










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkActa botánica mexicana
versão On-line ISSN 2448-7589versão impressa ISSN 0187-7151
Act. Bot. Mex no.90 Pátzcuaro Jan. 2010
Diversidad de frutos de los árboles del bosque tropical perennifolio de México*
Tree fruit traits diversity in the tropical rain forest of Mexico
Guillermo Ibarra–Manríquez1 y Guadalupe Cornejo–Tenorio1
1 Universidad Nacional Autónoma de México, Centro de Investigaciones en Ecosistemas, Antigua carretera a Pátzcuaro Núm. 8701, Col. San José de La Huerta, 58190 Morelia, Michoacán, México. gibarra@oikos.unam.mx
Recibido en septiembre de 2008
Aceptado en septiembre de 2009
RESUMEN
Se describe la morfología del fruto (color, tipo y tamaño) de 881 especies arbóreas del bosque tropical perennifolio de México, incluidas en 338 géneros y 85 familias. De un total de 46 tipos de frutos, tres abarcan 52.7% de las especies (212, 163 y 89 con bayas, drupas y cápsulas loculicidas, respectivamente) y 10 se registraron sólo en una. Los colores de fruto más frecuentes (n = 865 especies) fueron el negro (307 especies; 35.5%), pardo (227; 26.2%) y rojo (159; 18.4%); las bayas fueron usualmente negras (66%), las drupas negras (53.9%) o rojas (39.3%) y las cápsulas loculicidas en su mayoría pardas (66.3%). El largo de los frutos del total de especies osciló entre 0.8 y 430 mm (promedio 31.7 mm) y el ancho de 1 a 145 mm (promedio 16.1 mm), con una correlación significativa entre sí (r = 0.55, p < 0.05; n = 743 especies). Las bayas, drupas y cápsulas loculicidas tuvieron una longitud promedio menor de 22 mm de largo, en tanto que las legumbres y cámaras fueron más largas (> de 100 mm). El número máximo de tipos de fruto por familia fue de siete (Rubiaceae) y 47 de éstas exhibieron sólo uno. La frecuencia de tipos de fruto para ésta categoría taxonómica mostró una relación significativa con el número de especies que incluye (r = 0.58, p < 0.05; n = 85 familias), lo que no aconteció a nivel de género, ya que 328 (97%) presentan una sola categoría de fruto. Los resultados indican que la morfología de los frutos de las especies arbóreas del bosque tropical perennifolio es un elemento relevante para entender su compleja composición y estructura.
Palabras clave: color de frutos, selva alta perennifolia, tamaño de frutos, tipos de fruto.
ABSTRACT
Fruit morphology (color, size and type) of 881 tree species (338 genera and 85 families) that inhabit the tropical rain forest in Mexico was assessed. We found 46 fruit types and about 52.7% of species had berries (212 species), drupes (163) or loculicidal capsules (89); ten fruit types were represented by just one species. The most abundant fruit colors were (n = 865 species) black (307 species; 35.5%), brown (227; 26.2%) and red (159; 18.4%). Berries were usually black (66%), drupes black (53.9%) or red (39.3%) and more than half of loculicidal capsules were brown (66.3%). Fruit length varied from 0.8 to 430 mm (mean 31.7 mm) and fruit width from 1 to 145 mm (mean 16.1 mm); these variables were positively correlated (r = 0.55, p < 0.05; n = 743 species). Berries, drupes and loculicidal capsules were small (mean size < 22 mm long), while legumes and camaras were longer (mean > 100 mm long). The largest numbers of fruit types per family was recorded for Rubiaceae (seven) and 47 of them displayed just one. Fruit type frequency and species per family were correlated (r = 0.58, p < 0.05; n = 85 families). Such correlation was not sustained at generic level, because 328 genera (97%) showed just one fruit type. Our results indicate that fruit morphological diversity of tree species is an important trait to understand the structure and composition of the tropical rain forest.
Key words: fruit color, fruit size, fruit type, tropical rain forest.
INTRODUCCIÓN
Desde una perspectiva amplia del concepto de biodiversidad, el bosque tropical perennifolio ("tropical rain forest") ocupa una posición destacada entre los diferentes ecosistemas terrestres de nuestro planeta, ya sea si se compara su número de especies por unidad de área o diferentes aspectos de su estructura y función (Whitmore, 1998). En este ecosistema las especies despliegan una gama muy amplia de tipos, tamaños y colores de flores y frutos (Standley y Steyermark, 1946; Standley et al., 1974; Gentry, 1982; Wheelwright et al., 1984; Corlett, 1996; Carpenter et al., 2003; Chen et al., 2004). En términos generales, el conocimiento de los frutos de las especies tropicales se ha enfocado hacia su dispersión y la influencia que este fenómeno puede desempeñar en la estructura y función de las comunidades animales y vegetales, por ejemplo, la diversificación de patrones fenológicos de especies con frutos que difieren en su vector de dispersión (anemocoría, autocoría o zoocoría) o el consumo de plantas por diferentes gremios de frugívoros como aves o primates (Gentry, 1982; Howe y Smallwood, 1982; Gautier–Hion et al., 1985; Ibarra–Manríquez y Oyama, 1992; Ibarra–Manríquez y Sinaca, 1995; Chapman y Onderdonk, 1998; Whitmore, 1998; Graham et al., 2002).
Una de las fuentes de información más frecuentemente utilizadas en esta clase de estudios ecológicos es la morfología de los frutos, principalmente su color, consistencia (seca o carnosa) o tamaño, lo que en conjunto puede ser usado para proponer síndromes de dispersión de las especies (Dansereau y Lems, 1957; Gentry, 1982, 1995; Wheelwright y Janson, 1985; Augspurger, 1986; Willson y Whelan, 1990; Ibarra–Manríquez et al., 1991; Solórzano et al., 2002). Recientemente este tipo de información se ha usado para determinar la importancia que pueden tener factores históricos (e.g. la filogenia) o ecológicos en la estructura y composición de especies del bosque tropical perennifolio (Ibarra–Manríquez et al., 2001; Chazdon et al., 2003; Chen et al., 2004). A pesar de su relevancia teórica y práctica en el entendimiento y manejo de comunidades tropicales, la información sobre la morfología de los frutos del bosque tropical perennifolio en México se encuentra dispersa en publicaciones florísticas o taxonómicas (e.g. Standley y Steyermark, 1946; Standley et al., 1974; Fryxell, 1988; Pennington, 1990; Nee, 1993; Sousa, 1993; Ibarra–Manríquez y Sinaca, 1995, 1996a, b; Pennington y Sarukhán, 2005; Barrie, 2005; Borhidi, 2006) y la literatura especializada es escasa (Niembro, 1983; Sánchez–Garfias et al., 1991; Lentz y Dickau, 2005) e incluye un porcentaje muy bajo de las 5,000 especies estimadas para el mencionado tipo de vegetación (Rzedowski, 1991a).
Debido a esta alta diversidad y al incompleto conocimiento de la morfología de sus frutos, una manera de empezar a recopilar tal información es restringirse al estudio de las especies arbóreas, ya que en conjunto con las hierbas, son las formas de crecimiento con mayor riqueza de especies a nivel local o del país en distintos tipos de comunidades vegetales, incluyendo el bosque tropical perennifolio (e.g. Meave, 1990; Rzedowski, 1991b; Ibarra–Manríquez y Sinaca, 1995; Alcántara y Luna, 2001; Castillo–Campos et al., 2008; Meave et al., 2008). Otro aspecto que justifica este enfoque es que la taxonomía y conocimiento florístico de los árboles están mejor documentados si se les compara con los que existen para otras formas de crecimiento, como por ejemplo las lianas (Gentry, 1991, 1995). Por lo tanto, las principales preguntas que plantea resolver el presente estudio son: 1) ¿Cuántos tipos de fruto presentan las especies arbóreas del bosque tropical perennifolio de México y cuáles son los más frecuentes? 2) ¿Qué variación despliega este conjunto de especies respecto al color y tamaño de sus frutos? 3) ¿Los distintos tipos de fruto tienen un color o tamaño que los caracterice? y 4) ¿Las familias y géneros más diversificados son los taxones que presentan el mayor número de tipos de frutos?
ÁREA DE ESTUDIO
En México, el bosque tropical perennifolio (BTP) se localiza desde el nivel del mar hasta los 1200 m de altitud y raramente hasta la cota de los 1500 m (Miranda y Hernández–X., 1963; Rzedowski, 1978; Challenger, 1988). El BTP se distribuye principalmente en las provincias fisiográficas Planicie Costera Nororiental, Planicie Costera Suroriental y Sistema Montañoso del Norte de Oaxaca (Rzedowski, 1978), en 11 estados localizados principalmente hacia el sureste del país (Fig. 1). Este tipo de vegetación se desarrolla en climas cálidos (tipo A de Köppen), con una temperatura media anual de al menos 22 °C y la del mes más frío superior a los 18 °C, con precipitaciones anuales entre 1100–2000 mm, aunque en algunas localidades sobrepasa los 4000 mm, sin una época de sequía o con un máximo de tres meses secos, excepto en la Península de Yucatán, donde este período alcanza 4 o 5 meses (Rzedowski, 1978; Wendt, 1993; Challenger, 1988; Pennington y Sarukhán, 2005).

El BTP incluye comunidades vegetales densas, con más de 75% de sus especies perennifolias durante la temporada con menor precipitación, en las cuales los árboles del dosel despliegan una altura considerable (25–40 m en promedio) y con una abundancia importante de lianas, epífitas y palmas (Miranda y Hernández–X., 1963; Rzedowski, 1978; Meave, 1990; Bongers et al., 1992; Wendt, 1993; Ibarra–Manríquez et al., 1997; Pennington y Sarukhán, 2005; Meave et al., 2008). Entre las especies de mayor tamaño y que destacan por su abundancia local están Alchornea latifolia Sw. (Euphorbiaceae), Ampelocera hottlei (Standl.) Standl. (Ulmaceae), Apeiba tibourbou Aubl. (Tiliaceae), Brosimum alicastrum Sw. (Moraceae), Bursera simaruba (L.) Sarg. (Burseraceae), Calophyllum brasiliense Cambess. (Clusiaceae), Cecropia obtusifolia Bertol. (Cecropiaceae), Ceiba pentandra (L.) Gaertn. (Bombacaceae), Cojoba arborea (L.) Britton & Rose (Fabaceae), Cymbopetalum baillonii R.E. Fr. (Annonaceae), Dendropanax arboreus (L.) Decne. & Planch. (Araliaceae), Dialium guianense (Aubl.) Sandwith (Fabaceae), Erblichia odorata Seem. (Turneraceae), Ficus aurea Nutt., F. insipida Willd., F. yoponensis Desv. (Moraceae), Guarea glabra Vahl, G. grandifolia DC. (Meliaceae), Guatteria anomala R.E. Fr. (Annonaceae), Handroanthus guayacan (Seem.) S.O. Grose (Bignoniaceae), Heliocarpus appendiculatus Turcz. (Tiliaceae), Licania platypus (Hemsl.) Fritsch, L. sparsipilis S.F. Blake (Chrysobalanaceae), Lonchocarpus cruentus Lundell (Fabaceae), Manilkara zapota (L.) P. Royen (Sapotaceae), Nectandra ambigens (S.F. Blake) C.K. Allen (Lauraceae), Ochroma pyramidale (Cav. ex Lam.) Urb. (Bombacaceae), Poulsenia armata (Miq.) Standl. (Moraceae), Pouteria campechiana (Kunth) Baehni (Sapotaceae), Pseudolmedia glabrata (Liebm.) C.C. Berg (Moraceae), Quararibea funebris (La Llave) Vischer (Bombacaceae), Swietenia macrophylla King (Meliaceae), Terminalia amazonia (J.F. Gmel.) Exell (Combretaceae), Trema micrantha (L.) Blume (Ulmaceae), Ulmus mexicana (Liebm.) Planch. (Ulmaceae), Vatairea lundellii (Standl.) Killip ex Record (Fabaceae) y Vochysia guatemalensis Donn. Sm. (Vochysiaceae).
MÉTODOS
Se realizó una extensa revisión de literatura taxonómica y florística para ubicar las especies de árboles que se mencionan como parte del BTP de México (e.g. Standley, 1920–1926; Pennington, 1990; Nee, 1993; Sousa, 1993; Wendt, 1993; Henderson et al., 1995; Ibarra–Manríquez y Sinaca, 1995, 1996a, b; Barneby, 1998; Fryxell, 1988; Lorea–Hernández, 2002; Ricketson y Pipoly, 2003; Plowman y Hensold, 2004; Barrie, 2005; Keller, 2005; Borhidi, 2006; Rico–Arce, 2007; Meave et al., 2008). Las especies incluidas en el presente estudio son descritas en la literatura y en los ejemplares de herbario revisados como árboles (a veces también como arbustos) y tienen una altura mínima de 3 m (Apéndice), pero con la capacidad de alcanzar hasta 87 m (Ulmus mexicana). A pesar de no ser estrictamente árboles, también se incluyen algunas especies arborescentes de Arecaceae.
Debido a la complejidad para definir claramente el BTP con respecto a otras coberturas vegetales en México (e.g. el bosque mesófilo de montaña o el bosque tropical subcaducifolio a subperennifolio), con las cuales puede confundirse o formar gradientes complejos (Miranda y Hernández–X., 1963; Rzedowski, 1978; Challenger, 1988; Pennington y Sarukhán, 2005), así como a la ambigüedad con que en ocasiones se indica el tipo de vegetación de una especie en la literatura o en las colecciones de herbario, la incorporación de una especie al presente estudio tuvo como requisitos complementarios: i) que su distribución geográfica fuera coincidente con los estados que indicó Rzedowski (1978) para el BTP (Fig. 1) y ii) y que altitudinalmente se encontrara entre el nivel del mar y los 1000 m s.n.m., independientemente de si se restringe o no a este intervalo de altitud.
El tipo, tamaño (largo y ancho) y color de los frutos de las especies enlistadas en el Apéndice fue obtenido de la literatura revisada, la consulta de especimenes depositados en distintos herbarios (CHAPA, F, IEB, MEXU, MO y XAL) y por observaciones de campo realizadas por el primer autor. El tipo de fruto de las especies fue determinado con base en la clasificación de Spjut (1994), que definen 95 categorías y permite describir con mayor precisión la diversidad morfológica que muestra esta estructura en las comunidades tropicales del mundo. Este sistema y sus lineamientos básicos pueden consultarse directamente en Spjut (1994) o también en http://www.worldbotanical.com/fruit_types.htm. Cabe destacar que el mencionado autor clasifica las especies con cápsulas de acuerdo con su tipo de dehiscencia (e.g. loculicida o septicida). Sin embargo, para algunos de los árboles incluidos en el Apéndice no fue posible determinar con certeza este atributo y se mencionan entonces únicamente como cápsulas. Otras categorías que se manejan en el presente trabajo y que no son contempladas por Spjut (1994), son las legumbres indehiscentes, presentes en algunas especies de Caesalpiniaceae y Fabaceae, así como las cápsulas con deshiscencia apical (apicidas) encontradas en el género Renistipula (Rubiaceae).
Por otro lado, en Annonaceae y Capparaceae existen taxones aún sin describir para la ciencia. En el caso de la primera familia se trata de especies de los géneros Guamia Merr. y Desmopsis Saff. (denominadas en el Apéndice como Guamia sp. y Desmopsis sp., respectivamente), mientras que en Capparaceae esta entidad se denomina Capparis sp. El arreglo de las familias en el Apéndice se basa en la clasificación de Cronquist (1981).
RESULTADOS
En el presente trabajo se registraron 881 especies que se distribuyen en 338 géneros y 85 familias (Apéndice). A pesar de que en conjunto fueron reconocidos 46 tipos de frutos (Cuadro 1), su frecuencia entre los árboles estudiados es desigual, ya que un poco más de la mitad (52.6%) tienen bayas (212, 24%), drupas (163, 18.5%) o cápsulas loculicidas (89, 10.1%), mientras que 10 tipos de fruto fueron encontrados sólo en una. El color del fruto fue caracterizado para 865 especies, de las cuales 93.6% despliega alguno de los siguientes (Cuadro 1): negro (307 especies; 35.5%), pardo (227; 26.2%), rojo (159; 18.4%) y verde (117; 13.5%). Los tres tipos de frutos mencionados previamente como los más frecuentes fueron los únicos en presentar los ocho colores reconocidos en el presente estudio. Sin embargo, en el Cuadro 1 se observa que en las bayas el color más usual fue el negro (66%) y en las cápsulas loculicidas el pardo (66.3%), en tanto que las drupas pueden ser negras (53.9%) o rojas (39.3%). Otros frutos con colores predominantes fueron las glandes y sámaras, ya que 87.9% de las primeras son negras, en tanto que 89.5% de las segundas son pardas. Un total de 14 tipos de frutos presentaron un solo color (Cuadro 1).

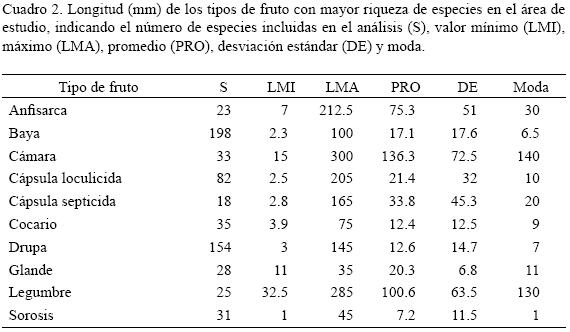
El largo del fruto entre las especies incluidas en el estudio varió de 0.8 mm (Piper aduncum, Piperaceae) a 430 mm (Handroanthus guayacan, Bignoniaceae), con un promedio de 31.7 mm. El ancho mostró un intervalo menor, desde 1 mm (varios árboles) hasta 145 mm (Carica papaya, Caricaceae), con un promedio de 16.1 mm. Estas dos características están significativamente correlacionadas (r = 0.55, p < 0.05; n = 743 especies). A pesar de que la longitud de los tipos de fruto con mayor riqueza de especies tiene una variación alta, seis son relativamente pequeños, ya que presentan un promedio y una moda menor de 22 mm (Cuadro 2); únicamente las legumbres y cámaras presentan para estas dos últimas medidas un valor que sobrepasa 100 mm. No se discute la variación del ancho de los frutos debido a su correlación significativa con la longitud.
El número mayor de tipos de frutos (siete) fue registrado en Rubiaceae y uno solo se halló en 47 familias (Apéndice). A este nivel taxonómico se reveló una correlación significativa entre el número de especies y el de los tipos de fruto que presenta (r = 0.58, p < 0.05; n = 85 familias). A pesar de lo anterior, es de llamar la atención que entre las 20 familias con mayor riqueza de especies, en la mitad se censan únicamente 1 o 2 categorías carpológicas diferentes (Cuadro 3).

No se encontró una relación significativa entre los tipos de fruto y la riqueza de especies de los géneros (r = 0.23, p < 0.05; n = 338 géneros). Únicamente Capparis (Capparaceae), Cordia (Boraginaceae), Lonchocarpus (Fabaceae), Diospyros (Ebenaceae), Ocotea (Lauraceae), Pouteria (Sapotaceae), Robinsonella (Malvaceae), Senna (Caesalpiniaceae), Talisia (Sapindaceae) y Urera (Urticaceae) presentaron dos tipos de fruto y el resto de los miembros de esta categoría supraespecífica tuvieron sólo uno (Apéndice), a pesar de que ocho se ubican entre los que tienen más de 10 especies: Miconia (25 especies, bayas), Eugenia (23, bayas), Inga (20, cámaras), Piper (18, sorosis), Psychotria (18, drupas), Ficus (17, siconos), Nectandra (15, bayas), Croton (12, cocarios), Ardisia (13, drupas), Calypthranthes (12, bayas) y Solanum (10, bayas).
DISCUSIÓN
Aunque determinar la riqueza de especies del bosque tropical perennifolio mexicano no es uno de los objetivos del presente estudio, la lista de 881 (Apéndice) casi dobla las 451 incluidas por Wendt (1993) en su análisis biogeográfico restringido a los árboles del dosel de este tipo de vegetación, abarcando 35.3% de las 2500 que Rzedowski (1991b) estimó para esta forma de crecimiento en México. Esta alta diversidad se ve reflejada en el número de tipos de frutos encontrado, que representa casi la mitad (47.3%) del total que propone Spjut (1994). Sin embargo, es importante destacar el predominio de 10 categorías (Cuadro 1), que en su conjunto engloban 76.9% del total de los árboles estudiados, especialmente las bayas y drupas (24 y 18.6%, respectivamente). Chen et al. (2004) también encontraron que entre 412 especies del BTP en China, pertenecientes a diferentes formas de crecimiento, las drupas (47%) y bayas (32%) fueron las más frecuentes. Previamente Carpenter et al. (2003) indicaron que 59% de 116 diferentes árboles en Nueva Caledonia poseen bayas, drupas o siconos. Tal porcentaje es cercano al obtenido en el presente trabajo, ya que 44.5% de las especies mostraron estos tipos de frutos (Cuadro 1); otra similitud entre los dos estudios es la rareza de algunas modalidades, como por ejemplo, las sámaras.
Con respecto al color, Carpenter et al. (2003) encontraron que los más frecuentes son el pardo (39%), rojo (28%), así como los azules, negros o púrpuras (23%); estas cifras son de alguna manera distintas de las encontradas en México, donde los predominantes corresponden al negro (35.5%), pardo (26.2%) y rojo (18.4%). En forma similar, Corlett (1996) señala que entre las especies dispersadas por vertebrados en China, los colores más importantes son el negro (45.1%) y el rojo (24.4%). En este mismo país, Chen et al. (2004) también encontraron que los frutos negros son los más frecuentes con 40% (n = 389), siguiendo en orden decreciente los rojos, pardos y amarillos (19%, 13% y 13%, respectivamente). Tales tendencias de coloración de los frutos en áreas tropicales y subtropicales han sido relacionadas con las plantas que presentan frutos carnosos (e.g. bayas, drupas o siconos), cuyos vectores de dispersión son animales (van der Pijl, 1972; Willson y Whelan, 1990; Chen et al., 2004).
La longitud promedio de los frutos del bosque tropical perennifolio de México fue de 31.7 mm, número cercano al encontrado por Carpenter et al. (2003), de 24.9 mm. En el presente estudio, este atributo difiere fuertemente entre los tipos de fruto más frecuentes (Cuadro 2), en especial si se comparan los de promedio más pequeño (baya y sorosis) con los de mayor valor (cámara y legumbre), discrepancias también muy notorias a nivel de las longitudes mínimas y máximas. A este respecto, Chen et al. (2004) separaron los frutos carnosos en tres categorías de tamaño, encontrando que los pequeños (< 20 mm) representaron 69% (277 especies), los medianos (20–50 mm) un 26% y los grandes (> 50 mm) solamente 5%. Si se considera el largo máximo que pueden alcanzar las frutos de los árboles mexicanos, puede afirmarse que también son predominantemente pequeños (467 especies, 62.9%), mientras que los medianos y grandes muestran proporciones similares (18.7 y 18.4%). Entre los tipos de frutos citados en el Cuadro 2, cuatro son carnosos (baya, drupa, glande (excepto las dos especies de Quercus) y sorosis) y es interesante que todos poseen promedios y modas menores de 20.5 mm. Finalmente, ante el hecho de que 62.9% de las especies arbóreas del BTP mexicano tienen frutos pequeños y que 54.1% son negros o rojos (Cuadro 1), es posible plantear como hipótesis que este conjunto de taxa podrían ser dispersados preferentemente por aves, tal y como ha sido postulado en trabajos realizados en otros bosques tropicales del mundo (van der Pijl, 1972; Stiles, 2000; Ganesh y Davidar, 2001; Carpenter et al., 2003; Chen et al., 2004; Link y Stevenson, 2004).
Por otro lado, diversos estudios han documentando que algunos atributos reproductivos que se observan a nivel de género o familia, por ejemplo el tamaño de las semillas, la proporción entre el ancho y largo de los frutos, el tipo de plántulas o el modo en que sus diásporas son dispersadas, se asocian fuertemente con aquellos que se detectan en las especies que incluyen estas categorías taxonómicas (Herrera, 1992; Ibarra–Manríquez y Oyama, 1992; Leishman et al., 2000; Willson y Traveset, 2000; Ibarra–Manríquez et al., 2001; Prinzing et al., 2001; Webb et al., 2002; Chazdon et al., 2003; Chen et al., 2004; Bolmgren y Eriksson, 2005). En el presente trabajo se encontró una correlación estadísticamente significativa a nivel de familia entre su riqueza de especies y la cantidad de las categorías carpológicas que incluyen, lo que no se obtuvo con los géneros. Lo anterior sugiere que la historia evolutiva de las familias en el bosque tropical perennifolio de México desempeña un papel importante en los tipos de frutos encontrados en este tipo de vegetación, tal y como ha sido documentado para otras comunidades tropicales (Webb et al., 2002; Chazdon et al., 2003; Chen et al., 2004; Bolmgren y Eriksson, 2005), lo cual tiene una importancia fundamental para entender diversos aspectos de su composición, estructura y funcionamiento que se observan hoy en día.
Un corolario de lo hasta ahora discutido es que se requiere avanzar en el conocimiento de los atributos de los frutos de las especies mexicanas, fundamentalmente en la caracterización de colores, dimensiones y pesos que poseen, lo que eventualmente debería de extenderse a otros atributos reproductivos (e.g. color y tamaño de flores, número de semillas por fruto, tipos de plántula), a otras formas de crecimiento (e.g. epífitas o lianas) y a otros tipos de vegetación de afinidad tropical, subtropical y templada. El presente trabajo muestra que existe un número alto de diferentes categorías de frutos en las especies arbóreas del BTP de México, que podría usarse como un criterio adicional para demostrar que se trata de uno de los ecosistemas con mayor diversidad biológica y justificar las acciones que deben hacerse para asegurar su conservación futura.
AGRADECIMIENTOS
Parte de esta investigación fue desarrollada durante el año sabático del primer autor, el cual contó con una beca otorgada por la Dirección General de Asuntos del Personal Académico (DGAPA), de la Universidad Nacional Autónoma de México. Se agradecen las facilidades para el desarrollo de la presente contribución por parte de los jefes de los herbarios CHAPA, F, IEB, MEXU, MO y XAL. Un reconocimiento especial para Robin B. Foster, de Field Museum of Chicago, Illinois, EUA, ya que nos permitió la consulta irrestricta de su acervo bibliográfico especializado sobre las plantas del Neotrópico, a José de Jesús Alfonso Fuentes Junco por la elaboración de la figura 1 y a Juan Martínez Cruz por su apoyo en el trabajo de campo y en la obtención de información de los frutos de algunas de las especies incluidas en el trabajo. Finalmente agradecemos las atinadas correcciones del Dr. Jerzy Rzedowski y dos revisores anónimos que permitieron mejorar la redacción y estructura del presente trabajo.
LITERATURA CITADA
Alcántara, O. e I. Luna. 2001. Análisis florístico de dos áreas con bosque mesófilo de montaña en el estado de Hidalgo, México: Eloxochitlán y Tlahuelompa. Acta Bot. Mex. 54: 51–87. [ Links ]
Augspurger, C. K. 1986. Morphology and dispersal potential of wind–dispersed diaspores of Neotropical trees. Amer. J. Bot. 73: 353–363. [ Links ]
Barneby, R. C. 1998. Silk tree, guanacaste, monkey's earring. A generic system for the synandrous Mimosaceae of the Americas. Part III. Calliandra. Mem. New York Bot. Gard. 74(3): 1–223. [ Links ]
Barrie, F. R. 2005. Thirty–five new species of Eugenia (Myrtaceae) from Mesoamerica. Novon 15: 4–49. [ Links ]
Bongers, F., J. Popma, J. Meave del Castillo y J. Carabias. 1992. Structure and floristic composition of the lowland rain forest of Los Tuxtlas, Mexico. Vegetatio 74: 55–80. [ Links ]
Bolmgren, K. y O. Eriksson. 2005. Fleshy fruits – origins, niche shifts, and diversification. Oikos 109: 255–72. [ Links ]
Borhidi, A. 2006. Rubiáceas de México. Academia de Ciencias de Hungría. Budapest, Hungría. 512 pp. [ Links ]
Carpenter, R. J., J. Read y T. Jaffré. 2003. Reproductive traits of tropical rain–forest trees in New Caledonia. J. Trop. Ecol. 19: 351–365. [ Links ]
Castillo–Campos, G., G. Halffter y C. E. Moreno. 2008. Primary and secondary vegetation patches as contributors to floristic diversity in a tropical deciduous forest landscape. Biodivers. Conser. 17: 1701–1714. [ Links ]
Challenger, A. 1988. Utilización y conservación de los ecosistemas terrestres de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad e Instituto de Biología, de la Universidad Nacional Autónoma de México. México, D.F. 847 pp. [ Links ]
Chapman, C. A. y D. A. Onderdonk. 1998. Forests without primates: primate/plant codependency. Amer. J. Primatol. 45: 127–141. [ Links ]
Chazdon, R. L., S. Careaga, C. Webb y O. Vargas. 2003. Community and phylogenetic structure of reproductive traits of woody species in wet tropical forests. Ecol. Monogr. 73: 331–348. [ Links ]
Chen, J., T. H. Fleming, L. Zhang, H. Wang y Y. Liu. 2004. Patterns of fruits in a tropical rain forest in Xishuangbanna, SW China. Acta Oecol. 26: 157–164. [ Links ]
Corlett, R. T. 1996. Characteristics of vertebrate dispersal fruits in Hong Kong. J. Trop. Ecol. 12: 819–833. [ Links ]
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press. Nueva York. 1202 pp. [ Links ]
Dansereau, P. y K. Lems. 1957. The grading of dispersal types in plant communities and their ecological significance. Contr. Inst. Bot. Univ. Montreal 71: 1–52. [ Links ]
Fryxell, P. A. 1988. Malvaceae of Mexico. Syst. Bot. Monogr. 25: 1–520. [ Links ]
Ganesh, T. y P. Davidar. 2001. Dispersal modes of tree species in the wet forest of southern Western Ghats. Curr. Sci. India 80: 394–399. [ Links ]
Gautier–Hion, A., J.–M. Duplantier, R. Quris, F. Feer, C. Sourd, J.–P. Decoux, G. Dubost, L. Emmons, C. Erard, P. Hecketsweiler, A. Moungazi, C. Roussilhon y J.–M. Thiollay. 1985. Fruit characters as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community. Oecologia (Berlin) 65: 324–337. [ Links ]
Gentry, A. H. 1982. Patterns of Neotropical plant species diversity. Evol. Biol. 15: 1–84. [ Links ]
Gentry, A. H. 1991. The distribution and evolution of climbing plants. In: Putz, F. E. y H. A. Mooney (eds.). The biology of vines. Cambridge University Press. Cambridge. pp. 3–49. [ Links ]
Gentry, A. H. 1995. Diversity and floristic composition of Neotropical dry forests. In: Bullock, S. H., H. A. Mooney y E. Medina (eds.). Seasonally dry tropical forests. Cambridge University Press. Cambridge. pp. 146–190. [ Links ]
Graham, C., J. E. Martínez–Leyva y L. Cruz–Paredes. 2002. Use of fruiting trees by birds in continuous forest and riparian forest remnants in Los Tuxtlas, Veracruz, Mexico. Biotropica 34: 589–597. [ Links ]
Henderson, A., G. Galeano y R. Bernal. 1995. Palms of the Americas. Princeton University Press. New Jersey. 352 pp. [ Links ]
Herrera, C. M. 1992. Interspecific variation in fruit shape: allometry, phylogeny, and adaptation to dispersal agents. Ecology 73: 1832–1841. [ Links ]
Howe, H. F. y J. Smallwood. 1982. Ecology of seed dispersal. Ann. Rev. Ecol. Syst. 13: 201–228. [ Links ]
Ibarra–Manríquez, G., B. Sánchez–Garfias y L. González–García. 1991. Fenología de lianas y árboles anemócoros en una selva cálido–húmeda de México. Biotropica 23: 242–254. [ Links ]
Ibarra–Manríquez, G. y K. Oyama. 1992. Ecological correlates of reproductive traits of Mexican rain forest trees. Amer. J. Bot. 79: 344–356. [ Links ]
Ibarra–Manríquez, G. y S. Sinaca C. 1995. Lista florística comentada de la Estación de Biología Tropical "Los Tuxtlas", Veracruz, México. Rev. Biol. Trop. 43: 75–115. [ Links ]
Ibarra–Manríquez, G. y S. Sinaca C. 1996a. Estación de Biología Tropical "Los Tuxtlas", Veracruz, México: lista florística comentada (Mimosaceae a Verbenaceae). Rev. Biol. Trop. 44: 41–60. [ Links ]
Ibarra–Manríquez, G. y S. Sinaca C. 1996b. Estación de Biología Tropical "Los Tuxtlas", Veracruz, México: lista florística comentada (Violaceae a Zingiberaceae). Rev. Biol. Trop. 44: 427–447. [ Links ]
Ibarra–Manríquez, G., M. Martínez–Ramos, R. Dirzo y J. Núñez–Farfán. 1997. La vegetación. In: González–Soriano, E., R. Dirzo y R. C. Vogt (eds.). Historia natural de Los Tuxtlas, Instituto de Biología e Instituto de Ecología, Universidad Nacional Autónoma de México. México, D.F. pp. 61–85. [ Links ]
Ibarra–Manríquez, G., M. Martínez–Ramos y K. Oyama. 2001. Seedling functional types in a lowland rain forest in Mexico. Amer. J. Bot. 88: 1801–1812. [ Links ]
Keller, B. T. 2005. Actinidiaceae. Flora of Chiapas 6: 1–14. [ Links ]
Leishman, M. R., I. J. Wright, A. T. Moles y M. Westoby. 2000. The evolutionary ecology of seed size. In: Fenner, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2nd. ed. CAB International. Wallingford. pp. 31–57. [ Links ]
Lentz, D. L. y R. Dickau. 2005. Seeds of Central America and Southern Mexico. Mem. New York Bot. Gard. 91: 1–296. [ Links ]
Link, A. y P. R. Stevenson. 2004. Fruit dispersal syndromes in animal disseminated plants at Tinigua National Park, Colombia. Rev. Chil. Hist. Nat. 77: 319–334. [ Links ]
Lorea–Hernández, F. G. 2002. La familia Lauraceae en el sur de México: diversidad, distribución y estado de conservación. Bol. Soc. Bot. Méx. 71: 59–70. [ Links ]
Meave, J. A. 1990. Estructura y composición de la selva alta perennifolia de los alrededores de Bonampak. Serie Arqueología. Instituto Nacional de Antropología e Historia. México, D.F. 147 pp. [ Links ]
Meave, J. A., M. A. Romero–Romero, A. Valle–Doménech, A. Rincón–Gutiérrez, E. Martínez y C. H. Ramos. 2008. Plant diversity assessment in the Yaxchilán Natural Monument, Chiapas, Mexico. Bol. Soc. Bot. Méx. 83: 53–76. [ Links ]
Miranda, F. y E. Hernández–X. 1963. Los tipos de vegetación de México y su clasificación. Bol. Soc. Bot. Méx. 28: 29–178. [ Links ]
Nee, M. 1993. Solanaceae II. Flora de Veracruz 72: 1–158. [ Links ]
Niembro, R. A. 1983. Caracterización morfológica y anatómica de semillas forestales. Departamento de Bosques, Universidad Autónoma de Chapingo. Chapingo, México. 211 pp. [ Links ]
Pennington, T. D. 1990. Sapotaceae. Flora Neotropica 52: 1–770. [ Links ]
Pennington, T. D. y J. Sarukhán. 2005. Árboles tropicales de México. Manual para la identificación de las principales especies. 3a. ed. Universidad Nacional Autónoma de México y Fondo de Cultura Económica. México, D.F. 523 pp. [ Links ]
Plowman, T. y N. Hensold. 2004. Names, types, and distribution of Neotropical species of Erythroxylum (Erythroxylaceae). Brittonia 56: 1–53. [ Links ]
Prinzing, A., W. Durka, S. Klotz y R. Brandl. 2001. The niche of higher plants: evidence for phylogenetic conservatism. Proc. R. Soc. Ser. B–Bio. 268: 2383–2389. [ Links ]
Rico–Arce, M. L. 2007. A checklist and synopsis of American species of Acacia (Leguminosae: Mimosoideae). Royal Botanic Gardens Kew y Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, D.F. 207 pp. [ Links ]
Ricketson, J. M. y J. J. Pipoly III. 2003. Revision of Ardisia subgenus Auriculardisia (Myrsinaceae). Ann. Mo. Bot. Gard. 90: 179–317. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Ed. Limusa. México, D.F. 432 pp. [ Links ]
Rzedowski, J. 1991a. Diversidad y orígenes de la flora fanerogámica de México. Acta Bot. Mex. 14: 3–21. [ Links ]
Rzedowski, J. 1991b. El endemismo en la flora fanerogámica mexicana: una apreciación analítica preliminar. Acta Bot. Mex. 15: 47–64. [ Links ]
Sánchez–Garfias, B., G. Ibarra–Manríquez y L. González–García. 1991. Manual de identificación de frutos y semillas anemócoros de árboles y lianas de la Estación "Los Tuxtlas", Veracruz, México. Cuadernos 12. Instituto de Biología, Universidad Nacional Autónoma de México. México, D.F. 86 pp. [ Links ]
Solórzano, S., G. Ibarra–Manríquez y K. Oyama. 2002. Liana diversity and reproductive attributes in two tropical forests in Mexico. Biodivers. Conser. 11: 197–212. [ Links ]
Sousa, S, M. 1993. El género Inga (Leguminosae: Mimosoideae) del sur de México y Centroamérica, estudio previo para la Flora Mesoamericana. Ann. Mo. Bot. Gard. 80(1): 223–269. [ Links ]
Spjut, R. W. 1994. A systematic treatment of fruit types. Mem. New York Bot. Gard. 70: 1–182. [ Links ]
Standley, P. C. y J. A. Steyermark. 1946. Flora of Guatemala. Ulmaceae to Krameriaceae. Fieldiana Bot. 24(4): 1–493. [ Links ]
Standley, P. C., L. O. Williams y G. D. Nash. 1974. Flora of Guatemala. Bignoniaceae to Plantaginaceae. Fieldiana Bot. 24(10): 153–466. [ Links ]
Standley, P. C. 1920–26. Trees and shrubs of Mexico. Contrib. U. S. Nat. Herb. 23: 1–1721. [ Links ]
Stiles, E. W. 2000. Animals as seed dispersers. In: Fenner, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2nd. ed. CAB International. Wallingford. pp. 111–124. [ Links ]
van der Pijl, L. 1972. Principles of dispersal in higher plants. 2a. ed. Springer–Verlag. Berlin. 162 pp. [ Links ]
Webb, C. O., D. D. Ackerly, M. A. McPeek y M. J. Donoghue. 2002. Phylogenies and community ecology. Ann. Rev. Ecol. Syst. 33: 475–505. [ Links ]
Wendt, T. 1993. Composition, floristic affinities, and origins of the canopy tree flora of the Mexican Atlantic slope rain forest. In: Ramamoorthy T. P., R. Bye, A. Lot y J. Fa (comp.). Biological diversity of Mexico: origins and distribution. Oxford University Press. Nueva York. pp. 595–680. [ Links ]
Wheelwright, N. T., W. A. Haber, K. G. Murray y C. Guindon. 1984. Tropical fruit eating–birds and their food plants: a survey of a Costa Rican lower montane forest. Biotropica 16: 173–192. [ Links ]
Wheelwright, N. T. y C. H. Janson. 1985. Colors of fruits displays of bird–dispersed plants in two tropical forests. Amer. Nat. 126: 777–799. [ Links ]
Whitmore, T. C. 1998. An introduction to tropical rain forest. Oxford University. Nueva York. 282 pp. [ Links ]
Willson, M. F. y A. Traveset. 2000. The ecology of seed dispersal. In: Fenner, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2nd. ed. CAB International. Wallingford. pp. 125–165. [ Links ]
Willson, M. F. y C. J. Whelan. 1990. The evolution of fruit color in fleshy–fruited plants. Amer. Nat. 136: 790–809. [ Links ]
* Este trabajo está dedicado a la memoria del Dr. Rogers McVaugh: una vida dedicada a promover de manera incansable el conocimiento de las plantas mexicanas.

