











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
Understanding the role of environmental factors in driving soil microorganisms’ distributions is vital for sustainable crop production, particularly in vulnerable agricultural systems. The presence of arbuscular mycorrhizal fungi (AMF) is known to stimulate biological activity in the rhizosphere, a phenomenon commonly known as the “mycorrhizosphere effect” (Robert et al., 2012; Ferreira et al., 2020). This effect includes the attraction and selection of specific bacterial strains, such as plant growth promoting rhizobacteria, as well as the suppression of attack by pests and diseases (Tarnabi, Iranbakhsh, Mehregan and Ahmadvand, 2020). By meeting nutritional needs, AMFs more effectively develop the defense system of plants (Pieterse, Leon, Van der Ent and van Wees, 2009).
Highlighting the main edaphic and climatic factors disturbing soil microorganisms, especially AM fungi, helps to better understand how environmental change will influence soil microbial communities.
Currently, it has been estimated that more than 90% of terrestrial plants are associated with AMF belonging to the phylum Glomeromycota (Stürmer, Bever and Morton, 2018). These mutualistic associations benefit a wide range of vascular flowering plants that develop in different ecosystems, especially those where phosphorus availability is limited. The objective of this research work was to isolate, identify and evaluate native AMF of the state of Chiapas in peanuts (Arachis hypogaea) as a crop of economic importance for the municipality of Suchiapa, Chiapas, Mexico.
Materials and Methods
The experiments were established in plots of the “El Capricho Divino” ranch in the municipality of Suchiapa, Chiapas, Mexico. Located at 16° 37’ N and 93° 60’ W, which corresponds to tropical regions, with more than 900 to 1000 mm of annual precipitation (INAFED, 2022), to evaluate the tritrophic interaction and the effect of AMF on important crops economical.
Obtaining native inoculums
The native inoculums were obtained from virgin rhizospheric soil of a tropical forest located at 16° 37’ N and 93° 60’ W, with more than 900 to 1000 mm of annual precipitation, corresponding to the municipality of Suchiapa, Chiapas, Mexico.
Soil samples were obtained at a depth of 20 cm, using a stainless-steel soil sampler tube (3 cm internal diameter). The samples were sieved (<2 mm) and stored at -20 °C for the extraction, physical-chemical (NOM-021-RECNAT-2000, 2002), and molecular evaluation of the present spores (Lui et al., 2016).
Structural and molecular characterization of spores from native inoculum
The characterization of the inoculum allowed obtaining the number of AMF spores g-1 from rhizospheric soil. The spores are removed and isolated according to Gerdemann and Nicolson (1963). Subsequently, the assembly was carried out in polyvinyl lactoglycerol (PVLG) for identification under the microscope binocular Zeiss Primo Star (Schenk and Pérez, 1990; Méndez, Marmolejo, Olalde, Cantú and Varela, 2012; Pérez-Luna, Alvarez, Hernández and Sánchez, 2016).
For molecular identification, the polymerase chain reaction denaturing gradient gel electrophoresis (PCR-DGGE) was used to analyze the AMF community. AMF-specific primers were designed based on the most variable part of the small subunit (SSU) rRNA gene region. Nested PCR was then conducted; the amplification primers were performed as indicated by Guo et al. (2020).
Evaluation of agronomic parameters
Seeds of the crop of interest (peanut) were sterilized by the solarization technique, with exposure to 45 °C for 24 h. Subsequently, the seeds were sown in the open field, in quadrants of 2×4 m with 30 to 40 cm between rows, placing 2 seeds per row, at the same time for a treatment 10 g of the substrate was inoculated with native AMF and for other treatments 2 g of commercial AMF, allowing a total of 70 to 100 spores to be inoculated, according to Karagiannidis, Thomidis and Panou-Filotheou (2012).
The experiment was established using a completely randomized design. Six treatments were developed (T1) Control plants without AMF; (T2) Plants with vermicompost leachate; (T3) Plants inoculated with commercial mycorrhizae; (T4) Plants inoculated with native mycorrhizae; (T5) Plants inoculated with commercial mycorrhizae + leachate; (T6) Plants inoculated with commercial mycorrhizae + native mycorrhizae. Each treatment was evaluated in triplicate with a total of 20 replications. Statistical analysis was performed using an ANOVA with the analysis of means using Tukey’s test (P < 0.05), establishing a confidence interval of 95%; Allowing us to assume that the evaluated variables are normally distributed. Using the statgraphics centurion software version 15.2.06 (Statgraphics Technologies, 2005; Tribouillois et al., 2016).
During the development of the plant, every 15 days the germination percentage, agronomic characteristics such as plant height, biomass width, leaf damage by Cercospora personata, and flower number were determined, at the end of the experiment the index of leaf area, root length, and fruit yield were determined (Akhter, Hage-Ahmed, Soja and Steinkellner, 2015). Likewise, the quality of the fruits was determined according to CODEX STAN 200-1995.
Peanut lipid extraction
At the end of the harvest, the peanuts were dried at 55 °C for 48 h, later the lipid extraction was carried out through the soxhlet method according to O’Fallon, Busboom, Nelson and Gaskins (2007). The samples were evaluated through gas chromatography coupled to a mass spectrometer (GC/MS), for this purpose 100 mg L-1 of oleic acid (C18) methyl ester (Sigma Aldrich) was used as a positive control according to the conditions established by Rhea et al. (2012).
Results and Discussion
The planting of peanuts was carried out at the “El Capricho Divino” ranch in the municipality of Suchiapa, Chiapas, in an area of 144 m2, giving rise to 18 quadrants with dimensions of 2×4 m each quadrant, where 6 treatments were evaluated in triplicate, the random distribution of the treatments is observed in Figure 1.

Figure 1: Random distribution of six treatments evaluated in the field on peanut crops. T1 = control; T2 = leached; T3 = commercial mycorrhiza; T4 = native mycorrhiza; T5 = leachate + commercial mycorrhiza; T6: commercial + native mycorrhiza.
The peanut plants developed vegetatively, and some physical changes were observed in the different established treatments; in the case of the control treatment, it was affected by the presence of the Cercospora personata pest, unlike the treatments evaluated with vermicompost leachate and mycorrhizae that presented better development and fruit yield.
Obtaining and characterizing the spores of the native inoculum
For the characterization of the native inoculum, 10 g of soil was analyzed, of which 55 to 70 spores were identified on average and the genera correspond to Rhizoglomus that presented solitary spores circular in shape, light yellow to amber in color and in some presence of hypha was observed and the genus Acaulospora was observed from a sporiferous saccule (mature spore) without supporting hyphae. Therefore, of the analyzed sample, 67% corresponds to the genus Acaulospora (Figure 2).

Figure 2: Morphological description of the spores identified in the native inoculum corresponding to the genus Rhizoglomus and Acaulospora.
According to this identification Birhane, Aregawi and Giday (2017) demonstrated a high density of spores in forest soils corresponding to the genera Rhizoglomus and Acaulospora, these being responsible for the restoration and restructuring of ecosystems. Likewise, these previously identified spores were evaluated by Ortas (2015), who mentions that these genera have a significant impact on the growth and foliar development of the inoculated plants, allowing an excellent root development that benefits the selection of nutritive substances from the soil for the formation of fruits (Buba and Muhammad, 2020). Rhizoglomus is also said to have a wide range of adaptations in terms of soil type and edaphoclimatic conditions. On the other hand, Acaulospora spp., find an ecological niche in soils of organic systems, so Rhizoglomus species are the first to sporulate, while Acaulospora species do so after the winter period, therefore, they are generally dominant species in agricultural soils (Oehl et al., 2004), therefore in peanut the use of mycorrhizae could contribute to the increase in yields and the reduction or substitution of chemical and that damage the environment by contaminating soil, water, and food with the consequent problems for human health.
Evaluation of agronomic parameters
After sowing peanuts, the germination percentage was evaluated, identifying the best treatment with 98% for the leachate + commercial mycorrhiza treatment, unlike the control treatment which showed a germination percentage of 78% (Figure 3).
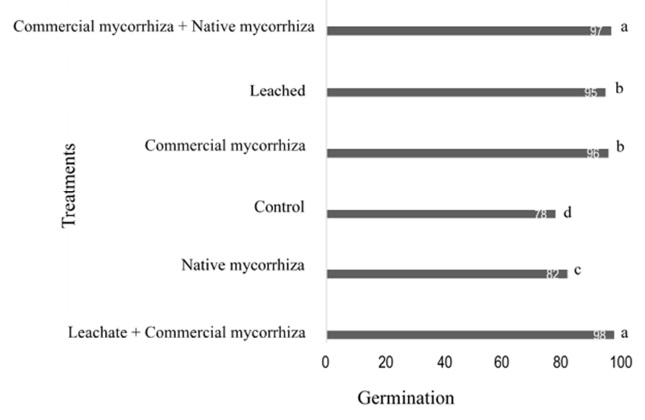
Figure 3: Evaluation of the germination percentage after 15 days of peanut sowing under the effect of six treatments. T1 = control; T2 = leached; T3 = commercial mycorrhiza; T4 = native mycorrhiza; T5 = leachate + commercial mycorrhiza; T6 = commercial + native mycorrhiza. Mean of three repetitions. Each column without a common superscript differs significantly at P <0.05.
Gazey, Abbott and Robson (1993) demonstrated that AMFs can overcome the dormant state of seeds and accelerate germination processes (Swangmaneecharern, Serivichyaswat and Nontachaiyapoom, 2012; Alghamdi, 2019). This promotion of germination by plants previously inoculated with mycorrhizal fungi is due to the increase in the synthesis of the plant hormone strigolactones, thus also, it allows high activity in the regulation of plant architecture (Kountche et al., 2018).
Pre-harvest evaluation
In the experimental area, agronomic evaluations were carried out every 15 days, before, during, and after the harvest. Before the peanut harvest, the plant height, biomass width, and the number of flowers were evaluated (Table 1).
Table 1: Evaluation of agronomic variables under the effect of vermicompost leachate and mycorrhizae on peanut crops.
| Days | Leachate + Commercial mycorrhiza | Native mycorrhiza | Control | Commercial mycorrhiza | Leachate | Commercial mycorrhiza + Native mycorrhiza | ||||||||||||
| 15G (%) | 98 + 0.5a | 82 + 0.7c | 78 + 0.1d | 96 + 0.1b | 95 + 0.3b | 97 + 0.6a | ||||||||||||
| A.V | Ha | Wa | Fb | Hb | Wb | Fa | Hc | Wc | Fb | Ha | Wa | Fc | Hb | Wb | Fb | Hb | Wb | Fb |
| - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - cm - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - | ||||||||||||||||||
| 30 | 17.7* + 0.6 | 30 + 0.9 | 0 + 0.1 | 13 + 0.2 | 25 + 0.4 | 0 + 0.0 | 11 + 0.5 | 20 + 0.3 | 0 + 0.1 | 16 + 0.2 | 30 + 0.3 | 0 + 0.1 | 12.3 + 0.1 | 24.6 + 0.2 | 0 + 0.02 | 12.7 + 0.9 | 24 + 0.1 | 0 + 0.4 |
| 45 | 22.4 + 0.4 | 33 + 0.4 | 1.7 + 0.6 | 20.6 + 0.4 | 28.1 + 0.2 | 1.7 + 0.2 | 20 + 0.7 | 31.8 + 0.4 | 1.68 + 0.2 | 20 + 0.4 | 33 + 0.5 | 1.69 + 0.3 | 20 + 0.9 | 30.6 + 0.3 | 1.6 + 0.4 | 21.6 + 0.1 | 29.2 + 0.5 | 1.2 + 0.3 |
| 60 | 25.8 + 0.5** | 47 + 0.2 | 4.3 + 0.2 | 25.3 + 0.7 | 31.3 + 0.1 | 7.9 + 0.1 | 23.3 + 0.6 | 39.6 + 0.8 | 4.53 + 0.4 | 23 + 0.1 | 36 + 0.9 | 3.2 + 0.4 | 23.2 + 0.8 | 38.3 + 0.1 | 3.9 + 0.9 | 23.4 + 0.6 | 41.7 + 0.8 | 3.8 + 0.6 |
| 75 | 30.2 + 0.3 | 59 + 0.6 | 35 + 0.7 | 27.6 + 0.9 | 45.7 + 0.3 | 31 + 0.2 | 28.1 + 0.1 | 53 + 0.9 | 29.7 + 0.7 | 30 + 0.6 | 45 + 0.7 | 31.7 + 0.7 | 27.2 + 0.9 | 43.7 + 0.5 | 31.1 + 0.9 | 29.9 + 0.2 | 62 + 0.7 | 34.2+ 0.2 |
| 90 | 36.3 + 0.9 | 63 + 0.7 | 13 + 0.6 | 35.3 + 0.6 | 65.0 + 0.2 | 1.3 + 0.6 | 35.5 + 0.1 | 57.4 + 0.5 | 6.56 + 0.5 | 34 + 0.5 | 58 + 0.8 | 1.5 + 0.9 | 30.5 + 0.7 | 60.5 + 0.7 | 1.5 + 0.3 | 37.3 + 0.2 | 70.1 + 0.4 | 9.8 + 0.2 |
| 105 | 45.9 + 0.7 | 74 + 0.9 | 0 + 0.1 | 40.4 + 0.4 | 67.8 + 0.1 | 0.2 + 0.4 | 38.1 + 0.4 | 60.3 + 0.3 | 0 + 06 | 41 + 0.2 | 60 + 0.4 | 0 + 0.7 | 40.36 + 0.9 | 70.0 + 0.1 | 0.03 + 0.2 | 45.4 + 0.1 | 79.6 + 0.3 | 0.06 + 0.1 |
A.V= agronomic variables; G = germination; H = high; W = width; F = flower. * Mean of three repetitions. ** The averages (± standard error) within each column without a common superscript differ significantly at P <0.05.
In table 1, it is observed that the highest values of the variables: plant height and biomass width were reached at 105 days in the leachate + commercial mycorrhiza and commercial mycorrhiza + native mycorrhiza treatments, without showing a significant statistical difference. This clearly suggests the effect of mycorrhizae and organic amendments on plant vigor and resistance to pest attack and disease development.
These results were observed in other crops (Cofré, Becerra, Marro, Domínguez and Urcelay, 2020) since the functional link between aerial systems and underground microorganisms in a plant system determines the success of restoration and reestablishment of degraded ecosystems; in this sense, the use of commercial or selected mycorrhizae and native, autochthonous or indigenous mycorrhizae promote the development of the plant, so it was shown that although autochthonous mycorrhizae contribute to plant growth, however, the participation of mycorrhizae selected is much higher (Birhane et al., 2017).
It is important to highlight the benefits of Arbuscular Mycorrhizal Fungi (AMF) since it has been shown that they are promoters of beneficial interactions between plants and growth-promoting bacteria (PGPB), as well as help to improve the absorption of nutrients by the host plant and they alter their physiology (controlling plant hormone levels) to resist abiotic factors and external pathogens, significantly affecting the quality of the crop (Emmanuel and Babalola, 2020).
On the other hand, it was observed that high flower production was obtained 75 days after sowing, in the leachate + commercial mycorrhiza treatments and the commercial mycorrhiza + native mycorrhiza treatment. The results coincide with that reported by Jami, Rahimi, Naghizadeh and Sedaghati (2020), who showed that the vermicompost and mycorrhiza association improve the assimilation and availability of nutrients at the rhizosphere level, promoting their maximum uptake to accelerate growth and flowering for fruiting.
Likewise, a wide relationship between the number of flowers and root elongation stimulated by the symbiotic association with AMF has been demonstrated, said the association is catalyzed by the calcium and calmodulin-dependent kinase enzyme (CCaMK), therefore, an increase in the number of flowers, it is synonymous with the increase in kinase production, guaranteeing an extraordinary symbiotic association between roots and AMF (Nouri and Reinhardt, 2015).
According to the constant evaluation (every 15 days) of agronomic variables, the following kinetics of the behavior of plant height, biomass width, and the number of flowers are observed (Figure 4).

Figure 4: Flowering kinetics, height and width of the development of peanut crops in a period of 105 days under the influence of vermicompost leachate and mycorrhizae. Mean of three repetitions.
In the previous graphs (Figure 4), the commercial leachate + mycorrhiza is observed as the best treatment, as a result of the tripartite interaction (Plant, AMF, and leachate), this triple association allowed the greater production of foliar biomass and flowers, due to the interaction synergistic that promotes the production of extra-radical hyphae to improve the uptake of micro and macronutrients responsible for the metabolic and biochemical processes of a plant (Xu et al., 2020).
On the other hand, the incidence of Cercospora personata, commonly referred to as leaf spot, considered the disease with the greatest affection in peanut cultivation was evaluated (Figure 5).

Figure 5: Image analysis of the damaged area in the leaves of the foliar biomass of the peanut crop under the influence of vermicompost leachate and mycorrhizae. * Mean of three repetitions. ** The averages (± standard error) within each column without common superscript differ significantly at P <0.05.
In figure 5, it is observed that the control treatment without the incidence of AMF presented the area with the highest damaged surface (41.5%) and the leached treatment + commercial mycorrhiza showed the lowest percentage (1.6%) of surface damage by Cercospora personata.
The results demonstrate the beneficial effect of the interaction between AMF and leachate against pathogenic fungi such as Cercospora personata, in this sense, Eke et al. (2020)) corroborate that AMF inoculation changed the biochemical processes of the bean plant (Phaseolus vulgaris L.) with a subsequent impact on growth and greater suppression of root rot by Fusarium under greenhouse conditions.
So too, El-Sharkawy, Rashad and Ibrahim (2018) showed that the use of mycorrhizae significantly reduces the disease caused by Trichoderma spp. in wheat due to the increase in the production of peroxidase and polyphenol oxidase enzymes, increasing the total phenol content and improving the growth and performance parameters tested. Among the reported underlying mechanisms of AM interaction in plant resistance against pathogens are the specific competition against pathogens, root alterations, and other defense related responses (Wu et al., 2013).
The results present AMF as antagonists to be implemented in the agricultural field as a biological control (Karliński, 2021).
On the other hand, the Leaf Area Index (IAF) was determined at 105 days of growth of the peanut plants in the different treatments evaluated (Figure 6).

Figure 6: Analysis of the Leaf Area Index (IAF) of the foliar biomass of the peanut crop under the influence of vermicompost leachate and mycorrhizae. * Mean of three repetitions. ** The averages (± standard error) within each column without a common superscript differ significantly at P < 0.05.
Figure 6 shows that the treatment with the highest Leaf Area Index were the plants treated with the mixture of commercial and native mycorrhizae (4.01) and the treatments with the lowest IAF (2.50) were the plants without receiving treatments (control).
In this sense, the mycorrhizae inoculated in the rhizosphere allow a balance between carbon and nitrogen, as well as adequate humidity values, which positively impact the production of foliar biomass (Karliński, 2021; Netherway, Bengtsson, Krab and Bahram, 2020).
Evaluation during harvest
During the harvest, the length of the roots of the peanut plants of the different treatments was evaluated (Figure 7).

Figure 7: Evaluation of the physical characteristics of the root system of peanut plants under the effect of vermicompost leachate and mycorrhizae. length; width. * Mean of three repetitions. ** Each column without a common superscript differs significantly at P < 0.05.
In figure 7, the incidence of mycorrhizae in peanut plants is observed, so that these treatments showed elongation of pivoting roots, however, the treatment that generated the greatest extension of the central root, were the plants treated with leachate. + Commercial mycorrhiza.
As demonstrated by Gazey, Abbott and Robson (1992), a critical length of the mycorrhizal root is associated with the initiation of sporulation of these fungi, since the length of the colonized root and the number of spores produced are well correlated in the case of the Rhizoglomus and Acaulospora genera.
Post-harvest evaluation
After the harvest, the number of peanuts per plant was analyzed in the first instance with respect to each treatment (Figure 8), in which, it was determined that the highest number of peanuts (31) was reached in the plants treated with vermicompost leachate + Commercial mycorrhizae, as well as the plants treated with commercial mycorrhizae + native mycorrhizae, these treatments did not show significant statistical differences.

Figure 8: Analysis of peanut yield per plant under the incidence of vermicompost leachate and mycorrhizae. Mean of three repetitions. Each column without a common superscript differs significantly at P < 0.05.
On the other hand, the yield of peanut production was evaluated, in Figure 9 the highest production in the plants was observed under the effect of commercial and native mycorrhizae (10.5 kg per 8 m2).
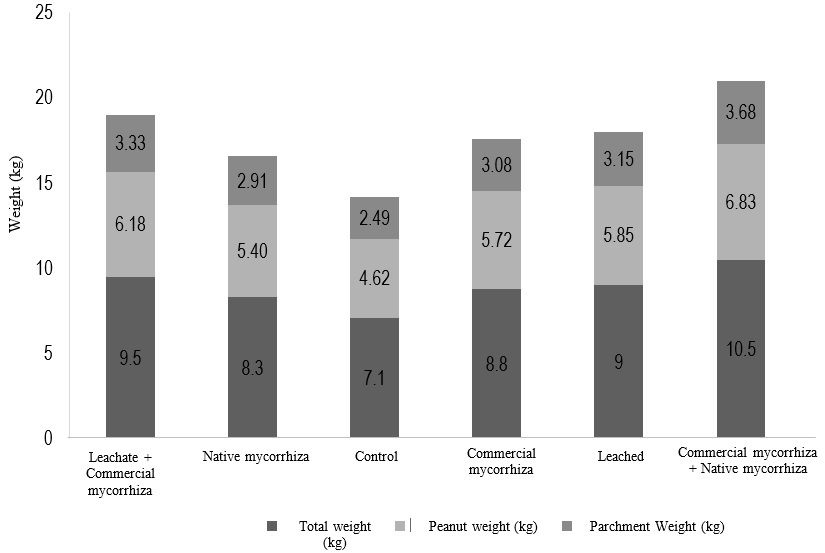
Figure 9: Analysis of the effect of vermicompost leachate and mycorrhizae on the relationship between fruit weight and parchment weight. Mean of three repetitions.
The effects of AMF in favor of the increase in fruit yield are due to the fact that they allow the greater absorption of P and K destined for the metabolism of the formation of the fruits, thus also, they benefit the increase of the activity of superoxide dismutase (SOD), catalase (CAT), peroxidase (POD) and ascorbate peroxidase (APX) reducing oxidative damage, benefiting the increase in the production of quality fruits (Latef and Chaoxing, 2011).
The following evaluations were carried out based on the quality of the peanut, during this evaluation it was identified that the largest peanuts correspond the dimensions (Length; Diameter) of 4.32; 1.63 cm, and 4.34; 1.47 cm, for the plants treated with leachate and leachate + commercial mycorrhiza, respectively (Table 2).
Table 2: Evaluation of physical variables for peanut quality according to CODEX STAN 200-1995.
| Treatments | Structure | Length | Diameter | Quality |
| - - - - - - - - - - cm - - - - - - - - | ||||
| Leached | Exocarpus | 4.32* + 0.12** | 1.63 + 0.21 | Ba |
| Endocarpus | 1.86 + 0.18 | 0.85 + 0.16 | ||
| Control | Exocarpus | 3.59 + 0.23 | 1.42 + 0.43 | Cd |
| Endocarpus | 1.32 + 0.24 | 0.85 + 0.23 | ||
| Native mycorrhizae | Exocarpus | 4.11 + 0.19 | 1.43 + 0.12 | Bc |
| Endocarpus | 1.89 + 0.12 | 0.85 + 0.29 | ||
| Commercial mycorrhiza + native mycorrhiza | Exocarpus | 4.22 + 0.11 | 1.45 + 0.16 | Bb |
| Endocarpus | 1.91 + 0.18 | 0.88 + 0.26 | ||
| Commercial mycorrhiza | Exocarpus | 4.10 + 0.10 | 1.38 + 0.27 | Bc |
| Endocarpus | 1.94 + 0.11 | 0.86 + 0.31 | ||
| Leached + Commercial mycorrhiza | Exocarpus | 4.34 + 0.09 | 1.47 + 0.19 | Ba |
| Endocarpus | 1.87 + 0.14 | 0.84 + 0.11 | ||
* Mean of three repetitions. ** The averages (± standard error) within each column without a common superscript differ significantly at P <0.05.
This is due to the fact that biofertilizers have been shown to increase the chlorophyll content of the leaves, which is related to a higher rate of photosynthesis, as well as the average daily CO2 assimilation rate during the crop production cycle, increasing the yield in fruit production (Zhang et al., 2020).
Regarding the quality of peanuts, according to the criteria of CODEX STAN 200-1995, extra quality peanuts correspond to a length between 4.5 and 5 cm (Category A), peanuts that could be exported. On the other hand, the peanuts under the treatments implemented in this research work correspond to the basic quality with dimensions of 4 to 4.5 cm (Category B), peanuts that can be marketed in the country destined for agro-industrial transformation.
Likewise, the content of total lipids was determined under the effect of the different treatments, according to the above, the highest concentrations of lipids are found in the plants treated with vermicompost leachate (Figure 10).

Figure 10: Analysis of the effect of vermicompost leachate and mycorrhizal on total lipid yield of the peanut fruits.
Figure 10 shows that the plants treated with leachate allowed lipid yields of up to 59%, in this sense Girardi, Passone, García, Nesci and Etcheverry (2018) mentioned that peanuts are an oilseed with a high concentration of lipids of 40 to 75% depending on the extraction method, the solvent extraction method is the best technique for obtaining lipids from peanuts, in this sense, peanuts under the effect of the treatments evaluated in this research work reached concentrations of total lipids higher than 45% (Russo and Webber III, 2012; Li et al., 2017).
Figure 11 shows the average fatty acid composition of peanut oil. Unsaturated fatty acids represented the largest fraction, the fatty acid that was identified in the highest percentage was oleic acid at 52% and linoleic at 21%, as for saturated fatty acids, palmitic acid was identified with a 20%, These percentage data were identified in treatment 5 which consisted of the application of leachate + commercial mycorrhizae on the peanut’s plants (Table 3).

Figure 11: Chromatogram of the free fatty acid profile of peanuts under the effect of vermicompost leachate.
Table 3: Abundance of the profile of fatty acids present in peanut oil under the incidence of vermicompost leachate.
| N | Compounds | Chemical formula | Parent Ion | RT1 | Treatment Abundance percentage | Fatty acid | |||||
| (m/z)2 | (min) | - - - - - - - - - - - - - - % - - - - - - - - - - - - - | |||||||||
| T1 | T2 | T3 | T4 | T5 | T6 | ||||||
| 1 |

|
C18H34O2 | 282.47* | 16.50 | 41d** | 49b | 49.5b | 47.7c | 52a | 51a | Monounsaturated |
| 2 |
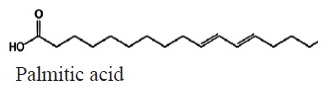
|
C18H32O2 | 280.44 | 18.00 | 30a | 21b | 20c | 22b | 21b | 20c | Saturated |
| 3 |
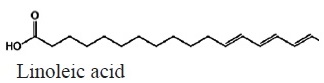
|
C18H32O2 | 280.44 | 14.60 | 20a | 21a | 20.5a | 19.5b | 20a | 19.2b | Polyunsaturated |
| 4 |
|
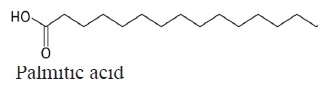
C18H36O2 | 284.48 | 11.20 | 5ª | 5a | 5.5a | 6.3a | 5a | 6.5a | Saturated |
| 5 |
|
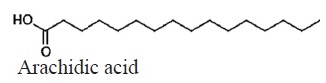
C20H40O2 | 312.53 | 19.00 | 1.3a | 0.7b | 2a | 1.7a | 1a | 1.5a | Saturated |
| 6 |
 |
C14H28O2 | 228.37 | 12.80 | 1.5a | 1.3a | 1.8a | 2a | 0.7b | 1.3a | Saturated |
| 7 |

|
C22H44O2 | 340.58 | 17.30 | 1.2a | 2a | 0.7b | 0.8b | 0.3b | 0.5b | Saturated |
T1 = control; T2 = leached; T3 = commercial mycorrhiza; T4 = native mycorrhiza; T5 = leachate + commercial mycorrhiza; T6 = commercial + native mycorrhiza; RT = retention time. Parent ion (m/z) = molecular ions of the standard compounds (mass to charge ratio). * Mean of three repetitions. ** Each column without a common superscript differs significantly at P < 0.05.
Conclusions
The establishment of the symbiosis between the fungus and the plant leads to a sequence of recognition stages causing both morphological and physiological changes in the two interacting organisms. Therefore, in the present research work, it was demonstrated that the best treatments were the plants inoculated with vermicompost leachate + commercial mycorrhizae and the commercial mycorrhiza treatment + native mycorrhizae with a leaf area index of 3.79 and 4.01 respectively, however, the treatment that achieved the best yields in terms of peanut production was the mixture of commercial mycorrhizae + native mycorrhizae with 10.5 kg in an area of 8 m2, the native mycorrhizae isolated in the present investigation were made up of the genera Rhizoglomus and Acaulospora (Rhizoglomus clarum, Acaulospora alpina, and Acaulospora aff. bireticulata), on the other hand, it was shown that mycorrhizae significantly decreased (+ 40%) the pathogenic effect generated by Cercospora personata in peanut plants. Finally, the highest concentration of lipids was obtained in peanuts under the effect of vermicompost leachate. The scientific contributions of this work will significantly impact the peanut value production chain, of interest to the agribusiness and pharmaceutical industry of Mexico.

