










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkActa botánica mexicana
versión On-line ISSN 2448-7589versión impresa ISSN 0187-7151
Act. Bot. Mex no.92 Pátzcuaro jul. 2010
Variación morfológica del género Prochnyanthes (Agavaceae)
Morphological variation of the genus Prochnyanthes (Agavaceae)
Arturo Castro–Castro, Aarón Rodríguez, Georgina Vargas–Amado y Raymundo Ramírez–Delgadillo
Universidad de Guadalajara, Departamento de Botánica y Zoología, Apdo. postal 139, 45101 Zapopan, Jalisco, México. arca68@hotmail.com
Recibido en junio de 2008
Aceptado en mayo de 2010
RESUMEN
Se analizó la variación morfológica del género Prochnyanthes, mediante el examen de 29 caracteres, en una muestra de 544 individuos de 19 localidades. Se aplicó el Análisis de Componentes Principales, el cual muestra los patrones de variación, analiza las similitudes de las unidades de clasificación e identifica grupos. Con base en la evidencia morfológica, se amplió la descripción de P. mexicana y se logró reconocer dos fenotipos, considerándolos como extremos de su variación. El tamaño y el color del perianto así como la longitud de los pedicelos no mostraron la suficiente variación para diferenciar grupos y en consecuencia se reconoció a P. mexicana como la única especie del género.
Palabras clave: Agavaceae, análisis de componentes principales, morfología, Prochnyanthes.
ABSTRACT
The morphologic variation of the genus Prochnyanthes was analyzed by means of the examination of 29 characters, in a sample of 544 individuals from 19 localities. The Principal Components Analysis was used, which shows the variation trends, analyses the similarities among the classification units and identifes groups. Based on the morphological evidence, it was possible to recognize two phenotypes, considered as extremes of a morphological cline. Therefore, the morphological species concept of P. mexicana was broaden to include all the variation observed. Finally, the size and color of the perianth and the length of the pedicels did not show enough variation to identify groups and consequently P. mexicana is the only recognized species of the genus.
Key words: Agavaceae, morphology, principal components analysis, Prochnyanthes.
INTRODUCCIÓN
El género Prochnyanthes S. Watson es endémico de México. Sus componentes crecen a lo largo de la Sierra Madre Occidental y el Eje Volcánico Transversal en los estados de Aguascalientes, Durango, Guanajuato, Jalisco, Michoacán, Nayarit, Querétaro y Zacatecas (García–Mendoza y Galván, 1995). Habitan en laderas rocosas con vegetación de bosque de encino, pino y encino, pastizal y zonas de transición entre el encinar y el bosque tropical caducifolio, en altitudes de 1000 a 2700 m. Los nombres comunes registrados para Prochnyanthes son amole, amolilla, huaco y lirio. El macerado del cormo y de las raíces se utiliza como sustituto del jabón e insecticida (Verhoek, 1978).
Las relaciones filogenéticas de Prochnyanhtes han sido analizadas pero no existe hasta ahora un acuerdo al respecto. Bogler y Simpson (1996) mostraron la estrecha relación entre los géneros Manfreda Salisb., Polianthes L. y Prochnyanthes, considerándolos como un clado independiente de Agave L. Más aún, Thiede y Eggli (1999), con base en datos moleculares y morfológicos (Bogler et al., 1995; Bogler y Simpson, 1995, 1996; Clary y Simpson, 1995; Hernández, 1995), incluyeron a los géneros Manfreda, Polianthes y Prochnyanthes dentro de Agave, reconociendo el epíteto Agave bulliana (Baker) Thiede & Eggli, bajo el cual se incluye a Prochnyanthes mexicana.
Los límites interespecíficos dentro del género son poco claros. Algunos autores consideran que se trata de una sola especie morfológicamente muy variable (Verhoek–Williams, 1975; McVaugh, 1989; Espejo–Serna y López–Ferrari, 1993; Williams, 1998). Otros reconocen más y han descrito a las especies: P. viridescens S. Watson (1887), P. bulliana Baker (1895) y P. mexicana (Zucc.) Rose (1903) como taxa separados. Los caracteres utilizados para su delimitación son la longitud del pedicelo y el tamaño del perianto; sin embargo, estos rasgos son los que muestran mayor variación entre individuos de una población e incluso en un mismo individuo pero no distinguen a poblaciones.
Prochnyanthes bulliana fue diferenciada de P. viridescens con base en sus pedicelos cortos y flores púrpura–verdosas. Una segunda descripción indica que P. bulliana presenta flores sésiles con la parte externa del perianto de color verde–pardo y el interior amarillento (Baker 1884, 1895). Prochnynahtes mexicana fue definida como planta con flores sésiles de perianto más largo y delgado que el de P. bulliana. De acuerdo con el protólogo de P. viridescens, ésta presenta flores amarillo–verdosas con tintes pardos y pedicelos ascendentes de 2.5–4 cm de longitud (Verhoek–Williams 1975; Cuadro 1).

El propósito del presente trabajo fue estudiar estadísticamente la variación morfológica en Prochnyanthes, mediante la aplicación del Análisis de Componentes Principales (ACP) que permite analizar la matriz de correlaciones y transformar un grupo de variables relacionadas en un conjunto menor de variables independientes entre sí llamadas componentes principales. De esta manera, se consideran a la vez varias características, resolviendo las dimensiones latentes en los datos (Lamboy, 1990; Torres, 2000), y se analizan los patrones de variación.
MÉTODOS
Se estudiaron 19 poblaciones localizadas en los estados de Aguascalientes, Durango, Guanajuato, Jalisco, Michoacán, Nayarit, Querétaro y Zacatecas (Apéndice 1). El criterio para la selección de los sitios de colecta se basó en ejemplares de herbario que durante la revisión mostraron características interesantes para el análisis y que cubrieron la mayor parte del área de distribución. También se consideraron las sugerencias de otros botánicos, lo reportado en la literatura botánica y la experiencia personal producto de colectas previas. Asimismo, se puso interés especial en las localidades tipo (Zuccarini, 1837; Baker, 1884, 1895; Watson, 1887; Rose, 1903; McVaugh, 1989).
La consulta de herbarios y recolección de material botánico se realizó de agosto a octubre de 2005. Se revisaron 95 ejemplares botánicos de los siguientes herbarios: Herbario de la Universidad de Guadalajara (IBUG), Herbario del Instituto de Ecología A.C., Centro Regional del Bajío (IEB), Herbario del Centro Interdisciplinario para el Desarrollo Integral Regional–Durango (CIIDIR), Herbario del Instituto de Biología de la Universidad Nacional Autónoma de México (MEXU) y el Herbario de la Universidad Autónoma de Querétaro (QMEX). Estos ejemplares no se analizaron porque no se apreciaban en ellos todas las características estudiadas (Apéndice 2).
Se examinaron 30 individuos vivos en estado reproductivo por población. La única excepción fue la del estado de Querétaro, en el cual sólo fue posible estudiar cuatro plantas (Apéndice 1). Se evaluaron 29 caracteres (Fig. 1; Cuadro 2). Las características florales se obtuvieron del par de flores con el estilo maduro al momento de la medición. El tamaño de la muestra consistió de 544 ejemplares. La matriz básica de datos está disponible a través del primer autor. Se colectaron cinco individuos por población y se distribuyeron a los siguientes herbarios: ENCB, IBUG, IEB, MEXU y XAL.
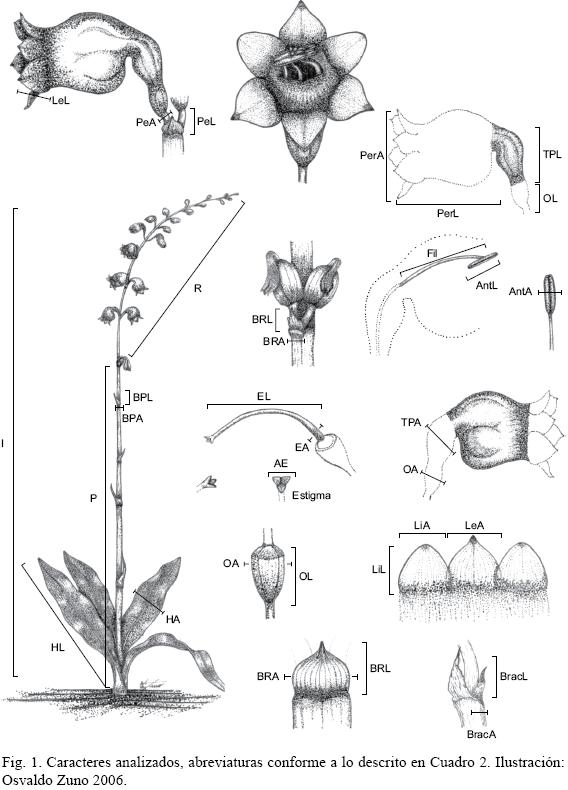

El ACP se ejecutó a través del programa de cómputo SPSS 9.0 (Anónimo, 1999) y se realizaron dos análisis. En el primero se consideraron las 29 variables y se encontró que ocho de ellas: longitud de la bráctea del pedúnculo, longitud de la antera, ancho de la antera, longitud del estilo, ancho del estilo, ancho de la bráctea del pedúnculo, ancho de la bráctea del raquis y ancho del ovario, presentaron coeficientes de correlación bajos y aportaron poca variación al modelo. En consecuencia, se eliminaron de subsecuentes análisis. Asimismo, la longitud del pedúnculo y la longitud del raquis fueron eliminadas por ser redundantes, debido a que se consideraron variables correlacionadas lógicamente con la longitud de la inflorescencia, además de mostrar bajo aporte de variación.
Con base en los resultados del primer ACP, se elaboraron variables compuestas para la ejecución del segundo análisis. Estas fueron las siguientes proporciones: 1) largo/ancho del perianto (forma del perianto), 2) largo/ancho de las hojas (forma de las hojas), 3) largo/ancho de las anteras (forma de las anteras), y 4) largo/ancho del pedicelo (forma del pedicelo). La forma de la hoja y pedicelo, así como la longitud del perianto y filamento aportaron poca variación al modelo, sin embargo se decidió conservarlos para el análisis debido a su supuesto valor en el reconocimiento de las especies de Prochnyanthes (Cuadro 1).
RESULTADOS
El análisis mostró siete componentes principales que contienen 77.9% de la variación total (Cuadro 3). La extracción se realizó con base en el criterio de raíz latente (Crisci y López–Armengol, 1983; Hair et al., 1999). De las 23 variables estudiadas, 20 están contenidas en los tres primeros componentes: 15 en el principal, tres en el segundo y dos en el tercero (Cuadro 4).
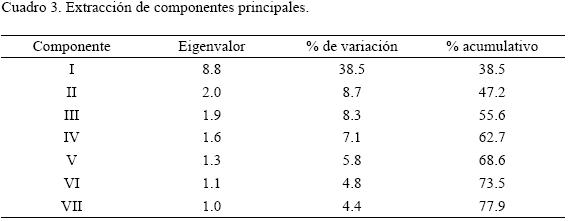

Generalmente y con fines prácticos se utilizan e interpretan los tres primeros componentes que contienen la mayor variación, lo que reduce el número de dimensiones (Anónimo, 1999). En el presente estudio, la variación contenida en los tres primeros es de 55.6%.
El componente principal I (CP I) acumuló 38.5% de la variación. Éste tiene relación con la talla de la planta y la forma del perianto; aquellos individuos que tienen mayores dimensiones, tienden a presentar flores campanuladas (Fig. 2. C–L). En contraste, los de menor tamaño, propenden a producir flores tubulares (Fig. 2. A, B). Los caracterizados por tallas menores y flores tubulares (CP I menores a –1, CP II mayores a 0, Fig. 3) forman parte de las dos poblaciones del estado de Aguascalientes y de la correspondiente al Puerto la Campana, municipio de Atenguillo, Jalisco. En el otro extremo, las poblaciones con individuos grandes y de flores campanuladas son características del resto de la muestra y representan 77.9%. Este último fenotipo concuerda con las descripciones realizadas por Baker (1884 y 1895), Watson (1887) y Rose (1903) y coincide con lo examinado por Verhoek–Williams (1975) y Williams (1998). La talla de la planta y la forma del perianto permiten el reconocimiento de dos morfos; sin embargo, sólo se pueden considerar extremos de variación de una sola especie, ya que de acuerdo con la gráfica de dispersión entre el CP I y el CP II, no cabe reconocer grupos independientes (Fig. 3).

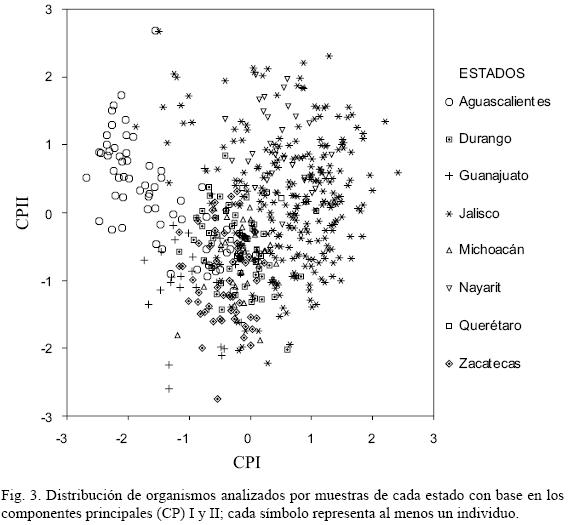
En general, existe una alta relación morfológica entre los individuos analizados. No obstante, se observan extremos de la variación que corresponden a las dos localidades del estado de Aguascalientes. Los ejemplares de San José de Gracia (AGSGracia) muestran caracteres intermedios entre los de Sierra Fría (AGSsp) y todos los demás, siendo este último sitio el que señaló mayor diferencia fenotípica (Fig. 2; Apéndice 1).
Las plantas estudiadas de Jalisco señalaron un amplio intervalo de variación morfológica y distribución espacial, relacionándose con el resto de los sujetos analizados (Fig. 3, 4). Se observa una gran similitud de los organismos entre localidades, con una agrupación centralizada en torno a la localidad tipo de Prochnyanthes viridescens (JALR.Bco) y extremos de variación que corresponden a las poblaciones del Puerto la Campana (JALCampa), Ameca (JALAmec) y Tapalpa (JALTapalp; Apéndice 1).

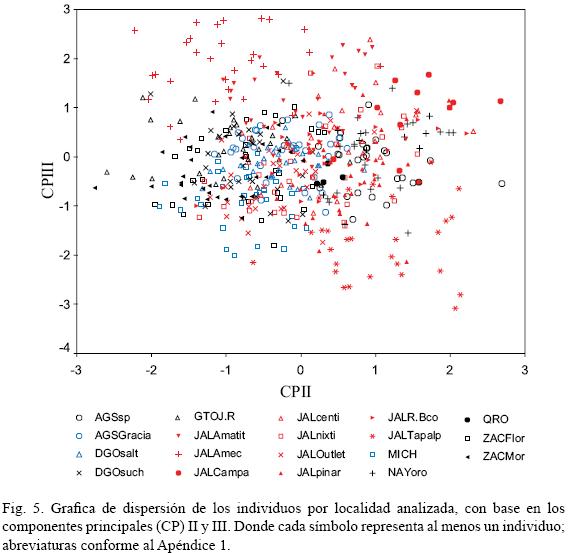
El componente principal II (CP II) contiene 8.7% de la variación. Se relaciona negativamente con el ancho de la hoja y positivamente con la longitud del filamento y la forma de la misma. Es decir, a medida que aumenta el ancho de la hoja, ésta tiende a ser elíptica y se presentan flores con filamentos cortos. Sin embargo, tal variación es clinal, de modo que la forma foliar y el largo del filamento no permiten el reconocimiento de ningún fenotipo.
El componente principal III (CP III) acumula 8.3% de la variación. Se relaciona positivamente con la forma del pedicelo y con la longitud de la bractéola. Por tradición, la longitud del pedicelo ha sido utilizada como un carácter útil en la delimitación de las especies (Baker, 1895; Watson, 1887; Rose, 1903). Sin embargo, en nuestro análisis la forma del pedicelo y el largo de la bractéola no permiten el reconocimiento de fenotipos. La figura 5 muestra la distribución espacial entre el CP II y el CP III. En su extremo superior se observan agrupados individuos con pedicelos largos (1.375–8.94 cm), los cuales corresponden a las localidades de Ameca y Amatitán en el estado de Jalisco, que se ajustan a la descripción de Prochnyanthes viridescens (Watson, 1887) y que proceden de sitios cercanos a la localidad tipo (Río Blanco, Zapopan, Jalisco). Por su parte, los de Tapalpa, Jalisco, con pedicelos cortos (0.329–2.097 cm, Fig. 2 I, L), se asemejan mejor a lo referido por Baker (1895) y Rose (1903) (Cuadro 1).
La figura 6 exhibe las tendencias fenotípicas de los individuos en relación con su ubicación en los diferentes estados de la República. Se observa afinidad entre los organismos de los estados de Jalisco y Nayarit (tendencia A), que se caracterizan por presentar hojas elípticas y filamentos largos, respecto a aquellos de Zacatecas, Durango, Guanajuato y Michoacán (tendencia B), con características antagónicas. Por último, las poblaciones del estado de Aguascalientes (tendencia C, Fig. 2) se diferencian por tener tallas menores y flores tubulares.

Estas observaciones contrastan con lo conseguido por otros autores en diferentes grupos de plantas, que utilizando la misma técnica, muestran en sus resultados grupos de individuos bien segregados, los cuales son considerados como taxa diferentes (Allen, 1984; Franceschinelli et al., 1999; Henderson, 2005; Theriot y Stoermer, 1984 y Vargas, 2006).
Cuarenta por ciento de la variación morfológica analizada en Prochnyanthes permite separar plantas pequeñas de flores tubulares de plantas grandes con flores campanuladas. Sin embargo, estos fenotipos representan dos extremos de la variabilidad contenida en el género. Los patrones analizados definen una variación continua en las características morfológicas, que impiden el reconocimiento de las especies propuestas por otros autores. Asimismo, la longitud de los pedicelos no es un carácter útil, pues muestra un amplio intervalo de variación dentro de una población e incluso dentro de un individuo, pero no entre poblaciones. En consecuencia, P. mexicana es la única entidad reconocida del género, ameritando una ampliación de la descripción morfológica. Finalmente, se plantea la hipótesis de que las diferencias morfológicas observadas en las poblaciones analizadas sugieren la existencia de un proceso de especiación dentro del grupo.
Prochnyanthes mexicana (Zucc.) Rose, Contr. U.S. Nat. Herb. 8: 14. 1903. Polianthes mexicana Zucc. Abh. Math.–Phys. Cl. Königl. Bayer. Akad. Wiss. 2: 319. Prochnyanthes viridescens S. Watson, Proc. Amer. Acad. 22: 457. 1887. Prochnyanthes bulliana (Baker) Baker, Bot. Mag. 121: t. 7427. 1895. Bravoa bulliana Baker, Gard. Chron. ser. 2, 22: 328. 1884.
Hierba erecta, perenne, que emerge de un cormo vertical el cual produce raíces fusiformes carnosas; hojas (1)2–3(7), las más largas en una roseta basal, a medida que las hojas se acercan al escapo floral se hacen más pequeñas y delgadas y se confunden con las brácteas, láminas lineares a elípticas, de 8–65 cm de longitud y de 1–8 cm de ancho, blandas o coriáceas, verdes, ápice rematando en una espina blanda, subpecioladas, con numerosas papilas ásperas en las nervaduras y una fuerte nervadura central, bases de las hojas dilatadas, de color café, fibrosas, imbricadas, formando un bulbo delgado, fibroso y ovoide, de 8–13 cm de longitud; inflorescencia pseudoracemosa, escapiforme, de 0.6–2.8 m de altura, con 2–25 nudos florales, raquis de 20–30(–80) cm de longitud; flores dos por nudo, pedicelos de (0.02)2.5–4(–9) cm de longitud y 0.07–0.35 cm de diámetro, brácteas lanceoladas, de 0.5–6(–10) cm de longitud; bractéola filiforme, de 0.1–1(–1.3) cm de longitud; perianto tubular, de 1.1–2.9 cm de longitud, de simetría bilateral, lóbulos 6, erectos, superpuestos, deltoides, de 0.3–1.5 cm de longitud y 0.3–1.2 cm de ancho, pubescentes en el ápice, mucho más cortos que el tubo, tubo erecto o ascendente, de 0.5–1.75 cm de longitud y de 0.3–0.8 cm de diámetro, que declina y se expande en la mitad, parte distal campanulada a tubular, de 1.1–2.5 cm de largo y 0.5–2.0 cm de ancho; estambres 6, incluidos, filamentos insertos cerca de la base de la porción expandida del tubo del perianto, de 0.8–2.0 cm de longitud, anteras lineares, de 0.3–1.5 cm de longitud y 0.1–0.7 cm de ancho, dorsifijas; estilo filiforme, incluido pero rebasando a los estambres, de 1.2–3.2 cm de largo y 0.6–2.2 mm de diámetro, estigma dilatado distalmente con tres lóbulos apicales, planos, recurvados y ovados, de 0.7–4.0 mm de largo; ovario ínfero, trilocular, óvulos axilares y biseriados; fruto capsular, globoso a oblongo, de 0.8–2.5 cm de largo y 1.0–1.7 cm de diámetro, trilocular, cápsula coronada por el perianto persistente; semillas planas, deltoides o semicirculares, de 0.4–0.6 cm de diámetro, negras.
AGRADECIMIENTOS
Agradecemos a los curadores de los herbarios CIIDIR, IBUG, IEB, MEXU y QMEX las facilidades concedidas. Asimismo, gracias por las revisiones y sugerencias al manuscrito hechas por Jorge A. Pérez de la Rosa, Mollie Harker, Martha Cedano y a los revisores anónimos. A Luis Hernández se agradece la ayuda en la localización de los individuos de Querétaro. A Osvaldo Zuno agradecemos la elaboración de las ilustraciones. Por último, reconocemos la valiosa ayuda de todas las personas involucradas en el trabajo de campo.
LITERATURA CITADA
Allen, G. A. 1984. Morphological and cytological variation in the Western North America Aster occidentalis complex (Asteraceae). Syst. Bot. 9: 175–191. [ Links ]
Anónimo. 1999. Statistical Package for the Social Sciences (SPSS), Base 9.0. Applications Guide. Chicago, EUA. [ Links ]
Baker, J. G. 1884. Bravoa bulliana Baker n. sp. Gard. Chron. 22: 328. [ Links ]
Baker, J. G. 1895. Prochnyanthes bulliana. Bot. Mag. 121: t. 7427. [ Links ]
Bogler, D. J., J. L. Neff y B. B. Simpson. 1995. Multiple origins of the yucca–yucca moth association. Proc. Nat. Acad. Sci. USA 92: 6864–6867. [ Links ]
Bogler, D. J. y B. B. Simpson. 1995. A chloroplast DNA study of the Agavaceae. Syst. Bot. 20: 191–205. [ Links ]
Bogler, D. J. y B. B. Simpson. 1996. Phylogeny of Agavaceae based on its rDNA sequence variation. Am. J. Bot. 83: 1225–1235. [ Links ]
Clary, K. H. y B. B. Simpson. 1995. Systematics and character evolution of the genus Yucca L. (Agavaceae): evidence from morphology and molecular analyses. Bol. Soc. Bot. Mex. 56: 77–88. [ Links ]
Crisci, J. V. y M. F. López–Armengol. 1983. Introducción a la teoría y práctica de la taxonomía numérica. Departamento de Asuntos Científicos y Tecnológicos, Secretaría General de la Organización de los Estados Americanos. Washington, D. C. pp. 71–82. [ Links ]
Espejo–Serna, A. y A. R. López–Ferrari. 1993. Las monocotiledoneas mexicanas: una sinopsis forística. Lista de referencia I: Agavaceae, Alismataceae, Alliaceae, Alstroemeriaceae y Amaryllidaceae. Consejo Nacional de la Flora de México, Universidad Autónoma Metropolitana–Iztapalapa, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, D.F. 78 pp. [ Links ]
Franceschinelli, E. V., K. Yakamoto y G. J. Shepherd. 1999. Distinctions among three Simarouba species. Syst. Bot. 23: 479–488. [ Links ]
García–Mendoza, A. y R. Galván. 1995. Riqueza de las familias Agavaceae y Nolinaceae en México. Bol. Soc. Bot. Mex. 56: 7–24. [ Links ]
Hair, J. F. Jr., R. E. Anderson, R. L. Tathan y W. C. Black. 1999. Análisis multivariante. Prentice Hall. Quinta edición. Madrid. 832 pp. [ Links ]
Henderson, A. 2005. A multivariate study of Calyptrogyne (Palmae). Syst. Bot. 30: 60–83. [ Links ]
Hernández, L. 1995. Análisis cladístico de la familia Agavaceae. Bol. Soc. Bot. Mex. 56: 57–68. [ Links ]
Lamboy, W. F. 1990. Disjoint Principal Component Analysis: a statistical method of botanical identification. Syst. Bot. 51: 3–12. [ Links ]
McVaugh, R. 1989. Bromeliaceae to Dioscoreaceae. Flora Novo–Galiciana: a descriptive account of the vascular plants of Western Mexico Vol. 15. University at Michigan Herbarium. Ann Arbor, Michigan. 398 pp. [ Links ]
Rose, J. N. 1903. Prochnyanthes and its species. Contr. U. S. Natl. Herb. 8(1): 13–14. [ Links ]
Theriot, E. y E. F. Stoermer. 1984. Principal component analysis of variation in Stephanodiscus rotula y S. niagarae (Bacillariophyceae). Syst. Bot. 9: 53–59. [ Links ]
Thiede, J. y U. Eggli. 1999. Einbeziehung von Manfreda Salisbury, Polianthes Linné und Prochnyanthes S. Watson in Agave Linné (Agavaceae). Kakt. and Sukk. 50: 109–113. [ Links ]
Torres M., J. P. 2000. La utilización de técnicas estadísticas multivariables. Estudio de caso. Tesis de maestría. Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Guadalajara, Jalisco. 102 pp. [ Links ]
Vargas, A. G. 2006. Variación morfológica foliar de Dioon (Zamiaceae) en la vertiente del Pacífico. Tesis de maestría. Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Guadalajara, Jalisco. 202 pp. [ Links ]
Verhoek–Williams, S. E. 1975. A study of the tribe Poliantheae (including Manfreda) and revision of Manfreda and Prochnyanthes (Agavaceae). PhD thesis, Cornell University. Ithaca, New York. 405 pp. [ Links ]
Verhoek, S. 1978. Huaco and amole: a survey of the uses of Manfreda and Prochnyanthes. Econ. Bot. 32: 124–130. [ Links ]
Watson, S. 1887. List of plants collected by Dr. Edward Palmer in the state of Jalisco, Mexico in 1886. Proc. Amer. Acad. Arts 22: 396–465. [ Links ]
Williams, S. 1998. Agavaceae. In. Kubitzki, K. (ed.). The families and genera of vascular plants III: flowering plants. Monocotyledons, Lilianae (except Orchidaceae). Springer. Berlin. pp. 60–70. [ Links ]
Zuccarini, J. G. 1837. Plantarum novarum vel minus cognitarum. Abh. Math.–Phys. Cl. Königl. Bayer. Akad. Wiss. 2: 309–380. [ Links ]

