











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
En los sistemas forestales, la biodiversidad de plantas se ha relacionado con la productividad de estos ecosistemas en forma positiva (Liang et al., 2016), por lo que es importante la caracterización con métricas de biodiversidad asociadas. La riqueza (número) de especies es una de las métricas más ampliamente usadas para evaluar la diversidad en bosques y otras comunidades vegetales (NRC, 2000; Newton y Kapos, 2002), particularmente con relación a diferentes usos del suelo, al desarrollar índices de biodiversidad en función de un estado de referencia (vegetación sucesionalmente madura) para analizar los cambios (NRC, 2000; Croezen, Bergsma, Clemens, Sevensteen y Tulleners, 2011). La evaluación indirecta de especies es la base para evaluar la diversidad de otros grupos biológicos (Lindenmayer y Likens, 2011), particularmente con base a conocimiento de relaciones ecológicas.
El uso de las plantas vasculares para el desarrollo de métricas de diversidad vegetal se utiliza ampliamente por su facilidad de medición, dado que representa la mayor parte de la biomasa forestal, además de generar múltiples hábitats para definir relaciones ecológicas con diversos grupos biológicos (Santi et al., 2010; Gao, Nielsen y Hedblom, 2015).
Ante cambios ambientales, la riqueza de especies no influye directamente en las respuestas de la vegetación, contrario a los grupos o tipos funcionales (Phillips, Gentry, Hail, Sawyer y Vazquez, 1994; Tilman, Knops, Wedin y Reich, 2002), los cuales son conjuntos de individuos con ciertas características morfológicas, fisiológicas y adaptativas comunes, que brindan una respuesta especifica al ambiente, independientemente de su afinidad taxonómica (Díaz y Cabido, 2001). Los tipos funcionales de plantas plantean una respuesta relativamente similar al ambiente (Shugart, 1996; Díaz y Cabido, 2001). La correlación entre las métricas de diversidad funcional con la riqueza de especies no siempre está establecida (Díaz y Cabido, 2001; Hooper et al., 2005).
Entre los múltiples enfoques utilizados para caracterizar tipos funcionales de los atributos de las plantas vasculares, el sistema de clasificación de Gillison es útil (Gillison, 1981, 2002 y 2006; Gillison y Carpenter, 1997), ya que es sencillo y práctico. Los atributos de la estructura fotosintética (tamaño e inclinación de las hojas, clorotipo y tipo de hoja) y de la estructura vascular de soporte (forma de vida y tipo de raíz) está considerada en este sistema. Así, la riqueza de tipos funcionales (Sg) de plantas vasculares plantea la evaluación de la redundancia funcional en la vegetación. Con relación a otros tipos de diversidad, la relación S/Sg está asociada a la riqueza de especies de termitas, aves y escarabajos (Gillison, Jones, Susilo y Bignell, 2003; Gillison y Liswanti, 2004; Gillison et al., 2013).
El área de muestreo define la riqueza de especies (Rosenzweig, 1995), por lo que es necesario definir la relación funcional entre el área de muestreo (A) y la riqueza de especies (S). Para el caso de la riqueza de tipos funcionales (Sg), la relación Sg-A esta bien establecida (Gillison, 2002), en forma similar a la relación S-A. De las contadas relaciones ecológicas generales, la relación riqueza de especies y área de muestreo es una de las pocas con amplio soporte empírico (Lawton, 1999; Lomolino, 2000).
La búsqueda de modelos de la relación S-A ha sido analizada previamente (Tjørve, Matthews y Whittaker, 2021a), por lo que en la literatura hay diferentes relaciones matemáticas para la relación riqueza-área (Scheiner, 2003; Tjørve, 2003, 2009 y 2011; Dengler, 2009; Williams, Lamont y Henstridge, 2009), aunque no existe un modelo universalmente aceptable de los treinta modelos propuestos (Tjørve y Tjørve, 2021b). El modelo potencial de Arrhenius (1921, 1923a) y el logarítmico de Gleason (1922, 1925) son de los más comunes en su uso, lo que ha generado controversia con relación a su calidad (Gleason, 1922; Arrhenius, 1923b). No obstante, en las evaluaciones empíricas, el modelo potencial ha presentado mejores resultados en los ajustes (Williams et al., 2009; Dengler, 2009; Tjørve, 2009).
En este trabajo se evalúan dos enfoques de modelación de las relaciones S-A y Sg-A, una de tipo empírico y otra en función de cinéticas de orden n, para el desarrollo de un modelo genérico que tiene como caso particular los modelos potencial y logarítmico. Los modelos presentados se utilizan para analizar la relación S-Sg asociada a las relaciones S-A y Sg-A. Lo anterior permite el uso de modelos simplificados en las aplicaciones, además de unificar el uso de modelos.
Materiales y Métodos
Base de datos de riqueza-área
Para analizar las parametrizaciones de los modelos S-A y Sg-A se utilizaron las mediciones en transectos de diferentes usos del suelo (Gillison, 2002, 2004 y 2006), los cuales consisten de ocho parcelas de muestreo contiguas de 5 m × 5 m, con un área total de 200 m2. Los 30 transectos considerados en la base de datos riqueza-área están mostrados en el Cuadro 1, generado a partir de publicaciones de datos en la literatura.
Table 1: Characteristics of the species-richness database in global literature.
| País | Uso de suelo | No. Transecto | S | Sg | S/Sg |
| India | Pastizal | 1 | 46 | 25 | 1.840 |
| India | Bosque degradado | 2 | 72 | 47 | 1.532 |
| India | Sabana | 5 | 107 | 27 | 3.963 |
| India | Bosque | 6 | 107 | 74 | 1.446 |
| India | Monocultivo (arroz) | 11 | 18 | 14 | 1.286 |
| India | Plantación | 14 | 30 | 22 | 1.364 |
| Indonesia | Selva | 5 | 115 | 37 | 3.108 |
| Indonesia | Bosque secundario (tiradero de leños) | 3 | 52 | 27 | 1.926 |
| Indonesia | Sabana | 12 | 14 | 10 | 1.400 |
| Indonesia | Sistemas agroforestales (selva de caucho) | 10 | 115 | 47 | 2.447 |
| Indonesia | Plantaciones (caucho) | 6 | 69 | 38 | 1.816 |
| Indonesia | Plantaciones | 7 | 49 | 33 | 1.485 |
| Indonesia | Mandioca (3 años) | 14 | 17 | 15 | 1.133 |
| Indonesia | Zona de barbecho (Chromolaena - 3 años) | 16 | 42 | 33 | 1.273 |
| Nueva Guinea | Selva | 2 | 120 | 52 | 2.308 |
| Nueva Guinea | Bosque secundario | 1-1965 | 90 | 55 | 1.636 |
| Nueva Guinea | Plantaciones (aceite de palma) | 4 | 23 | 23 | 1.000 |
| Brasil | Selva (bosque intacto/arena blanca) | 24 | 43 | 26 | 1.654 |
| Brasil | Sabana (matorral) | 22 | 94 | 39 | 2.410 |
| Brasil | Plantaciones (aceite de palma) | 5 | 24 | 20 | 1.200 |
| Camerún | Bosque secundario | 1 | 100 | 38 | 2.632 |
| Camerún | Sabana (matorral) | 17 | 45 | 40 | 1.125 |
| Camerún | Sistemas agroforestales (jungle cocon) | 10 | 79 | 48 | 1.646 |
| Camerún | Plantaciones (cacao) | 15 | 65 | 45 | 1.444 |
| Camerún | Mandioca (2 años) | 16 | 50 | 38 | 1.316 |
| Camerún | Mandioca (nuevo jardín) | 3 | 20 | 19 | 1.053 |
| Camerún | Zona de barbecho (Chromolaena - 2 años) | 2 | 62 | 37 | 1.676 |
| Camerún | Zona de barbecho (Chromolaena - 4 años) | 6 | 28 | 22 | 1.273 |
| Kenia | Bosque secundario (caducifolio) | 1 | 35 | 57 | 0.614 |
| Madagascar | Sabana (Palma) | 2 | 26 | 26 | 1.000 |
S= riqueza de especies; Sg = riqueza de tipos funcionales; S/Sg = relación riqueza de especies y de tipos funcionales.
S = species richness; Sg =richness of functional types; S/Sg = richness relation of species and functional types.
Tipos funcionales de la clasificación de gillison
Gillison (2002, 2006) desarrollo una taxonomía de tipos funcionales. Los atributos funcionales evaluados constan de 36 caracteres, denominados elementos funcionales (Cuadro 2), por lo que las especies y morfoespecies con la misma combinación de elementos funcionales definen un tipo funcional (Gillison, 2002 y 2006).
Cuadro 2: Atributos y elementos funcionales del sistema de clasificación de Gillison (2006) para plantas vasculares.
Table 2: Attributes and functional elements of the Gillison (2006) classification system for vascular plants.
| Componentes del modelo funcional | Atributos funcionales | Elementos funcionales | Descripción |
| Cubierta fotosintética | Tamaño foliar | nr | Unidad foliar no repetida |
| pi | Picófila (< 0.2 cm) | ||
| le | Leptófila (0.2-0.8 cm) | ||
| na | Nanófila (0.8-2.5 cm) | ||
| mi | Micrófila (2.5-7.5 cm) | ||
| no | Notófila (7.5-12.5 cm) | ||
| me | Mesófila (12.5-25 cm) | ||
| pl | Platífila (25-36 cm) | ||
| ma | Macrófila (36- 83 cm) | ||
| mg | Megáfila (> 83 cm) | ||
| Inclinación foliar | ve | Vertical (> 30° por encima de la horizontal) | |
| la | Lateral (± 30° a la horizontal) | ||
| pe | Pendular (> 30° por debajo de la horizontal) | ||
| co | Compuesta | ||
| Clorotipo foliar | do | Dorsoventral | |
| is | Isobilateral o isocéntrico | ||
| de | Caducifolio | ||
| ct | Córtico (tallo fotosintético) | ||
| ac | Aclorófilo (sin clorofila) | ||
| ro | Roseta | ||
| so | Sólido tridimensional | ||
| Morfotipo foliar | su | Suculento | |
| pv | Nervadura paralela | ||
| fi | Filicoide (helechos) | ||
| ca | Insectívoro | ||
| Estructura vascular de soporte | Forma de vida | ph | Fanerofita |
| ch | Caméfita | ||
| hc | Hemicriptófita | ||
| cr | Criptófita | ||
| th | Terófita | ||
| li | Lianoide | ||
| Tipo de raíz | ad | Adventicia (e.g. contrafuertes) | |
| ae | Aérea (e.g. neumatóforos) | ||
| ep | Epifítica (e.g. Tillandsia spp.) | ||
| hy | Hidrofítica (e.g. Nymphaea spp.) | ||
| pa | Parasítica (e.g. Cuscuta spp.) |
Modelos empíricos de la relación riqueza - área
La relación S-A es dependiente del tipo de esquema de muestreo utilizados, entre los que se incluye el anidado, contiguo, no contiguo, islas, entre otros (Scheiner, 2003); por lo cual los resultados dependen del método de muestreo utilizado, por lo que se pueden obtener patrones diferentes, que son producto del método o artificios (Drakare, Lennon y Hillebrand, 2006; Dengler, 2008; Palmer, McGlinn y Fridley, 2008). De la discusión previa, los análisis presentados están fundamentados en muestreos anidados para establecer modelos de la relación S-A de los transectos considerados.
Entre los principales modelos de la relación S-A se encuentra el modelo potencial (Arrhenius, 1921 y 1923a):
Otro modelo es el logarítmico (Gleason, 1922 y 1925):
La relación S-Sg para el caso de los modelos potenciales para S-A y Sg-A, se puede estimar al despejar A de las relaciones (1) y (2) e igualarlas, definiendo:
con:
Para los modelos logarítmicos, despejándolos para ln(A) e igualándolos, se obtiene:
con:
El caso del modelo potencial para la relación S-A, y modelo logarítmico para la relación Sg-A, despejándolo para A e igualando los resultados se obtiene:
con:
Finalmente, con relación al modelo logarítmico de S-A y el modelo potencial para la relación Sg-A, despejando los modelos para A e igualándoles, se obtiene:
con:
Un modelo empírico propuesto para la relación S-Sg es un polinomio de segundo orden, con origen en (0, 0), que puede escribirse como:
Con origen en S = 1 y Sg = 1, bajo la consideración de S ≥ Sg.
Modelos de cinética de orden n de la relación riqueza - área. Los autores He y Legendre (1996) propusieron una cinética definida por:
Con f(S) = α+βS+γS2+ο(S3), donde el cuarto término representa el resto de la expansión de la serie de Taylor alrededor de S = 0 para f(S). El modelo logarítmico se genera de f(s) = α, el modelo potencial de f(S) = α + βS y el modelo de Archibald (1949) de f(S) = α + βS + γS2.
Un modelo alternativo propuesto para f(S) está dado por:
Cuya solución general, n ≠ 1, está dada por:
Para el caso n = 0, la solución de la relación (21) está dada por:
Que es el modelo logarítmico. Para el caso n = 1, la solución de la relación (21) está dada por:
Que es el modelo potencial.
El modelo de cinética de orden n de la relación (21) establece valores iniciales para S (S0) y A (A0).
Para el caso de la relación S-Sg, ambas con soluciones definidas por la relación (22) para S-A y Sg-A (parámetros S 0g , A 0g , k ng y n g ), se pueden despejar para ln(A), e igualar las ecuaciones para obtener:
con:
Ajustes estadísticos y métricas de incertidumbre
El modelo potencial generalmente se ajusta a los datos experimentales en función de sus logaritmos, por lo que la inversión al espacio aritmético introduce sesgos que son necesarios corregir (Baskerville, 1972; McGuinness, 1984). Para evitar estas correcciones, todos los ajustes estadísticos fueron realizados por regresión no lineal con el uso de la función SolverMR del software ExcelMR al minimizar el error de estimación.
Las métricas utilizadas de incertidumbre de los ajustes estadísticos, incluyeron el coeficiente de determinación (R2), la raíz del error cuadrático medio (RECM):
y el error relativo medio (ERM) y error absoluto medio (EAM) (en %):
donde
Resultados y Discusión
Modelos empíricos de la relación riqueza-área
Para el caso del modelo potencial, el ajuste de las relaciones S-A y Sg-A se muestra en la Figura 1, donde los ajustes estadísticos resultaron buenos (R2 > 0.98). En general, los ajustes para la relación S-A fueron mejores que para la relación Sg-A.
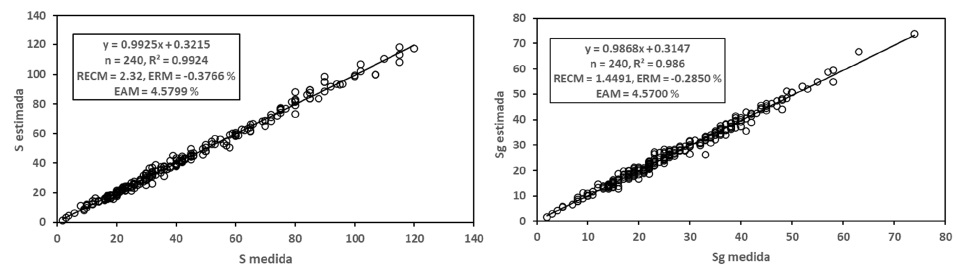
En la Figura 2 se muestran los resultados de los indicadores estadísticos de ajuste obtenidos en el caso del modelo logarítmico, los cuales fueron buenos (R2 > 0.98). En comparación, los resultados del modelo potencial resultaron en mejores ajustes que los del modelo logarítmico.
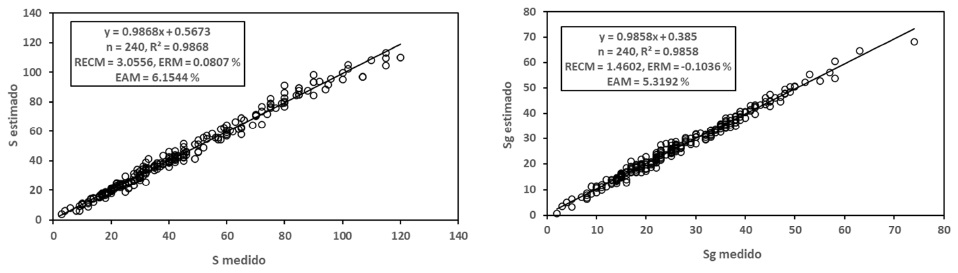
El uso del modelo de cinética de orden n presentó los mejores ajustes estadísticos (R2 > 0.99) y menores RECM respecto a los modelos potencial o logarítmico, tanto en el caso de la relación S-A, como la Sg-A, tal como se muestra en la Figura 3.
Relaciones s-sg
Las relaciones entre la riqueza de especies y la riqueza de tipos funcionales (S-Sg) se evaluaron usando los parámetros estimados de las relaciones S-A y Sg-A, como punto de partida para estimarlas, optimizando los parámetros del modelo S-Sg particular.
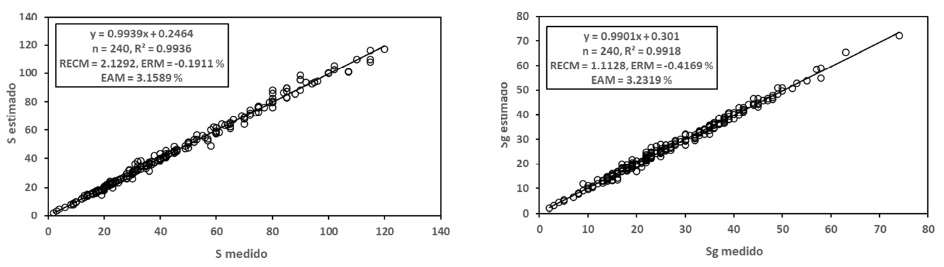
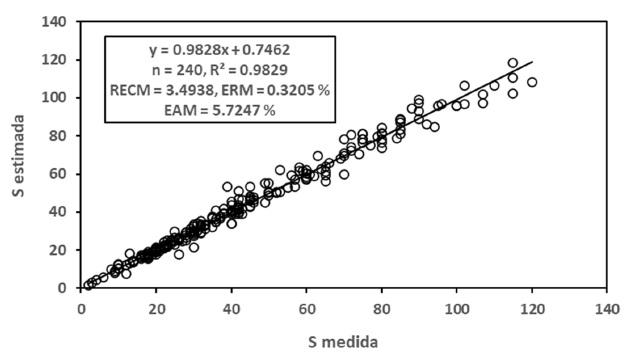
En la Figura 4 se muestran los resultados obtenidos para el modelo potencial, los cuales fueron buenos (R2 > 0.99). En la Figura 5 se presentan los resultados de los ajustes estadísticos para el modelo logarítmico, con resultados también buenos (R2 > 0.98). En general, los ajustes estadísticos del modelo potencial fueron mejores que los del modelo logarítmico.
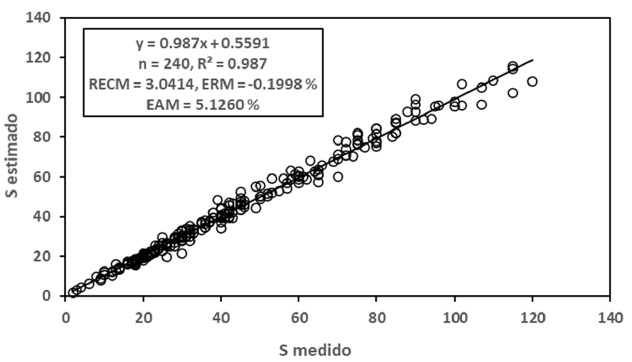
Para el caso de modelos diferentes de las relaciones S-A y Sg-A, en el Cuadro 3 se muestran los resultados obtenidos para estas relaciones, por lo que el modelo de la relación S-Sg se estimó usando los mejores modelos para las relaciones S-A y Sg-A.
Cuadro 3: Resultados de los ajustes estadísticos para las relaciones riqueza de especies-área (S-A) y riqueza de tipos funcionales-área (Sg-A).
Table 3: Results of the statistical adjustments for relationships of species richness-area (S-A) and richness of functional types-area (Sg-A).
| País | No. Transecto | Modelo Potencial | Modelo Logarítmico | Mejor Modelo | Modelo Potencial | Modelo Logarítmico | Mejor Modelo | ||||||||
| RECM | cg | zg | RECM | cg | zg | RECM | c | z | RECM | c | z | ||||
| India | 1 | 0.565 | 7.201 | 0.237 | 0.509 | -0.439 | 4.800 | Ln | 1.283 | 10.259 | 0.287 | 1.236 | -8.457 | 10.333 | Ln |
| India | 2 | 0.940 | 13.418 | 0.229 | 1.243 | 1.496 | 8.143 | Pot | 3.376 | 5.989 | 0.459 | 5.046 | -35.134 | 18.816 | Pot |
| India | 5 | 0.676 | 12.919 | 0.139 | 0.736 | 9.817 | 3.204 | Pot | 3.648 | 13.317 | 0.380 | 4.401 | -41.741 | 26.184 | Pot |
| India | 6 | 6.657 | 1.431 | 0.744 | 2.827 | -76.038 | 27.229 | Ln | 3.590 | 13.044 | 0.384 | 4.403 | -42.696 | 26.367 | Pot |
| India | 11 | 0.509 | 8.644 | 0.098 | 0.483 | 7.452 | 1.331 | Ln | 0.358 | 10.116 | 0.112 | 0.329 | 8.412 | 1.862 | Ln |
| India | 14 | 0.907 | 10.339 | 0.148 | 0.910 | 7.168 | 2.906 | Pot | 1.290 | 7.408 | 0.273 | 1.281 | -4.191 | 6.644 | Ln |
| Indonesia | 5 | 0.728 | 8.956 | 0.263 | 1.047 | -2.690 | 7.193 | Pot | 4.055 | 10.905 | 3.491 | 3.491 | -68.866 | 33.666 | Ln |
| Indonesia | 3 | 1.594 | 0.491 | 0.766 | 2.127 | -32.372 | 11.029 | Pot | 2.863 | 1.006 | 3.448 | 3.448 | -60.131 | 20.548 | Pot |
| Indonesia | 12 | 0.271 | 0.095 | 0.881 | 0.860 | -11.957 | 3.951 | Pot | 0.448 | 0.037 | 1.544 | 1.544 | -18.903 | 5.810 | Pot |
| Indonesia | 10 | 1.340 | 7.831 | 0.340 | 1.044 | -16.403 | 11.861 | Ln | 2.255 | 8.620 | 2.731 | 2.731 | -84.181 | 37.256 | Pot |
| Indonesia | 6 | 1.853 | 6.300 | 0.336 | 2.356 | -9.403 | 8.616 | Pot | 2.002 | 3.522 | 4.142 | 4.142 | -52.046 | 21.960 | Pot |
Ln = logarítmico; Pot = potencial; c = ordenada al origen del modelo riqueza de especies-área (S-A) cg = ordenda al origen de la relación riqueza de tipos funcionales-área (Sg-A); z = pendiente de la relación riqueza de especies-área (S-A) zg = pendiente del modelo riqueza de tipos funcionales-área (Sg-A); RECM=raíz del error cuadrático medio.
Ln = logarithm; Pot = potential; c = ordered to the origin of the model of species richness-area (S-A) cg = ordered to the origin of the relationship of functional types-area (Sg-A); z = slope of the relationship of species-area (S-A) zg = slope of the model of richness of functional types-area (Sg-A); RMSE = root mean square error.
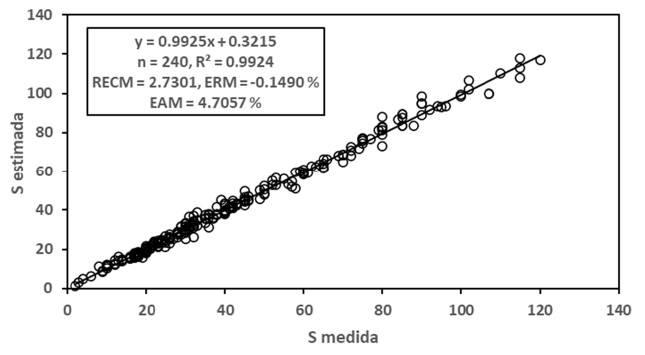
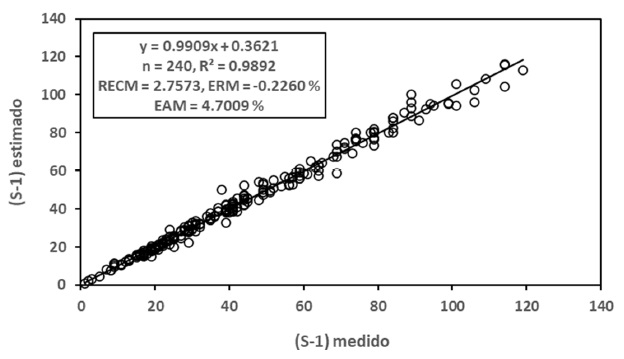
La Figura 6 muestra los resultados de los ajustes estadísticos para el caso de modelos iguales y diferentes de las relaciones S-A y Sg-A para la relación S-Sg, donde los resultados, aunque buenos (R2 > 0.98), no mejoran las estimaciones realizadas con modelos únicos.
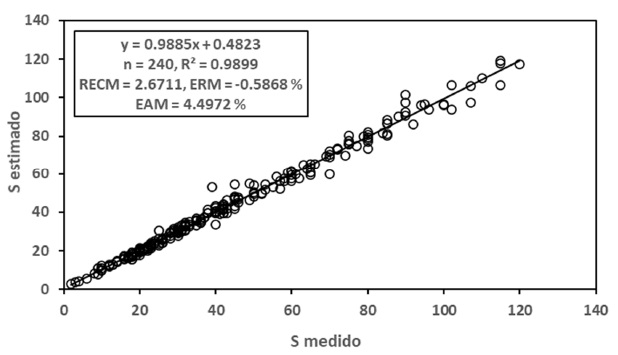
Para el caso del modelo polinómico de segundo grado, en la Figura 7 se presentan los resultados de los ajustes estadísticos, los cuales fueron buenos (R2 > 0.98).
Finalmente, el caso del modelo de cinética de orden n para las relaciones S-A y Sg-A, en la Figura 8 se muestran los resultados para la relación S-Sg, donde es evidente que los ajustes estadísticos fueron buenos (R2 ≈ 0.99).
Para validar teóricamente la relación riqueza de especies-área, principalmente del modelo potencial y logarítmico, se han propuesto diferentes hipótesis y teorías (Connor y McCoy, 1979; McGuinness, 1984; Turner y Tjørve, 2005), aunque no hay evidencia que soporte su validez generalizada (Connor y McCoy, 1979).
Los ajustes estadísticos de los modelos potencial y logarítmico presentan resultados satisfactorios, por lo que se pueden usar ambos modelos en forma confiable, además de que representan relaciones satisfactorias para S-Sg. El modelo potencial resulta en mejores ajustes generales.
El modelo de cinética de orden n que se introdujo superó a los modelos potencial, logarítmico y al del polinomio de segundo grado para la relación S-Sg, por lo que resultó ser el mejor modelo evaluado, por lo que su versatilidad de aplicación supera a los modelos potencial y logarítmico.
Conclusiones
El modelo de cinética de orden n resultó el mejor modelo en función del análisis estadístico, siendo consistente en tener los mejores ajustes en relación a los casos de los modelos potencial y logarítmico, al ser estos modelos casos particulares del modelo de cinética de orden n.
Los modelos potencial y logarítmico son viables para su aplicación, lo que se soporta con la evidencia de los ajustes estadísticos empíricos (R2 > 0.98) presentados en este trabajo, aunque no son los mejores, esto se basa en los ajustes estadísticos logrados con el modelo.
La cinética de orden n es un modelo flexible que puede ser utilizado en la gran mayoría de las aplicaciones, con ajustes estadísticos adecuados (R2 > 0.98), lo que permite un enfoque de modelo único de las relaciones riqueza - área y entre tipos de riqueza.

