










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa botánica mexicana
versión On-line ISSN 2448-7589versión impresa ISSN 0187-7151
Act. Bot. Mex no.100 Pátzcuaro jul. 2012
Assessing introduced Leguminosae in Mexico to identify potentially high-impact invasive species*
Evaluación de leguminosas introducidas en México para identificar especies invasoras de alto impacto potencial
Judith Sánchez-Blanco1, Clara Sánchez-Blanco1*, Mario Sousa S.2 y Francisco J. Espinosa-García1,3
1 Universidad Nacional Autónoma de México, Centro de Investigaciones en Ecosistemas, Antigua Carretera a Pátzcuaro 8701, Colonia Ex Hacienda de San José de la Huerta, 58089 Morelia, Michoacán, México.
2 Universidad Nacional Autónoma de México, Instituto de Biología, Departamento de Botánica, Ciudad Universitaria, 04510 México, D.F., México.
3 Author for correspondence:espinosa@oikos.unam.mx
Recibido en febrero de 2012.
Aceptado en mayo de 2012.
ABSTRACT
Most introduced plant species in Mexico have not been assessed to prioritize management actions to eradicate or contain them, or for damage mitigation after plant invasions. We assessed 42 introduced weed taxa in the Leguminosae using empirical and theory derived criteria in a model focused in the species behavior abroad, their residence time and number of occupied localities, and the presence in Mexico of closely related relatives (native and non-native) at the genus level. Data were obtained from bibliographic sources and from the "Malezas Introducidas en México" database, which includes information from 11 Mexican herbaria. We also developed a scoring process to qualify introduced weed expansion using residence time and number of occupied localities. We classified the analyzed introduced legumes in four priority of attention categories. We suggest that Albizia lebbeck, Pueraria phaseoloides, Lablab purpureus, Securigera varia, Delonix regia, Clitoria ternatea, and Spartium junceum should receive high-priority attention; eight species were considered to require medium priority attention; seven low priority attention, and 20 taxa were classified as non-priority. The developed assessment model still needs further refinement, as seemingly innocuous species scored high and a potentially dangerous species (Cassia fistula) were classified in the non-priority category. We hope that this assessment model will work as a structured, low expert-dependent approach to identify the introduced species that require a further risk analysis to prioritize efforts for noxious plant management.
Key words: introduced weeds, invasive plants, naturalized species, residence time, risk analysis.
RESUMEN
La mayoría de las especies de plantas introducidas en México no han sido evaluadas para priorizar las acciones de gestión para erradicarlas o contenerlas o para mitigar los daños causados por invasiones de plantas nocivas. Se evaluaron 42 taxa (41 especies y una variedad) de malezas introducidas de la familia Leguminosae usando criterios empíricos y derivados de teoría, en el desarrollo de un modelo centrado en el comportamiento de las especies en el extranjero, su tiempo de residencia y las localidades ocupadas, y la presencia en México de los parientes congenéricos nativos y no nativos. Los datos se obtuvieron a partir de fuentes bibliográficas y de la base de datos "Malezas Introducidas en México" que incluye información de 11 herbarios de México. También desarrollamos un proceso de puntuación para calificar la expansión de las introducidas de acuerdo con tiempo de residencia y número de localidades ocupadas. Clasificamos a las leguminosas introducidas en cuatro categorías de prioridad de atención. Albizia lebbeck, Pueraria phaseoloides, Lablabpurpureus, Securigera varia, Delonix regia, Clitoria ternatea y Spartium junceum deben recibir prioridad de atención alta; consideramos que ocho especies requieren una atención prioritaria media, y siete atención prioritaria baja. 20 taxa fueron clasificados como no prioritarios. El modelo de evaluación desarrollado todavía necesita un mayor refinamiento, ya que especies aparentemente inocuas obtuvieron puntajes altos y una potencialmente peligrosa (Cassia fistula) se clasificó en la categoría no prioritaria. Esperamos que el presente modelo de evaluación funcionará como una herramienta y con baja dependencia de expertos, para identificar a las especies introducidas que requieren un análisis de riesgo más completo para priorizar esfuerzos para el manejo de las plantas introducidas nocivas.
Palabras clave: análisis de riesgo, especies naturalizadas, malezas introducidas, plantas invasoras, tiempo de residencia.
INTRODUCTION
The invasion of natural communities by introduced species is one of the most serious threats to biodiversity and human economic interests, because invasive species displace or replace native species and cause large losses in crop and livestock agroecosystems (Holm et al., 1991; Vitousek et al., 1996; Lonsdale 1999; Lockwood et al., 2001; Vilá et al., 2011). Although very few incoming species in a new area become problematic (Williamson and Fitter, 1996), the prediction of which species will be noxious is not yet accurate enough. This occurs in spite of intense research in characteristics of successful invasive species, characteristics of invaded areas, and the interaction among invasive species and invaded environments (Blackburn et al., 2011).
The arrival of exotic species into new territory, intentionally or inadvertently, is by trade and the constant movement of people and plant and animal products (Mack, 1996; Dehnen-Schmutz et al., 2007; von der Lippe and Kowarik, 2008). The risk that an introduced plant can escape, stabilize, and spread, depends on several interacting biological, environmental, and anthropogenic factors that restrict the final number of successful invaders (Wilson et al., 2007). When any exotic plant species establishes, it often takes a long time before it causes a problem ("lag phase"); this period varies from less than 20 years in annuals to over 300 for woody plants (Williamson, 1996; Groves et al., 2005). During the lag phase, evolutionary processes enable the species' following explosive range expansion ("expansive phase") (Williamson, 1996). Problems caused by invasive plants are usually detected at the expansive phase, but at this stage eradication is unviable and only very expensive contention or mitigation actions may be available (Rejmánek and Pitcairn, 2002). Thus, the identification of potentially dangerous introduced plant species becomes imperative besides the identification of exotic species to be excluded from entrance in a new territory. We use "introduced" to refer to exotic species growing without direct human help in natural or managed ecosystems.
Risk assessment models use empirical and theoretically derived criteria that can be useful in assessing introduced species. For example, invasive plant behavior (in the original and introduced ranges) and residence time in the new area, have been used to predict invasions and manage exotic or introduced species (Panetta and Mitchell, 1991; Mack et al., 2000; Meyer, 2000; Rejmánek, 2000; Heger and Trepl, 2003; Essl, 2007; Ricciardi and Cohen, 2007; Wilson et al., 2007; von der Lippe and Kowarik, 2008). In addition, useful exotic species become invasive more easily than non-useful ones, due to assisted propagation, pest protection, and increased propagule pressure (Williamson, 1996). Thus escaped crops, forages, ornamentals, and other horticultural species, account for more than 50% of naturalized species in several parts of the world (Grotkopp et al., 2010). Other important empirical criteria useful in predicting invasions, are unavailable from herbarium based information, for example, seedling growth rate, genome size, minimum generation time, and relative growth rate, among others (Grotkopp et al., 2002; Grotkopp et al., 2010).
Some hypotheses proposed to explain biological invasions can be useful to derive applied criteria in identifying potentially noxious invasive species. The "biotic resistance" proposes that native species in the community resist invasion by occupying all available niches leaving no space for newcomers, thus a negative relationship between native and introduced biodiversity is expected (Elton, 1958; Heger and Trepl, 2003). The "taxonomic affinity" or "phylogenetic repulsion" hypothesis (Mack, 1996; Lockwood et al., 2001; Strauss et al., 2006) predicts that the invasion success of a plant is inversely related with the number of close taxonomic relatives in the community to invade. A practical criterion derived from these hypotheses would be to qualify as more dangerous those exotic species expected to invade low diversity communities and/or those without close relative native species. However, evidence for both hypotheses has been inconclusive, thus they have not used firmly in predicting invasions (Espinosa-García et al., 2004a; Strauss et al., 2006).
Thus, although there are several empirical and theoretical approaches to explain biological invasions, there are practical limitations to predict which species will become noxious (Heger and Trepl, 2003; van Kleunen et al., 2010). This is a problem as efforts to control and/or eradicate exotic species are limited for the many introduced species established in a region, thus priorities have to be set (Randall et al., 2001).
After making an inventory of introduced species, one of the first steps to prioritize their management is to analyze their origin, introduction, distribution, and residence time (Huang et al., 2009; Wu et al., 2003), together with behavior abroad (Randall, 2002). For Mexico, in addition to studies of inventory and distribution patterns of introduced weeds at the national scale (Villaseñor and Espinosa-García, 2004; Espinosa-García et al., 2004a; 2004b; 2009), there are very few studies on distribution, abundance or importance for most introduced species the country (Espinosa-García et al., 2009). All those tasks are necessary in accordance with the National Strategy for Invasive Species in Mexico (Anonymous, 2010a). Although national experts identified 23 invasive plant species of potentially high impact to biodiversity in Mexico, they did it without a formal risk analysis (March Mifsut and Martínez Jiménez, 2007). Furthermore, most of the species mentioned in that publication are not present in the country or their presence in Mexico is only suspected. Thus, the risk assessment for more than 600 introduced species in Mexico (Espinosa-García et al., 2009) is pending. However, a full risk analysis (including potential distribution modeling) is a very difficult task for so many species. Thus, in this paper we are proposing a low expert-dependent assessment model to detect the potentially dangerous species to be assessed with a full risk analysis.
The purpose of this paper is to assess introduced legumes in Mexico with empirical and theory-derived criteria to identify potentially high-impact species using herbarium and bibliographic data. The family Leguminosae (s.l.) (Cronquist, 1981) has approximately 650 genera and 18,000 species worldwide; it is one of the most species-rich families in local floras. It occupies the third place with in the global exotic flora after Poaceae and Asteraceae (Pyšek, 1998). In Mexico, Leguminosae is the second most important family with 130 genera and approximately 1800 species (Sousa and Delgado, 1993); it also ranks second in importance in the flora of alien weeds with 36 genera and 57 species, representing 9.2% of introduced weeds (Villaseñor and Espinosa-García, 2004). This family has contributed the most damaging introduced weeds in Australia (Emms et al., 2005).
We expect that this assessment will serve as a first approach model to indicate the introduced species that require more extensive risk assessments in order to focus eradication, control, containment or mitigation efforts. We used empirical criteria on distribution, residence time, invasive behavior, and usage detailed in the methods sections. We obtained theory-derived criteria testing the hypothesis of phylogenetic repulsion; a negative relationship between native and introduced weed species within the same genus was expected. All species were scored for each criterion. The scoring for criteria was inspired in those used in various weed risk assessment models (e.g. Pheloung, 1999; Koop et al., 2012).
MATERIALS AND METHODS
The analysis was based on the 42 introduced legume species included in the "Malezas Introducidas in México" (MIM) database (Espinosa-García et al., 2000). Twenty-nine introduced legume species added to the introduced flora of Mexico after 2000, and therefore absent in the MIM database, were not included in this analysis (Appendix). We used the records in the MIM database obtained from 1164 specimens in the 11 Mexican herbaria, with the best representation of the flora of Mexico (Table 1).
For each herbarium specimen data were taken on locality (municipality and state), collection date, life form characteristics, and uses. Data were collected on the geographical origin, climatic affinity, and year of its first record in herbarium to estimate residence time for each species. Data not found on the specimens labels were obtained through literature or online searches. We verified the identification of each specimen using reliable botanical descriptions and diagnostic traits obtained from various sources (Espinosa-García et al., 2000)
We used the number of native and introduced weed species within the same genus to determine whether a negative correlation between these groups of weeds occurred as predicted by the phylogenetic repulsion hypothesis. To this end we obtained a list of native weed species in the Leguminosae from the Weed Catalog of Mexico (Villaseñor and Espinosa-García, 1998). Native non-weed species were excluded because the disturbed environments colonized by introduced species are those occupied by native weeds. In those environments, native non-weed species are frequently absent and biotic interactions occur mainly between weedy groups (Espinosa-García et al., 2004a).
Criteria for the assessment of introduced species
The evaluation of the naturalized species requires a risk analysis similar to those used to assess the ban on entry of a species to a country. Naturalized species are assessed for current or potential damage to natural or modified ecosystems. The assessment is done through a number of attributes related to their behavior in their places of origin and their introduced ranges; the presence of close relatives in the settled area and the presence invasive close relatives in other parts of the world; its residence time, and the number of localities and habitats occupied in the invaded country. Australians have rating scales for naturalized species according to their importance and current and potential effects on wild and managed ecosystems (Groves et al., 2005). In Mexico, knowledge of the effects of naturalized species is very limited and information is available for very few species (Espinosa-García et al., 2009), making it impossible to apply the Australian scales to assess weeds naturalized in Mexico. Thus, we only used herbarium and literature information as a first approach to prioritize species to avoid potential problems.
Scores were assigned to species according to a) behavior abroad; b) residence time and expected distribution; c) number of habitat types occupied reported in Mexico; d) growth form; e) number of climate types in which it was detected; f) presence of native weeds and invasive relatives within the same genus. Scores for each item were assigned as described below.
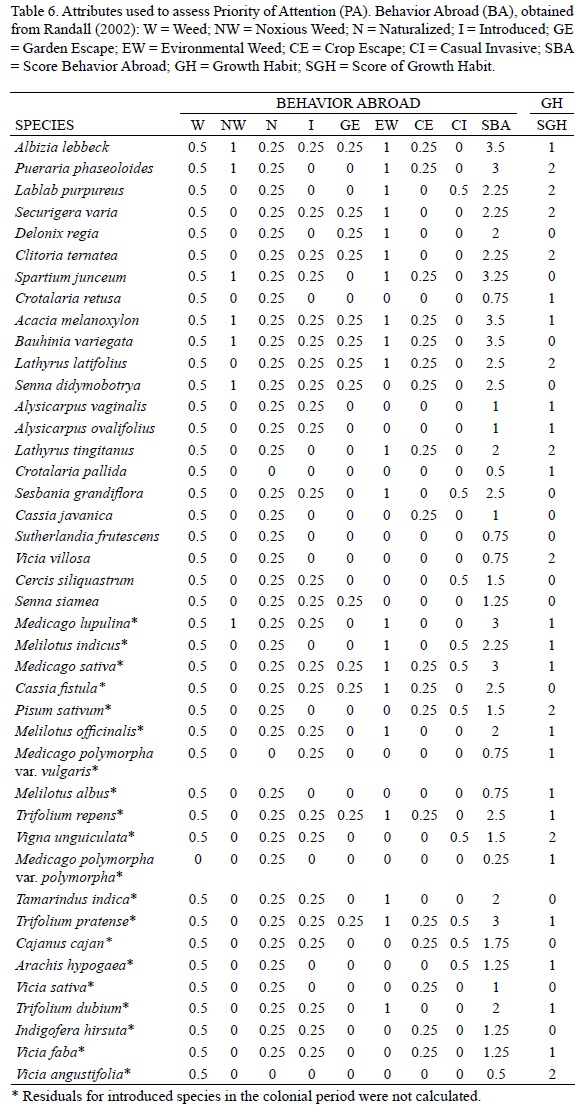
For the first criterion, we classified the introduced legumes' behavior in the world according to the World Weed Compendium (Randall, 2002), assigning values for each category: W (Weed) = 0.5; NW (noxious weed) = 1, N (Naturalized) = 0.25, I (Introduced) = 0.25; GE (Garden Escape) = 0.25; EW (Environmental Weed) = 1, EC (Escaped Crop) = 0.25, CI (Casual Invasive) = 0.5.
For the second criterion, we assigned scores according to the magnitude of the residuals of the equation for expected values obtained by correlating residence time and number of localities recorded for all species (Fig. 2b). The rating was calculated according to the deviation of the observed data from the confidence limits of the expected values for each species. The score was 0 if the residuals for the species fell within the confidence limits of the regression line (Fig. 2b). If the residual exceeded the value of the upper confidence limit one to two times, the score was 1; if the residual exceeded two to three times the upper confidence limit, it was 1.5; if the residual exceeded the limit four or more times, the score was 2. For residual values smaller than the lower confidence limit, the score was -1, if the residual was two to three times smaller than the lower confidence limit, the score was -1.5, and if the residual was four or more times smaller than the lower confidence limit, the score was -2.
For the third criterion (habitat breadth), we assigned a value according to the number of habitat types where the species has been recorded. The score was 0 for species recorded in a single habitat type; for species reported in two to four habitat types, 2; the score was 3 if the number of reported habitat types were five to seven, and 3 when they were eight to eleven habitat types. We recognized 11 habitat types: wet areas (WA), disturbed areas (DA), ruderal (R), secondary or disturbed vegetation (SVD), freshwater (FW), shores of salty water (SSW), cropland for annuals (CA), cropland for perennials (CP), natural vegetation (VN), livestock areas (LA) and gardens and meadows (GM). This criterion was based on Bradshaw et al. (2008), who found that in legumes, herbs and vines occupying multiple habitat types were more likely to become invasive.
For the fourth criterion, we assigned values to life forms: 0 for woody perennials (trees and shrubs); 1 for herbaceous plants, and 2 for vines or climbing herbs. Growth habit has been associated with serious damage, mainly with vines or climbing plants (Phillips et al., 2010) or with leguminous trees (Emms et al., 2005).
In the fifth criterion, we assigned 0 to the species occupying areas with one kind of climate and 1 when occupying two or more. We only were able to consider broad climate type categories due to the scarcity or vagueness of climate data on herbarium specimen labels, tropical (A in Köppen's classification), dry (Köppen's B), temperate (Köppen's C), and cold (Köppen's H).
For the sixth criterion we scored -1 for introduced species with close native relatives (within the same genus); a score of 1 to species without close native relatives; 1 for introduced species with introduced relatives, and 0 to introduced species without introduced relatives. These scores were assigned after the analysis performed with introduced and native luminous weeds, where we found patterns (see the results section) mostly consistent with the phylogenetic repulsion hypothesis (Mack, 1991; Lockwood et al., 2001; Strauss et al., 2006).
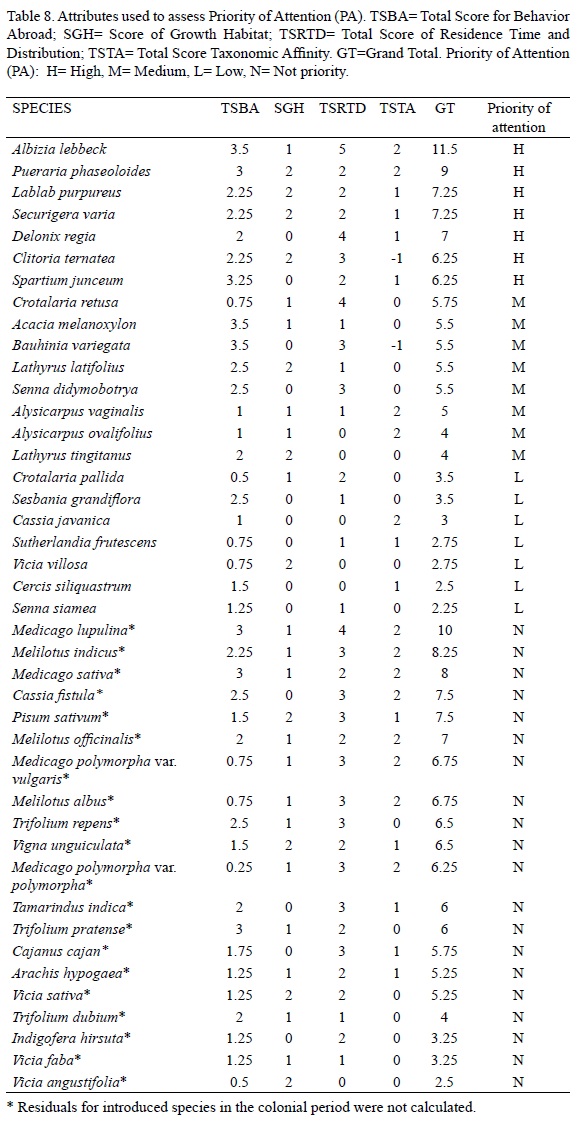
We correlated the partial sums for criterion groups (behavior abroad, residence time and distribution, and taxonomic affinity) to determine if they were redundant. The species were classified according to the sum of their scores: low-priority attention (L), 1 to 3.5; medium priority (M), 4 to 6.25, and high priority (H), 6.5 to 11.5. The scores for all criteria were based on non-proportional ordinal scales to estimate qualitatively the characteristics used for risk assessment (Randall et al., 2001; Ricciardi and Cohen, 2007).
RESULTS AND DISCUSSION
Introduced legumes are a very diverse group in terms of taxonomic composition, origins, uses and distribution in Mexico (Table 2). The 42 analyzed taxa are distributed in nine genera in three subfamilies: Caesalpinioideae (eigth species); Papilionoideae (31 species and one variety), and Mimosoideae (two species). The leguminous introduced species not considered in this analysis include eight in Caesalpinioideae, 19 in Papilionioideae, and four in Mimosoideae (Appendix). Thus, the subfamily Mimosoideae is strongly underrepresented in the MIM database, as 66.7% of the known introduced species in Mexico for this group were not included there.
Today Mexican herbaria have the best collection of plant specimens collected in Mexico. However, all herbaria, except the Herbario Nacional (MEXU), have information mainly from the 1960's to the present. Although MEXU herbarium was formed with three late nineteenth century herbaria, most specimens in its collection were obtained during and after the 1970's. Mexico's historical collections are in European and United States herbaria; the oldest studied is that of W. Houstoun (with 1731 Veracruz and Campeche specimens), deposited in the British Museum (BM) (Rzedowski et al., 2009). The most complete collection of Mexican plants collected before the 1960's is at the Smithsonian Institution herbarium (US) in Washington. This herbarium has nearly 100,000 Mexican specimens, most of them without duplicates in Mexican institutions. We were not able to revise these foreign herbaria. Therefore, residence times are underestimated, especially for species naturalized in Mexico before 1950; it is difficult to know the degree of underestimation, as many botanists (especially in the first half of the 20th century and before) purposefully ignored weeds or disturbed environments. By the time that "Malezas Introducidas en México" (MIM) database was obtained (Espinosa-García et al., 2000), we estimate that the 11 revised herbaria had more than 2,200,000 specimens, but for 2012, they had more than 2,900,000 (Table 1). Herbaria other than MEXU, which has the best representation of the Mexican flora, were selected for their regional representation, especially when they were located in large agricultural regions or at agronomy universities, e.g. CHAPA, ANSM, UAS (Table 1). Thus, the MIM database is a representative sample of weeds in Mexico.
We will describe and discuss some general features for the analyzed species before we continue to the results and discussion for scoring and evaluation.
Origin
Most analyzed species (95%) come from the Old World (Africa, Asia and Europe), and the rest come from the Americas or Oceania (Tables 1, 2). Most species in the subfamily Papilionoideae originated in the Old World; their herbarium records span are 6 to 100 years old. The Mimosoideae have origins in Oceania and Asia, with early herbarium records from 60 years ago. The Caesalpinoideae come from the Old World and South America, and is the most recently detected subfamily according to their herbarium records (Table 1).
Bibliographic sources sometimes refer to origins in specific continents or regions, and frequently in "the Old World" thus, it is not possible to make a thorough analysis as to the origin of most species. The predominance of Old World species could be explained because during the colonial period Spaniards traded and exchanged plants with Europe and Asia. During this time the main supply bases were the Española Island and Cuba, which in turn received commodities from Europe and Africa (Challenger, 1998). The Nao of China was another important way for species introductions from eastern Asia that were transported to New Spain via Acapulco. The Spaniards also introduced livestock, fodder and crops intentionally, and accidentally, plant propagules as stowaways on animals or fodder and as contaminants in crop seeds. Propagules are still introduced accidentally to Mexico in grain and seeds shipments from North America, South America and Europe (Calderón and Espinosa-García, 1997).
Use
All analyzed species have at least one use (Table 2), which suggests that most of them were introduced purposefully into Mexico. The main uses for introduced Caesalpinoideae species are ornamental, medicinal, forage and food (Table 3). Most forage species are in the Papilionoideae (65.6%); ornamentals are in the Mimosoideae (2 species only), Caesalpinoideae (100%) and Papilionoideae (28.1%). Medicinal legumes represent 19.1% and the food species 21.4%. Legumes used as green manure or that are toxic are rare (Table 2. Several species have multiple uses, for example Tamarindus indica, whose fruit pulp is used to make syrups, preserves, juice concentrates, sweets, and sauces. This tree is also ornamental and medicinal; its seeds are used for necklaces, earrings and crafts. Other species such as Arachishypogaea, Lablab purpureus, Sesbania grandiflora, Spartium junceum, and Viciafaba are also species with more than one use (Table 2).
The usefulness of these plants has probably been important in their introduction, facilitating their establishment and dispersal in the country. They are imported, planted in different places and then these plants escape into the wild (Emms et al., 2005; Huang et al., 2009). In South Africa, introduced ornamentals disseminated faster than introduced species with other uses (Wilson et al., 2007). Another advantage for introduced ornamentals and crops is the continued introduction of new genetic material, hybrid formation and selection of new varieties, which increase the likelihood of adapted local biotypes (Essl, 2007; Wilson et al., 2007). In Britain, the ornamental species introduced by the horticultural industry have had a major impact on the invasions, as they are favored by frequent low prices and high market availability, which translates into a large propagule pressure (Dehnen-Schmutz et al., 2007).
Climate affinity
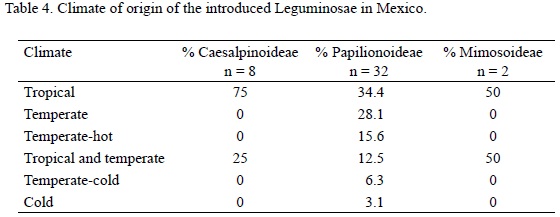
Introduced legumes in Mexico are mainly from tropical climates. All species of the subfamily Caesalpinoideae and a third of Papilionoideae originated in areas with these climate conditions. Few species in the Mimosoideae and Caesalpinoideae grow in both tropical and temperate climates. The Papilionoideae grow in a wider range of climatic conditions than the other two subfamilies (Anonymous, 1979) (Table 4). The distribution of the introduced legumes in Mexican climatic areas agrees with the homoclimate hypothesis (Panetta and Mitchell, 1991) as species of tropical origin have been registered in tropical areas of Mexico, and those of temperate origin in temperate areas the country (Sánchez-Blanco, 2003).
Taxonomic affinity and the phylogenetic repulsion hypothesis
In Mexico legume weeds are represented by approximately 159 native species (Villaseñor and Espinosa-García, 1998) and 72 introduced species (41 in the MIM database and 31 in the appendix). The subfamily of legumes best represented in both introduced and native weeds is Papilionoideae, with 69.4% and 66.7% respectively, followed by Caesalpinoideae (22.2% introduced, 12.6% native) and Mimosoideae (8.3% introduced, 20.7% native). Legume weeds are found in 75 genera in Mexico, including 12 that share native and introduced species (Table 5).

Of the legume genera reported in Mexico, 52% have only native weed species, 32% have introduced weeds only, and those with both types of species represent 16%. The fact that most genera (63) have either native or introduced weeds is consistent with the phylogenetic repulsion hypothesis, which predicts that introduced species success correlates negatively with the presence of related native species. According to this hypothesis, we should expect 100% of the genera have only introduced or native species. Our data indicate that there are 12 genera with both types of species. Even with this number of genera with species of both types, the obtained distribution differs from the null hypothesis in which the expected frequency would be equal proportions for the three genera categories (χ2 = 19.53, df 2, p <0.0001).
For genera with both types of species, the phylogenetic repulsion hypothesis predicts an inverse relationship between the frequency of native and introduced species in the same genus. There is no detectable pattern in the incidence of native and introduced within the same genus (Spearman rank correlation r = 0.26, p = 0.86). In Crotalaria, Acacia, and Mimosa, native species prevail clearly with a ratio native/introduced of 3 to 9 (Table 5). In five genera (Caesalpinia, Indigofera, Lathyrus, and Vicia) introduced species prevail over the natives, although in smaller proportions (0.5 to 0.33). Possibly, the number of legume genera with both types of weed species was not large enough to test the phylogenetic repulsion hypothesis adequately. This hypothesis has been tested at the scale habitat or communities. For example, Strauss et al. (2006) found that in California grasslands, the introduced Poaceae species with high ecological impact are phylogenetically more distant to native grasses than the low impact introduced grasses. Another possibility to explain the failed test is that native and introduced species within the same genus are not occurring within the same plant community, a pattern that we could not detect with our data.
Overall, we found that most legume genera either have native or introduced weed species. This is consistent with the phylogenetic repulsion hypothesis and the findings made in floras of different areas of the United States (Mack, 1991; Lockwood et al., 2001; Strauss et al., 2006).
Residence time and distribution
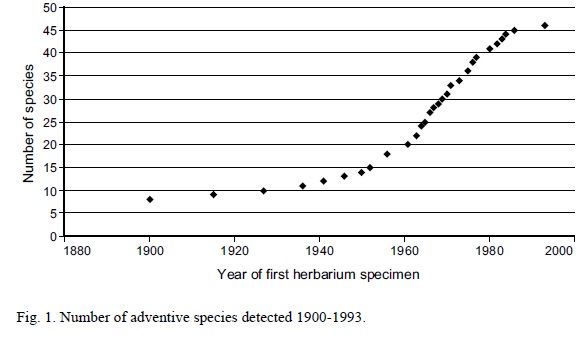
Specimens of introduced Leguminosae in Mexican herbaria date from 1900 to 1993. The year with most first records was 1900 (8 species) and then few records of new species appeared until 1960 (Fig. 1). From the late 1960's to the 1980's there is a steep linear increase in the number of newly detected introduced species, which apparently stopped in 1993. This pattern probably reflects the botanical activity in Mexico more than the species introduction rate. A similar pattern was found in Taiwan, where there were two periods of increased introduced detection (1910-1930 and 1970-2000) with no species detected from 1930 to 1960 (Wu et al., 2003).
Massive plant introductions occurred in Mexico during the colonial period (Challenger, 1998), but we have no precise information on the introduction date for each species. For example, the Spaniards must have introduced many species, deliberately or accidentally, with their cattle, crops and agricultural technology (Challenger, 1998). The species that very likely were introduced in this period are identified in the following residence time analysis. As most of the collections of weeds in Mexico are recent (1950's to present), and foreign herbaria with historical Mexican plant collections were not included in this study, it is likely that residence time and accumulation rate are underestimated, especially for species naturalized in Mexico before the twentieth century.
The 1900 specimens represent forage species or those associated with livestock or crop seeds; they probably were introduced from the sixteenth to eighteenth centuries. Apparently, most ornamental species were introduced after the forage plants in the early 1950. This is consistent with reports by Dehnen-Schmutz et al. (2007) in Britain, where introduced species are mostly ornamental. The pattern of ornamental introduced species accumulation detected from herbarium specimens in South Africa is different from what we found; there, exponential accumulation of the ornamental species occurred around 1840 and probably ended in 1950 (Wilson et al., 2007).
Very few new legume introduced species were detected in Mexico from 1993 to 2002 (Espinosa-García et al., 2009, Appendix). This contrasts with the pattern found in Australia, where the introduction of species increased 10% per year (Groves et al., 2005). The scarcity of new records in that period could be due to a reduced weed collection, a long time elapsed from specimen collection, to inclusion in herbaria, a very slow new species introduction rate, or all of the above.
The correlation between the number of localities for a species and the years since its first herbarium record is significant (r = 0.39, p = 0.012) (Fig. 2a). This suggests that the earlier the first record, the wider the distribution. However, there are many species outside of the model confidence limits, some with more localities than the expected and vice versa. Most species have residence times of 22 to 47 years and occupy up to 50 localities (Fig. 2a). We suggest that many species with wider distribution than expected most likely were introduced during the colonial period (sixteenth to eighteenth centuries). These early introductions are not documented in herbaria with specimens, and their modern specimen's dates distort the regression analysis. Therefore, we decided to make the residence time analysis only for post-colonial introduced species.
The residence time correlation with the number of localities for species introduced between the 16th and 18th (Table 1) was not significant. Apparently, this group of herbaceous species did not become problematic after more than 200 years of its introduction. This time is much longer than the duration of the noxious invasives lag phase ranging from 20 to 54 years in herbs and from 80 to 300 years trees (Mack, 1981; Kowarick, 1995; Groves, 2006). Invasive neophytes in Spain occupied their potential distribution completely in 143 years on average (Gasso et al., 2010), which is consistent with the 150 years (or possibly twice) estimated in Ireland, Britain Germany and the Czech Republic (Williamson et al., 2009). In tropical climates the lag phase time between introduction and first evidence of dispersion for 23 species in Hawai'i, was 5 years for herbs and 14 years trees (Daehler, 2009).
For the species introduced after the 18th century, the correlation for residence time and number of localities was significant (r = 0.53, p = 0.012) (Fig. 2b). The R2 of this regression model is 0.28, thus, the variation explained by residence time is 28%. This value is not as high as 39 and 44% obtained by Ahern et al. (2010) in Michigan and California, but it is higher than 23% the obtained by Wu et al. (2003).
The increase in number of localities according to residence time is a well established pattern in plant invasions (Rejmánek and Richardson, 1996). Our results agree with this pattern (Fig. 2b) and those obtained with South African plants (Wilson et al., 2007); with invasive ornamental species in Great Britain (Dehnen-Schmutz et al., 2007), and introduced leguminous species in Taiwan (Wu et al. 2003). However, some introduced legumes in Mexico with short residence times now occur in many states; other species, despite having longer residence time, have been reported in few localities (Fig. 2b). Wilson et al., (2007) and Wu et al. (2003) also reported these kinds of discrepancies.
Species within the confidence limits in the regression with few occupied localities (Fig. 2b), are probably in lag phase (Williamson, 1996; Crooks, 2005); those species could be candidates for successful eradication. The species that could be more worrisome are those expanding faster than expected. However, our data for specific introduced species could be biased by the botanical collection because many botanists have been more interested in collecting native non-weed species than introduced weeds. Thus, instead of focusing in individual species data points, we used the regression model residuals to score the species by estimating their spread rate relative to their residence time.
Priority of attention estimation for introduced leguminous species in Mexico
Priority was estimated by obtaining partial sums for criteria groups (behavior abroad, residence time and distribution, and taxonomic affinity) and then adding these partial sums with the growth habit score (Tables 6, 7, 8). The criteria groups had no significant correlations among them. The total scores sum was obtained for each species (Table 8); then, the species were sorted according to their total to rank each one.
Introduced legumes in Mexico from the MIM database are considered elsewhere in the world (Randall, 2002) as naturalized, 42 (41 species and 1 variety); weed, 41; introduced weed, 26 environmental weed, 21; crop escape, 18; garden escape, 13; invasive casual, 10, and noxious species, 7 (Table 6). The behavior of weeds in other parts of the world has been used as an indicator of the potential behavior of the species in a new area (Rejmánek, 2000); however, the history of the species is not a completely reliable predictor (Rejmánek and Richardson, 1996). Moreover, the behavior of weeds abroad was not correlated with the criterion group of residence time and distribution, indicating that behavior of species abroad might not be a reliable predictor of the introduced Leguminosae behavior in Mexico. Thus, the score sum obtained for the species' behavior abroad, and for the other criterion groups, (Table 8) should be considered more as qualitative indicators more than arithmetic results. We expect that these indicators reflect the ability of the species to invade ecosystems causing economic or environmental damage.
Total scores equal or higher than 6.25 were found for seven species; we suggest that Albizia lebbeck, Pueraria phaseoloides, Lablab purpureus, Securigeravaria, Delonix regia, Clitoria ternatea, and Spartium junceum should receive high-priority attention. These species are distributed in more sites than expected by year of introduction, they have been recorded as weeds and environmental weeds in other countries, and occupy up to nine habitat types. Albizia lebbeck, a widespread ornamental, would require increased vigilance because it has been detected as escaped in almost half the country. Another potentially dangerous species is Puerariaphaseoloides because it is a fodder vine and because its relative P. lobata, is causing serious damage in the countries it has invaded (Starr et al., 1999). P. phaseoloides has no native relatives and its use as fodder facilitates its dissemination. Lablab purpureus, Securigera varia (= Coronilla varia), and Clitoria ternatea are potentially dangerous also being vines used as fodder. The most harmful invasive species in Australia were introduced vines (Phillips et al., 2010), because they can spread rapidly and smother native vegetation. According to Humphries et al. (1991) the exotic vines are one of the plant functional groups most destructive in the ecological context. Delonixregia, a common ornamental tree, has been recorded in nine habitat types in Mexico and, aside of having no Mexican relatives, it is considered a weed or environmental weed elsewhere (Table 6). However, its escape into the wild has not been confirmed. The shrub Spartium junceum, considered elsewhere as environmental and noxious weed, was in the high attention group, although it is reported in fewer localities than expected according to its residence time. However, it does not have close relatives in Mexico, and it might be in the lag phase.
Eight species were considered to require medium priority attention because they have been reported as common environmental weeds or weeds. Some of them are distributed in fewer localities than the expected according to their residence time and it is possible that some are in their lag phase. Feral species, ornamental or fodder crops, predominate in this group: Acacia melanoxylon, Alysicarpus vaginalis, Alysicarpusovalifolius, Crotalaria retusa, Lathyrus latifolius, Lathyrus tingitanus, and Senna didymobotrya. We also classified Bauhinia variegata in this category, despite being known as noxious weed, weed, and relative of environmental weeds because it has 18 non-weed and one weed relatives within the same genus (Torres-Colin, 2006). Thus, it is likely that Bauhinia natural enemies could be attacking B.variegata, preventing it from becoming a problem; Acacia melanoxylon and Crotalaria retusa are probably in the same situation with 11 and 9 native relatives respectively.
We classified seven species as low priority: Cassia javanica, Cercissiliquastrum, Crotalaria pallida, Senna siamea, Sesbania grandiflora, Sutherlandiafrutescens, and Vicia villosa. These species are distributed in fewer localities than expected according to residence time and most have native and introduced relatives. We also included Crotalaria pallida in this category although it is distributed in more sites than expected, but it has 11 native weed relatives in the same genus (Villaseñor and Espinosa-García, 1998).
The remaining 20 taxa (19 species and 1 variety) were classified in a non-priority attention category. These species comprise herbaceous feral crops and agricultural weeds probably introduced in the colonial period. Many of them have high total scores in our model, but none has been mentioned as particularly problematic in the more than 200 years residence in Mexico. Indigofera hirsuta and Tamarindusindica, a shrub and a tree respectively, are not regarded as a problem. We have reserves about Cassia fistula being in this group. It is considered environmental weed, it has been recorded in ten habitat types without native relatives and it has harmful relatives elsewhere. All this suggests that Cassia fistula has great plasticity and no biotic barriers, so it could be in a closing lag phase. This species is also a very showy ornamental tree species cultivated in many places, so propagule pressure is high, thereby facilitating its geographic expansion. This Asian ornamental was first recorded by the Royal Botanical Expedition in the New Spain around 1794 (Blanco Fernández de la Caleya et al., 2010), but it is not clear whether the herbarium specimen was obtained from a feral or cultivated tree. Before we knew about this ancient herbarium record, we classified this species as a high priority of attention species and, if this specimen came from a cultivated tree, Cassia fistula should be considered a high priority species.
Final considerations
The analysis identified species considered noxious or categorized as belonging to the worst weeds in the world, such as Securigera varia, (Williams and Hunyadi, 1987), Albizia lebbeck and Clitoria ternata (Holm et al., 1991), which justified their inclusion in the high priority group. This coincidence suggests that these species, and all the included in the high priority of attention group, should be assessed thoroughly in Mexico, and if necessary, they should be eradicated or contained. It is vital to monitor the dangerous species within and outside their places of origin to take timely management actions (van Kleunen et al., 2010; Ricciardi and Cohen, 2007).
The assessment model presented in this paper still needs further refinement, as seemingly innocuous species scored high and a potentially dangerous species was classified in the non priority category. In both cases the decisive criterion was a very early introduction, thus additional criteria should be developed to prevent false positives or negatives. Still, we hope that the present first-phase introduced assessment model will work as a structured, low expert-dependent approach to identify the introduced species that require a further risk analysis to prioritize efforts for noxious plant management.
ACKNOWLEDGEMENTS
We thank to CONABIO U024 project "Malezas introducidas en México" for the full access to the database. We are very grateful to the staff and curators of the herbaria visited during the development of the MIM database. Heberto Ferreira and Alberto Valencia constantly supported us with computer assistance. Nora Sánchez Blanco helped us capture part of the information. This manuscript benefited greatly with the observations and English corrections from an anonymous reviewer and from copy editing from Rosa María Murillo. This study was supported by a grant from the Centro de Investigaciones en Ecosistemas, Universidad Nacional Autónoma de México, awarded to Francisco J. Espinosa-García (POFJEG).
LITERATURE CITED
Ahern, G. R., A. D. Landis, A. A. Reznicek and W. D. Schemske. 2010. Spread of exotic plants in the landscape: the role of time, growth habit, and history of invasiveness. Biol. Invasions 12: 3157-3169. [ Links ]
Anonymous. 1979. Tropical legumes: Resources for the future. National Academy of Sciences, Library of Congress Catalog Number. 79-64185. [ Links ]
Anonymous. 2010a. Estrategia nacional sobre especies invasoras en México, prevención, control y erradicación. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Comisión Nacional de Áreas Protegidas, Secretaría de Medio Ambiente y Recursos Naturales. México, D.F., México. 110 pp. [ Links ]
Anonymous. 2010b. Herbario CICY, Centro de Investigaciones Científicas de Yucatán. http://www.cicy.mx/ofer-tec-herbario/herbario. Curadoría del Herbario CICY. Consulted 14/02/2012. [ Links ]
Blackburn, T. M., P. Pyšek, S. Bacher, J. T. Carlton, R. P. Duncan, V. Jarosik, J. R. U. Wilson and D. M. Richardson. 2011. A proposed unified framework for biological invasions. Trends Ecol. Evol. 26: 333-339. [ Links ]
Blanco Fernández de la Caleya, P., A. Espejo-Serna and A. R. López Ferrari. 2010. Catálogo del herbario de la Real Expedición Botánica de La Nueva España (1787-1803) conservado en el Real Jardín Botánico de Madrid. Consejo Superior de Investigación Científica del Real Jardín Botánico. Madrid, España. 687 pp. [ Links ]
Bradshaw, C. J. A., X. Giam, H. T. W. Tan, B. W. Brook and N. S. Sodhi. 2008. Threat or invasive status in legumes is related to opposite extremes of the same ecological and life-history attributes. J. Ecol. 96: 869-883. [ Links ]
Calderón, O. and F. J. Espinosa-García. 1997. Manual de identificación de semillas de maleza. Secretaría de Agricultura, Ganadería y Desarrollo Rural. México, D.F., México. 113 pp. [ Links ]
Challenger, A. 1998. Utilización y conservación de los ecosistemas terrestres de México: Pasado, presente y futuro. Comisión Nacional para el Conocimieto y Uso de la Biodiversidad, Instituto de Biología, Universidad Nacional Autónoma de México y Sierra Madre, S.C. México, D.F., México. 847 pp. [ Links ]
Chávez C., A. 2010. Angiospermas. Herbario EBUM de Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo. http://sites.google.com/site/herbarioebum/colecciones/angiospermas. Consulted 14/02/2012. [ Links ]
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press. New York Botanical Garden. New York, U.S.A. pp. xiii-xviii. [ Links ]
Crooks, J. A. 2005. Lag times and exotic species: The ecology and management of biological invasions in slow-motion. Ecoscience 12: 316-329. [ Links ]
Daehler, C. C. 2009. Short lag times for invasive tropical plants: Evidence from experimental plantings in Hawai'i. PloS One 4: 1-5. [ Links ]
Dávila, P. 1992. Un análisis de los herbarios mexicanos. Ciencias 6: 57-61. [ Links ]
Dehnen-Schmutz, K., J. Touza, C. Perrings and M. Williamson. 2007. A century of the ornamental plant trade and its impact on invasion success. Biodivers. Res. 13: 527-534. [ Links ]
Delgadillo R., J. 2008. Flora (vascular) y fauna (aves) de los Parques Nacionales del estado de Baja California: Constitución de 1857 y Sierra San Pedro Mártir. Universidad Autónoma de Baja California. Facultad de Ciencias. Base de datos, del proyecto BK060. Sistema Nacional de Información sobre Biodiversidad-Comisión Nacional para el Conocimieto y Uso de la Biodiversidad. México, D.F., México. 2 pp. http://www.conabio.gob.mx/institucion/proyectos/resultados/BK060_L077_CARCARAT%20BASE%20DE%20DATOS_PLANTAS.pdf. Consulted 14/02/2012. [ Links ]
Elton, C. 1958. The ecology of invasions by animals and plants. Methuen. London, U.K. 186 pp. [ Links ]
Emms, J., J. G. Virtue, C. Preston and W. Bellotti. 2005. Legumes in temperate Australia: A survey of naturalisation and impact in natural ecosystems. Biol. Conserv. 125: 323-333. [ Links ]
Espinosa-García, F. J., J. L. Villaseñor and H. Vibrans. 2004a. The rich generally get richer, but there are exceptions: Correlations between species richness of native plant species and alien weeds and in Mexico. Divers. Distrib. 10: 399-407. [ Links ]
Espinosa-García, F. J., J. L. Villaseñor and H. Vibrans. 2004b. Geographical patterns in native and exotic weeds of Mexico. Weed Technol. 18: 1552-1558. Suppl. S. [ Links ]
Espinosa-García, F. J., J. L. Villaseñor and H. Vibrans. 2009. Biodiversity, distribution, and possible impacts of exotic weeds in Mexico. In: Van Devender, T., F. J. Espinosa-García, B. L. Harper-Lore and T. Hubbard. (eds.). Invasive plants on the move. Controlling them in North America. Arizona-Sonora Desert Museum Press. Tucson, U.S.A. pp. 43-52. [ Links ]
Espinosa-García, F. J., J. Sánchez-Blanco, E. Medina-Murillo and C. Sánchez-Blanco. 2000. Malezas introducidas en México. Universidad Autónoma de México. Centro de Investigaciones en Ecosistemas. Base de datos, del proyecto U024. Sistema Nacional de Información sobre Biodiversidad-Comisión Nacional para el Conocimieto y Uso de la Biodiversidad. México, D.F., México. 6 pp. http://www.conabio.gob.mx/institucion/proyectos/resultados/U024_CARATULA%20BASE%20DE%20DATOS.pdf. Consulted 14/02/2012. [ Links ]
Essl, F. 2007. From ornamental to detrimental? The incipient invasion of Central Europe by Paulownia tomentosa. Preslia. 79: 377-389. [ Links ]
Gasso, N., P. Pyšek, M. Vila and M. Williamson. 2010. Spreading to a limit: the time required for a neophyte to reach its maximum range. Divers. Distrib. 16: 310-311. [ Links ]
Grotkopp, E., E. Erskine-Ogden and M. Rejmánek. 2010. Assessing potential inasiveness of woody horticultural plant species using seedling growth rate traits. J. Appl. Ecol. 47: 1320-1328. [ Links ]
Grotkopp, E., M. Rejmánek and T. L. Rost. 2002. Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species. Am. Nat. 159: 396-419. [ Links ]
Groves, R. H. 2006. Are some weeds sleeping? Some concepts and reasons. Euphytica 48: 111-120. [ Links ]
Groves, R. H., R. Boden and W. M. Lonsdale. 2005. Jumping the garden fence: invasive garden plants in Australia and their environmental and agricultural impacts. CSIRO report prepared for World Wildlife Fund. Australia, Sydney. http://wwf.org.au/publications/jumping_the_garden_fence/. Consulted 14/02/2012. [ Links ]
Heger, T. and L. Trepl. 2003. Predicting biological invasions. Biol. Invasions 5: 313-321. [ Links ]
Holm, L., D. Plucknett, J. Pancho and J. Herberger. 1991. The world's worst weeds: distribution and biology. University Hawaii Press. Honolulu; Krieger Publ. Co., Malabar, Florida, U.S.A. 610 pp. [ Links ]
Huang, Q. Q., J. M. Wu, Y. Y. Bai, L. Zhou and G. X. Wang. 2009. Identifying the most noxious invasive plants in China: role of geographical origin, life form and means of introduction. Biodivers. Conserv. 18: 305-316. [ Links ]
Humphries, S. E., R. H. Groves and D. S. Mitchell. 1991. Plant invasions of Australian ecosystems: a status review and management directions. In: Longmore, R. (ed.). Kowari 2. Plant invasions: The incidence of environmental weeds in Australia. Australian National Parks and Wildlife Service. Canberra, Australia. pp. 1-134. [ Links ]
Koop, A. L., L. Fowler, L. P. Newton and B. P. Caton. 2012. Development and validation of a weed screening tool for the United States. Biol. Invasions 14: 273-294. [ Links ]
Kowarick, I. 1995. Time lags in biological invasions with regard to the success and failure of alien species. In: Pyšek, P. K., M. Prach, M. Rejmánek and P. M. Wade (eds.). Plant invasions: General aspects and special problems. S. P. B. Academic Publishing. Amsterdam, The Netherlands. pp. 15-38. [ Links ]
Lockwood, J. L., D. Simberloff, M. L. McKinney and B. Von Holle. 2001. How many, and which, plants will invade natural areas? Biol. Invasions 1: 1-8. [ Links ]
Lonsdale, W. N. 1999. Global patterns of plant invasions and the concept of invasibility. Ecology 80(5): 1522-1536. [ Links ]
Mack, R. N. 1981. Invasion of Bromus tectorum L. into western North America: an ecological chronicle. Agro-Ecosystems 7: 145-165. [ Links ]
Mack, R. N. 1991. The commercial seed trade: an early disperser of weeds in the United States. Econ. Bot. 45: 257-273. [ Links ]
Mack, R. N. 1996. Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biol. Conserv. 78: 107-121. [ Links ]
Mack, R. N., D. Simberloff, W. M. Londsdale, H. Evans, M. Clout and F. A. Bazzaz. 2000. Biotic invasions: causes, consequences and control. Ecology 10: 689-710. [ Links ]
March Mifsut, I. J. and M. Martínez Jiménez. (eds.). 2007. Especies invasoras de alto impacto a la biodiversidad. Prioridades en México. Instituto Mexicano de Tecnología del Agua, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Grupo de Ecología y Conservación de Islas, A. C. AridAmérica, The Nature Conservancy. Jiutepec, México. 72 pp. [ Links ]
Meyer, J. Y. 2000. Preliminary review of the invasive plants in the Pacific Islands (SPREP Member countries). In: Sherley, G. (ed.). Invasive species in the Pacific: A technical review and draft regional strategy. South Pacific Regional Environmental Programme. Samoa, U.S.A. pp. 85-114. [ Links ]
Panetta, F. D. and N. D. Mitchell. 1991. Homoclimate analysis and the prediction of weediness. Weed Res. 31: 273-284. [ Links ]
Pheloung, P. C., P. A. Williams and S. R. Halloy. 1999. A weed risk assessment model for use as a biosecurity tool evaluating plant introductions. J. Environ. Manage. 57: 239-251. [ Links ]
Phillips, M. L., B. R. Murray, M. R. Leishman and R. Ingram. 2010. The naturalization to invasion transition: Are there introduction-history correlates of invasiveness in exotic plants of Australia? Austral. Ecol. 35: 695-703. [ Links ]
Pyšek, P. 1998. Is there a taxonomic pattern to plant invasions? Oikos 82: 282-294. [ Links ]
Randall, R. 2002. A global compendium of weeds. R. G. and F. G. Richardson. Meredith, Australia. 944 pp. [ Links ]
Randall, J. M., N. Benton and L. E. Morse. 2001. Categorizing invasive weeds: the challenge of rating the weeds already in California. In: Groves, R. H., F. D. Panetta and J. G. Virtue (eds.). Weed risk assessment. CSIRO Publishing. Collingwood, Australia. pp. 203-216. [ Links ]
Randall, J. M., L. E. Morse, N. Benton, R. Hiebert, S. Lu and Killeffer, T. 2008. The invasive species assessment protocol: a tool for creating regional and national lists of invasive nonnative plants that negatively impact biodiversity. Invasive Plant Sci. Manag. 1: 36-49. [ Links ]
Rejmánek, M. 2000. Invasive plants: approaches and predictions. Austral Ecol. 25: 497-506. [ Links ]
Rejmánek, M. and D. M. Richardson. 1996. What attributes make some plant species more invasive? Ecology 77: 1655-1661. [ Links ]
Rejmánek, M. and M. J. Pitcairn. 2002. When is eradication of exotic pest plants a realistic goal? In: Veitch, C. R. and M. N. Clout (eds.). Turning the tide: the eradication of invasive species. International Union for Conservation of Nature. Species Survival Commission. Invasive Species Specialist Group. International Union for Conservation of Nature. Cambridge, U.K. pp. 249-253. [ Links ]
Ricciardi, A. and J. Cohen. 2007. The invasiveness of an introduced species does not predict its impact. Biol. Invasions 9: 309-315. [ Links ]
Richardson, M. D., P. Pyšek, M. Rejmánek, M. G. Barbour, F. D. Panetta and C. J. West. 2000. Naturalization and invasion of alien plants: concepts and definitions. Divers. Distrib. 6: 93-107. [ Links ]
Rzedowski, J., G. Calderón de Rzedowski and A. Butanda Cervera. 2009. Los principales colectores de plantas activos en México entre 1700 y 1930. Instituto de Ecología, A.C., Centro Regional del Bajío. Pátzcuaro, México. 133 pp. [ Links ]
Sánchez-Blanco, C. 2003. Análisis de la distribución de malezas introducidas a México de las familias Cesalpinaceae, Fabaceae y Mimosaceae. Tesis de Biólogo. Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo. Morelia, Michoacán, México. 121 pp. [ Links ]
Serrano Cárdenas, V. 2010. Base de datos del Herbario de Querétaro "Dr. Jerzy Rzedowski" (QMEX). Base de datos, del proyecto EC011. Sistema Nacional de Información sobre Biodiversidad-Comisión Nacional para el Conocimieto y Uso de la Biodiversidad. Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro. México, D.F., México. 1 pp. http://www.conabio.gob.mx/institucion/proyectos/resultados/EC011%20CARATULA%20BASE%20DE%20DATOS.pdf. Consulted 14/02/2012. [ Links ]
Sousa, S. M. and A. Delgado. 1993. Mexican Leguminosae: Phytogeography, endemism and origins. In: Ramamoorthy, T. P., R. Bye, A. Lot and J. Fa (eds.). Biological diversity of Mexico: Origins and distribution Cap. 17. Oxford University Press. New York, U.S.A. pp. 459-511. [ Links ]
Starr, F., K. Martz and L. Loope. 1999. Kudzu (Pueraria lobata). An alien plant report. http://www.hear.org/species/reports/puelob_fskm_awwa_report.pdf. Consulted 14/02/2012. [ Links ]
Strauss, S. Y., C. O. Webb and N. Salamin. 2006. Exotic taxa less related to native species are more invasive. Proc. Natl. Acad. Sci. Environ. Sci. 15: 5841-5845. [ Links ]
Torres-Colín, R. 2006. Two new species of Bauhinia (Fabaceae, Caesalpinioideae, Cercideae) from Southeastern Mexico and Mesoamerica. Novon 16: 533-537. [ Links ]
van Kleunen, M., W. Dawson, D. Schlaepfer, J. M. Jeschke and M. Fischer. 2010. Are invaders different? A conceptual framework of comparative approaches for assessing determinants of invasiveness. Ecol Lett. 13: 947-958. [ Links ]
Vega Aviña, R. 2000. Catálogo y base de datos preliminar de la flora de Sinaloa. Facultad de Agronomía, Universidad Autónoma de Sinaloa. Base de datos, del proyecto L057. Sistema Nacional de Información sobre Biodiversidad-Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, D.F., México. 2 pp. http://www.conabio.gob.mx/institucion/proyectos/resultados/Conabio_Contenido%20L057.pdf. Consulted 14/02/2012. [ Links ]
Vibrans, H. 2009. Lathyrus odoratus L. Ficha Informativa. http://www.conabio.gob.mx/malezasdemexico/fabaceae/lathyrus-odoratus/fichas/ficha.htm. Consulted May 9, 2012. [ Links ]
Vilá, M., J. L. Espinar, M. Hejda, P. E. Hulme, V. Jarosik, J. L. Maron, J. Pergl, U. Schaffner, Y. Sun and P. Pyšek. 2011. Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 14: 702-708. [ Links ]
Villaseñor R., J. L. and F. J. Espinosa-García. 1998. Catálogo de malezas de México. Universidad Nacional Autónoma de México, Consejo Nacional Consultivo Fitosanitario, Fondo de Cultura Económica. México, D.F., México. 448 pp. [ Links ]
Villaseñor R., J. L. and F. J. Espinosa-García. 2004. The alien flowering plants of México. Divers. Distrib. 10: 113-123. [ Links ]
Vitousek, P. M., C. M. D'Antonio, L. Loope and R. R. Westbrooks. 1996. Biological invasions as global environmental change. Am. Sci. 84: 468-478. [ Links ]
von der Lippe, M. and I. Kowarik. 2008. Do cities export biodiversity? Traffic as dispersal vector acroos urban-rural gradients. Divers. Distrib. 14: 18-25. [ Links ]
Williams, G. and K. Hunyadi. 1987. Dictionary of weeds of Eastern Europe. Elsevier. Amsterdam, The Netherlands. 480 pp. [ Links ]
Williams, J.K. 2010. Additions to the alien vascular flora of Mexico, with comments on the shared species of Texas, Mexico, and Belize. Phytoneuron 3: 1-7. [ Links ]
Williamson, M. 1996. Biological invasions. Chapman & Hall, London, U.K. 244 pp. [ Links ]
Williamson, M. and A. Fitter. 1996. The varying success of invaders. Ecology 77: 1661-1666. [ Links ]
Williamson, M., K. Dehnen-Schmutz, I. Kuhn, M. Hill, S. Klotz, A. Milbau, J. Stout and P. Pyšek. 2009. The distribution of range sizes of native and alien plants in four European countries and the effects of residence time. Divers. Distrib. 15: 158-166. [ Links ]
Wilson, J. R. U., D. M. Richardson, M. Rouget, Ş. Procheş, M. A. Amis, L. Henderson and W. Thuiller. 2007. Residence time and potential range: crucial considerations in modelling plant invasions. Divers. Distrib. 13: 11-22. [ Links ]
Wu, S. H., S. M. Chaw and M. Rejmánek. 2003. Naturalized Fabaceae (Leguminosae) species in Taiwan: the first approximation. Bot. Bull. Acad. Sinica 44: 59-66. [ Links ]
NOTA
* Part of this work comes from the Bachelor's thesis of C. Sánchez-Blanco that was directed by F.J. Espinosa-García.

