se estimaron los valores promedios de los sitios de muestreo por cada tipo de suelo y se realizó un análisis de varianza de un factor (α = 0.05) mediante el paquete estadístico SPSS Versión 15.0. Los datos que no cumplieron con las características de homocedasticidad se sometieron a una transformación logarítmica (y´ = ln (xi + 1)) (Zar, 1999).
La similitud entre los sitios de muestreo (diversidad β) se determinó por el método de Sørensen cuantitativo (IS), el cual está basado en la relación presencia-ausencia de especies compartidas en cada sitio y el número total encontrado de los dos sitios que están siendo comparados. Se expresa mediante la siguiente ecuación (Magurran, 2004):
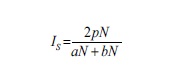
donde aN = número total de individuos en el sitio A, bN = número total de individuos en el sitio B, pN = sumatoria de la abundancia más baja de cada una de las especies compartidas entre ambos sitios.
RESULTADOS
Composición florística
Se registraron un total de 35 especies, pertenecientes a 29 géneros y 14 familias de plantas vasculares (Anexo 1). La familia Cactaceae fue la mejor representada con 16 especies, seguida por las leguminosas (Fabaceae) con cuatro. El resto de las familias tuvieron una o dos especies.
De igual forma, en el Anexo 2, se observa una disparidad ligera con respecto a los valores del IVI y de la densidad de especies por cada tipo de suelo. Por ejemplo, en suelos Rc+I/2 (Regosol calcárico + Litosol, textura media) las especies con los valores IVI y densidad más altos fueron: Larrea tridentata (23.31%; 1370 N/ha); Koeberlinia spinosa (17.01%; 623 N/ha) y Agave lecheguilla (10.17%; 1270 N/ha), mientras que en los suelos E+I/2 L (Rendzina + Litosol, textura media, lítico) los valores más relevantes los obtuvieron las especies: Agave lecheguilla (16.52%; 4414 N/ha), Larrea tridentata (13.13%; 1032 N/ha) y Leucophyllum frutescens (9.37%; 1032 N/ha).
Densidad y cobertura de copa
La densidad de individuos registrada en el suelo Rc+I/2 fue de 8909 ± 3027 N/ha (media y desviación estándar), mientras en el suelo E+I/2 L fue de 4790 ± 1555 N/ha; la diferencia es estadísticamente significativa (F = 32.20, P = < 0.000). Por su parte, la cobertura de las copas de la vegetación presente en el suelo Rc+I/2 fue de 1634 ± 678 m2/ha y del suelo E+I/2 L fue de de 1489 ± 1270 m2/ha; fue similar (F = 0.225, P = 0.638) (Fig. 2).

Riqueza y diversidad α
El índice de Margalef (DMg) para el suelo Rc+I/2 fue de 1.36 ± 0.39, mientras para el suelo E+I/2 L de 1.35 ± 0.52, sin que estas diferencias fueran estadísticamente significativas (F = 0.005, P = 0.944). De acuerdo con el índice de diversidad de Shannon (H´) el área con suelo Rc+I/2 presentó un valor de 1.55 ± 0.39 y el E+I/2 L de 1.63 ± 0.36 (Fig. 3). El análisis de varianza determinó la ausencia de diferencias significativas en la diversidad entre ambas áreas (F = 0.474, P = 0.495).

Diversidad β
De acuerdo con el coeficiente de Sorensen (IS) las áreas evaluadas presentan una similitud de 49.6%, lo que indica una semejanza media. Presentan en común 19 de las 35 especies.
DISCUSIÓN
La composición registrada corresponde a la esperada para el tipo de vegetación analizada, dada la predominancia de individuos del género Agave. Las 35 especies registradas pudieran indicar una riqueza baja en comparación con otras comunidades similares, como las estudiadas en Querétaro por Treviño-Carreón y Hernández-Sandoval (2000), quienes reportaron 83, o las de Coahuila por Encina-Domínguez et al. (2013) que registraron 97. Sin embargo, esto se puede explicar por la menor superficie de muestreo en este trabajo y la exclusión de las plantas herbáceas. La dominancia de la familia Cactaceae era de esperarse, como sucede en otros lugares donde se presenta este tipo de vegetación como el Valle de Tehuacán en Puebla y la Sierra El Doctor en Querétaro (Martorell y Ezcurra, 2002).
Ni la riqueza de especies ni la diversidad fueron significativamente diferentes entre las dos áreas estudiadas. Estos resultados no concuerdan con los reportados en otros estudios, como el de Huerta-Martínez y García-Moya (2004) en el que trabajaron con cuatro distintos tipos de matorrales con valores de diversidad (H) que variaron de 1.07 a 2.17. Sin embargo, es importante mencionar que sí hay una diferencia muy marcada en la composición florística de las dos parcelas. Es evidente que aunque las áreas tienen una similitud de casi 50%, las especies no compartidas son totalmente distintas, por lo que pueden responder entre otras cosas al tipo de suelo. Es notable que en el suelo Rc+I/2 (Regosol calcárico + Litosol, textura media) 11 de las 16 cactáceas registradas fueron exclusivas, mientras que en E+I/2 L (Rendzina + Litosol, textura media, lítico) las plantas que no se comparten son arbustos comunes como Acacia rigidula, Celtis pallida, Eysenhardtia texana y Helietta parvifolia (Anexo 2). Diversos estudios sobre las cactáceas han mencionado su preferencia por crecer en suelos de tipo regosol y/o con un alto contenido de yeso (Gómez-Hinostrosa y Hernández, 2000; Martínez-Ávalos y Jurado, 2005). El hecho de que los regosoles calcáricos sean suelos jóvenes y con poca retención de humedad (Porta et al., 1999) no favorece la presencia de plantas con mayor biomasa como los arbustos densos y altos. Por el contrario, permite el desarrollo de individuos de porte bajo y adaptados a una sequía mayor como las plantas suculentas. Este hecho también explica que la densidad de las especies fue mayor en el suelo Rc+I/2 y substancialmente menor en el E+I/2 L. Finalmente la cobertura de copa no indicó diferencias importantes a pesar de que las especies en cada área son en general distintas.
Aunque los resultados de esta investigación no coinciden con los de otros en los que se menciona que los factores ambientales influyen positivamente en la diversidad de especies en estudios de comunidades a nivel local (Huerta-Martínez y García-Moya, 2004), sí es importante señalar que otros aspectos de la vegetación son afectados en gran medida por ciertos factores. Esta idea ya se ha hecho notar en otros trabajos, algunos de los cuales han puesto de manifiesto la importancia de las características del suelo o substrato como la profundidad y la pendiente (Abd, 2000; Gallego, 2003; Rankin et al., 2007; Santibáñez-Andrade et al., 2009).
Los suelos representan un papel importante en la abundancia y composición florística de los matorrales estudiados, tomando en cuenta que otros factores físicos como el clima, la elevación y la exposición son similares. Es evidente que los suelos de tipo regosol calcárico son preferidos por las cactáceas, porque permiten un mayor drenaje que favorece un hábitat adecuado para estas especies.
AGRADECIMIENTOS
A la consultoría ambiental Gestión Estratégica y Manejo Ambiental S.C. por el apoyo en las actividades de campo. La investigación fue financiada parcialmente por el proyecto PROMEP/103.5/12/3585.
LITERATURA CITADA
Abd, M. M. 2000. Floristics and enviromental relations in two extreme desert zones of western Egypt. Global Ecol. Biogeogr. 9: 499-516. [ Links ]
Alanís, E., J. Jiménez, O. A. Aguirre, E. J. Treviño, E. Jurado y M. A. González. 2008. Efecto del uso del suelo en la fitodiversidad del matorral espinoso tamaulipeco. Ciencia UANL. 1: 56-62. [ Links ]
Anónimo. 2005. Suelos. Compendio de la situación del medio ambiente en México. Compendio de estadísticas ambientales. Secretaría de Medio Ambiente y Recursos Naturales. México, D.F., México. [ Links ]
Bautista, F., A. J. Zinck y S. Cram. 2009. Los suelos de Latinoamérica: retos y oportunidades de uso y estudio. Instituto Nacional de Estadística, Geografía e Informática. Bol. SNIEG. 2(3): 93-142. [ Links ]
Briones, O. y J. Á. Villarreal. 2001. Vegetación y flora de un ecotono entre las provincias del Altiplano y de la Planicie Costera del Noreste de México. Acta Bot. Mex. 55: 39-67. [ Links ]
Canizales, P. A., E. Alanís, R. Aranda, J. M. Mata, J. Jiménez, G. Alanís, J. I. Uvalle y M. G. Ruiz. 2009. Caracterización estructural del matorral submontano de la Sierra Madre Oriental, Nuevo León. Rev. Chapingo. Ser. cienc. for. amb. 15: 115-120. [ Links ]
Encina-Domínguez, J. A., J. A. Meave y A. Zárate-Lupercio. 2013. Structure and woody species diversity of the Dasylirion cedrosanum (Nolinaceae) rosette scrub of central and southern Coahuila State, Mexico. Bot. Sci. 91(3): 335-347 [ Links ]
Estrada-Castillón, E., J. Á. Villarreal-Quintanilla, E. Jurado-Ybarra, C. Cantú-Ayala, M. A. García-Aranda, J. Sánchez-Salas, J. Jiménez-Pérez y M. Pando-Moreno. 2012. Clasificación, estructura y diversidad del matorral submontano adyacente a la planicie costera del Golfo Norte en el Noreste de México. Bot. Sci. 90: 37-52. [ Links ]
Gallego, J. B. 2003. Distribución de especies de matorral en suelos ácidos y básicos de la Sierra de Grazalema, Sur de España. Anales Jard. Bot. Madrid 60: 51-61. [ Links ]
Gómez-Hinostrosa, C. y H. M. Hernández. 2000. Diversity, geographical distribution, and conservation of Cactaceae in the Mier y Noriega region, Mexico. Biodivers. Conserv. 9: 403-418. [ Links ]
González-Medrano, F. 1996. Algunos aspectos de la evolución de la vegetación de México. Bol. Soc. Bot. Méx. 58: 129-136. [ Links ]
González-Medrano, F. 2004. Las comunidades vegetales de México. Propuesta para la unificación de la clasificación y nomenclatura de la vegetación de México. Instituto Nacional de Ecología, Secretaría de Medio Ambiente y Recursos Naturales. 2a. ed. México, D.F., México. 82 pp. [ Links ]
Huerta-Martínez, F. y E. García-Moya. 2004. Diversidad de especies perennes y su relación con el ambiente en un área semiárida del centro de México: implicaciones para la conservación. Interciencia 29: 435-444. [ Links ]
Jiménez-Pérez, J., E. Alanís, M. A. González, O. A. Aguirre y E. J. Treviño. 2013. Characterizing woody species regeneration in areas with different land history tenure in the Tamaulipan thornscrub, Mexico. Southwest. Nat. 58(3): 299-304. [ Links ]
Jiménez-Pérez, J., E. Alanís, O. A. Aguirre, M. Pando y M. A. González. 2009. Análisis sobre el efecto del uso del suelo en la diversidad estructural del matorral espinoso tamaulipeco. Madera y Bosques 15: 5-20. [ Links ]
Magurran, A. 2004. Measuring biological diversity. Blackwell Science. Oxford, UK. 256 pp. [ Links ]
Martínez-Ávalos J. G. y E. Jurado. 2005. Geographic distribution and conservation of Cactaceae from Tamaulipas, Mexico. Biodivers. Conserv. 14: 2483-2506. [ Links ]
Martorell, C. y E. Ezcurra. 2002. Rosette scrub occurrence and fog availability in arid mountains of Mexico. J. Veg. Sci. 13: 651-662. [ Links ]
Mueller-Dombois, D. y H. Ellenberg. 1974. Aims and methods of vegetation ecology. John Wiley. Nueva York, USA. 547 pp. [ Links ]
Palacio, J. L., G. Bocco, A. Velásquez, J. F. Mas, F. Takaki, A. Victoria, L. Luna, G. Gómez, J. López, M. Palma, I. Trejo, A. Peralta, J. Prado, A. Rodríguez, R. Mayorga y F. González-Medrano. 2000. La condición actual de los recursos forestales en México: resultados del inventario forestal nacional 2000. Bol. Inst. Geogr. 43: 183-203. [ Links ]
Porta, J., M. López-Acevedo y C. Roquero. 1999. Edafología para la agricultura y el medio ambiente. Mundi-Prensa. Madrid, España. 849 pp. [ Links ]
Rankin, M. O., W. S Semple, B. W. Murphy y T. B. Koen. 2007. Is there a close association between 'soils' and 'vegetation'? A case study from central western New South Wales. Cunninghamia 10: 199-214. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Editorial Limusa, México. 432 pp. [ Links ]
Santibáñez-Andrade, G., S. Castillo-Argüero, J. A. Zavala-Hurtado, Y. Martínez-Orea y M. Hernández-Apolinar. 2009. La heterogeneidad ambiental en un matorral xerófilo. Bol. Soc. Bot. Méx. 85: 71-79. [ Links ]
Treviño-Carreón, J. 2004. Ecología de los matorrales rosetófilos de México: patrones geográficos y ecofisiológicos de las comunidades de Dasylirion. Tesis de doctorado. Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro. Querétaro, México. 164 pp. [ Links ]
Treviño-Carreón, J. y L. G. Hernández-Sandoval. 2000. Introducción a los matorrales rosetófilos de Querétaro, México. Memorias de los Mejores Trabajos del Simposio 2000. La Investigación y el Desarrollo Tecnológico en Querétaro. Consejo de Ciencia y Tecnología de Querétaro. Querétaro, México. pp. 16-25. [ Links ]
Ugalde, J., D. Granados-Sánchez y A. Sánchez-González. 2008. Sucesión en el matorral desértico de Larrea tridentata (DC.) Cov. en la Sierra de Catorce, San Luis Potosí, México. Terra Latinoamericana 26: 153-160. [ Links ]
Zak, D. R., W. E. Holmes, D. C. White, A. D. Peacock y D. Tilman. 2003. Plant diversity, soil microbial communities, and ecosystem function: are there any links? Ecology 84: 2042-2050. [ Links ]
Zar, J. 1999. Biostatistical analysis. 4ta ed. Prentice-Hall. Upper Saddle River, USA. 663 pp. [ Links ]

