











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
El ñame (Dioscorea sp.) es un cultivo de importancia en América tropical, debido a su contribución en la seguridad alimentaria de las poblaciones rurales. En Colombia se siembra principalmente en la región Caribe, donde ha alcanzado importancia social y económica debido a que aproximadamente 20.000 familias, principalmente de pequeños y medianos productores, derivan su sustento diario de este cultivo (Narváez et al., 2019; PNUD, 2019). En esta región, se siembran principalmente dos especies de ñame: el ñame “Criollo” (Dioscorea alata) y el ñame “Espino”, (Dioscorea rotundata), siendo este último el preferido debido a que presenta un mercado más amplio y además su calidad es apta para el mercado de exportación.
El ñame espino es originario del África, pero actualmente se encuentra ampliamente distribuido por las zonas tropicales y subtropicales del mundo, debido a que las plantas de esta especie forman tubérculos comestibles de alto valor calórico (Padhan y Panda, 2020). Los tubérculos formados son de variados tamaños, con coloración externa castaño y pulpa de color blanco.
Uno de los inconvenientes al que se enfrentan los agricultores para la siembra de este cultivo es el largo periodo de dormancia que presentan los tubérculos después de la cosecha (Darkwa et al., 2019). Esto obliga a almacenar los tubérculos cosechados durante largos períodos de tiempo (Swannell et al., 2003; Kone-Coulibaly et al., 2018), incluso si las condiciones del suelo para la siembra son adecuadas. Este almacenamiento puede durar entre 1.5 a 4 meses, y se realiza bajo condiciones de sombra, en lugares frescos y aireados, en la finca del productor. Sin embargo, durante este periodo de almacenamiento los tubérculos se exponen al ataque de plagas y enfermedades que ocasionan pérdidas de hasta 50% de la producción en almacenamiento (Sánchez et al., 2020; Amadioha et al., 2001; Aighewi et al., 2015). En la práctica, los agricultores almacenan los tubérculos que van a ser usados como semilla para la siembra, y cuando observan el inicio natural del rebrote, realizan las siembras. En Colombia, como en muchas otras partes del mundo, la siembra de los cultivos se realiza utilizando tubérculos enteros o porciones de estos, denominados comúnmente “cabeza”, “medio” y “cola” (Brice et al., 2019). Sin embargo, la brotación de los tubérculos no es uniforme, se presentan variaciones en la capacidad de las distintas yemas, lo que ocasiona desuniformidad en los cultivos, pérdida de población y bajo rendimiento de campo.
Si bien, se sabe mucho sobre el control de la dormancia en otras especies, en general en el cultivo del ñame el conocimiento del mecanismo de dormancia del tubérculo, y la forma de controlarlo han sido poco estudiados. Diferentes estudios se han realizado, en otros países, para tratar de romper la dormancia e inducir la brotación del tubérculo de ñame, con resultados variables debido a los efectos del cultivar, la madurez de los tubérculos, las concentraciones, los productos y la limitación en la penetración de estos en el tejido vegetal. Entre los productos que se reportan con mayor actividad en el rompimiento de la dormancia y el estímulo de la brotación se encuentran los reguladores del crecimiento vegetal (RCV) como la thiourea, el cloruro de mepiquat y el etefón (Cibes y Adsuar 1966; Hamadina y Craufurd, 2015). La presente investigación se llevó a cabo con el objeto de evaluar la efectividad de los RCV para la ruptura de la dormancia, usando secciones o porciones de tubérculos para facilitar la penetración de los productos, en lugar de tubérculo enteros, con diferente grado de desarrollo o madurez, en dos variedades de ñame espino.
Materiales y Métodos
Producción de tubérculos para el ensayo
Con el objeto de obtener tubérculos de diferentes edades o estados de desarrollo para evaluar la brotación, se realizaron siembras en campo de dos variedades de ñame 0307-49SB y 0307-50CB. Las plántulas se obtuvieron a partir de la siembra de secciones de tubérculo de 50 g de peso, obtenidas de las regiones apical, media y basal, las cuales fueron desinfectadas en solución acuosa con base de Mancozeb 80WP (800 g kg-1 de Mancozeb), en dosis de 1.0 g L-1 y oxicloruro de cobre 58.8WP (588 g kg-1 de oxicloruro de cobre), en dosis de 2.0 g L-1 por 10 min. Las secciones de tubérculo fueron sembradas en bandejas de germinación con sustrato conformado por arena, aluvión y cascarilla de arroz en proporción 1:3:1. Este sustrato fue previamente desinfectado empleando solución de West Terrasafe® (2.0 mL L-1 1.3 Diformil Propano) y Fipronil (1.0 mL L-1 de Fipronil). Las plántulas se dejaron crecer en casa de malla por espacio de 60 días, a temperatura ambiente con promedio de 30±2 °C y humedad relativa de 80±2%; posteriormente se seleccionaron plantas uniformes para su siembra en campo.
La siembra en campo se realizó en un área de terreno 800 m2, en una finca localizada en la vereda El Llano, municipio de Ciénaga de Oro, Córdoba, ubicada geográficamente a 8° 47’ 20” N y 75° 35’ 20.1” O, con 163 m de altitud. Las condiciones ambientales durante el desarrollo del cultivo en el sitio experimental fueron: temperatura media de 27.8 °C, humedad relativa de 85% y precipitación de 1350 mm. Las plántulas de ñame se sembraron al inicio de la temporada de lluvias, en el mes de abril, con distancia de 1.0 m entre surcos y 1.0 m entre plantas, para una densidad de población de 10 000 plantas por hectárea (pl ha-1). Las características fisicoquímicas del suelo en el sitio experimental se muestran en el Cuadro 1.
Cuadro 1: Propiedades fisicoquímicas del suelo en el sitio experimental de Vereda Los Llanos, Ciénaga de Oro, Córdoba, Colombia.
Table 1: Physicochemical properties of soil at the experimental site Vereda Los Llanos, Cienega de Oro, Córdoba, Colombia.
Parámetro |
Unidad |
Valor |
Textura |
||
pH |
6.8 |
|
Conductividad eléctrica |
dS m-1 |
0.14 |
Materia orgánica (MO) |
g 100 g-1 |
1.29 |
Fósforo disponible (P) Bray II |
mg kg-1 |
≤ 3.87 |
Acidez intercambiable (Al+H) |
g 100 g-1 |
ND |
Aluminio intercambiable (Al+H) |
cmol (+) kg-1 |
ND |
Calcio intercambiable (Ca) |
cmol (+) kg-1 |
3.09 |
Magnesio intercambiable (Mg) |
cmol (+) kg-1 |
0.94 |
Potasio intercambiable (K) |
cmol (+) kg-1 |
0.12 |
Sodio intercambiable (Na) |
cmol (+) kg-1 |
≤ 0.14 |
Capacidad intercambio catiónico (CICE) |
cmol (+) kg-1 |
4.29 |
† FA = franco arcillosa.
† FA = clay loam.
El cultivo de ñame se fertilizó con base en el análisis de suelos y los requerimientos del cultivo (O'Sullivan, 2010), aplicando una mezcla de 60.7 kg ha-1 de N, 24 kg ha-1 de P y 24 kg ha-1 de K, fraccionado en dos aplicaciones. La primera fertilización se realizó a 60 días después del trasplante y la segunda fertilización a los 90 días después de la primera fertilización.
Con el objeto de evaluar la brotación en tubérculos de diferente grado de desarrollo, se realizaron cosechas a los 149, 179, 269 y 326 DDS. Estas fechas de cosecha fueron seleccionadas con base en reportes de Ile et al. (2006), quienes indican que las mismas corresponden a cuatro estados de desarrollo de los tubérculos, así: a) a los 149 DDS se obtienen tubérculos en formación temprana en proceso de llenado del parénquima interior; b) a los 179 DDS se obtienen tubérculos de mayor peso, casi completamente formados, en inicio de la formación de yemas, lo cual coincide con el momento de la entrada en senescencia de las plantas; c) a los 269 DDS se obtienen tubérculos completamente formados, con mayor grado de madurez y desarrollo, que coincide con el periodo de máxima actividad celular para la formación de yemas; y d) a los 326 DDS se obtienen tubérculos con yemas completamente formadas, posterior a la senescencia del cultivo. De los anteriores, Ile et al. (2006), indican que probablemente los tubérculos correspondientes a las dos primeras fechas de cosecha (149 y 179 DDS), se encuentran aún en estado de latencia o reposo, dado que los tejidos meristemáticos no se han desarrollado completamente. Por su parte, indican que los tubérculos de 269 y 326 DDS, aunque se encuentran en periodo de reposo natural, ya cuentan con las estructuras meristemáticas formadas y podrían responder a tratamientos para el rompimiento de la dormancia.
Para lo anterior, periódicamente se realizó la cosecha de 10 plantas por variedad y tres repeticiones. Se registró el peso de los tubérculos cosechados, producción por planta, longitud, peso y diámetro de los tubérculos. En total se realizaron cuatro cosechas escalonadas, a fin de obtener tubérculos para la evaluación de 149, 179, 269 y 326 DDS. En cada cosecha, una muestra de los tubérculos obtenidos se empacó en bolsas de papel kraft o de embalaje, y se transportaron en neveras a -4 y -8 °C, al Laboratorio de Microbiología Agrícola Centro de Investigación Turipaná de la Corporación Colombiana de Investigación Agropecuaria-Agrosavia. En laboratorio se hizo lavado con agua corriente y luego se realizó desinfección superficial del tubérculo con hipoclorito de sodio al 2%, se enjuagaron con agua destilada y se dejaron secar a la sombra. Posteriormente, se tomaron seis secciones de tubérculo de 4 mm de grosor y 10 g y se aplicaron los diferentes tratamientos. Para las evaluaciones se utilizaron secciones de la región basal del tubérculo, la cual se encuentra más cercana al punto de unión del tubérculo con el tallo, debido a que es la sección que presenta menor variabilidad en la brotación (Ile et al., 2006).
Aplicación de reguladores de crecimiento vegetal (RCV)
Se evaluó la aplicación de tres dosis de tres RCV sobre la brotación de tubérculos de ñame cosechados a los 149, 179, 269 y 326 días después del tratamiento (DDT), en dos variedades de ñame espino (0307-49SB y 0307-50CB). Los 10 tratamientos evaluados se muestran en el Cuadro 2.
Cuadro 2: Tratamientos para evaluar el efecto de reguladores de crecimiento vegetal en la ruptura de la dormancia de tubérculos de ñame.
No. |
Regulador |
Dosis |
Abreviatura |
1 |
Control |
Agua destilada |
Testigo |
2 |
Thiourea |
10.0 g L-1 |
TH10 |
3 |
Thiourea |
20.0 g L-1 |
TH20 |
4 |
Thiourea |
30.0 g L-1 |
TH30 |
5 |
Cloruro de mepiquat |
1.0 g L-1 |
CM1 |
6 |
Cloruro de mepiquat |
2.0 g L-1 |
CM2 |
7 |
Cloruro de mepiquat |
3.0 g L-1 |
CM3 |
8 |
Etefón |
0.5 g L-1 |
ET05 |
9 |
Etefón |
1.0 g L-1 |
ET1 |
10 |
Etefón |
2.0 g L-1 |
ET2 |
Para la aplicación de los tratamientos, se dividió longitudinalmente la región basal de cada tubérculo en seis porciones, obteniendo suficiente número de secciones basales para la aplicación de 10 tratamientos con tres repeticiones y la realización de tres muestreos a 7, 14 y 21 DDT (Craufurd et al., 2001; Ile, 20041). Se tomaron seis de secciones basales de los tubérculos y se sumergieron en las distintas concentraciones de los RCV, así: thiourea y cloruro de mepiquat durante 60 min y etefón durante 5 min. Previo a inmersión, a cada solución se le agregaron siete gotas del surfactante Tween 20® (polisorbato) por litro de solución. Estos tiempos de inmersión fueron seleccionados con base en experimentos previos realizados por Ile et al. (2006) y Hamadina y Craufurd, (2015). Una vez aplicados los tratamientos de RCV, las secciones de tubérculo fueron sumergidas en una mezcla de los fungicidas Ridomil Gold® 68WG (4% de Metalaxil-M y 64% de Mancozeb), en dosis de 4 g L-1 y Benlate® 50WP (Benomil 500 g kg-1), en dosis de 4 g L-1, durante 5 min. Dado el carácter poroso del tejido, se observó alto contenido de humedad en las secciones, por lo que luego del tratamiento con fungicidas, fue necesario dejar secar las secciones bajo la luz directa del sol durante 8 h por 3 días. Las secciones basales completamente secas se colocaron en cajas plásticas de 21×16×10 cm, con doble capa de papel toalla en el fondo, con el fin de proveer las condiciones para la brotación. Las cajas se recubrieron con una capa de papel de aluminio, a fin de mantener ambiente oscuro y uniforme, similar al de las condiciones de almacenamiento. Las cajas se mantuvieron en cuartos de bioensayo a temperatura de 28±2 °C y humedad relativa de 80%. Periódicamente, a los 7, 14 y 21 días después de la aplicación de los tratamientos, se retiraron 720 muestras o secciones de tubérculos para observar bajo microscopio la presencia (1) o ausencia (0) de signos de brotación en las secciones de tubérculo.
Preparación de muestras de tejido del tubérculo
Cada muestra para observación se preparó removiendo la superficie del tejido y tomado porciones de 5.0×0.9×0.9 cm, las cuales fueron diafanizadas con inmersión en agua por 15 min. Posteriormente, fueron introducidas en alcohol al 70% (1 h), alcohol al 90% (1 h), xileno (1 h) e incluidas en parafina líquida. Con la ayuda de un microtomo rotativo (American Optical Corporation, Spencer 820, Bethlehem, Pennsylvania, USA), se obtuvieron 720 muestras del tejido para observación. Posteriormente, las muestras fueron desparafinadas mediante hidratación, empleando concentraciones decrecientes de etanol (100, 96, 80, 50% y xileno); y teñidas con azul de alcián al 0.05%. Seguidamente, los tejidos fueron deshidratados en grupos crecientes de etanol (96, 100% y xileno). Los tejidos se observaron en microscopio binocular para campo claro DM500 (Leica®, USA) conectado a una cámara ICC50W (Leica®, USA). Al microscopio se registró el número de yemas brotadas en cada una de las secciones.
Análisis estadístico
Los análisis estadísticos se realizaron asumiendo un diseño experimental completamente al azar, en arreglo factorial 2×3×10, con tres repeticiones, en donde el factor A correspondió a las variedades de ñame, el B a los tubérculos de diferentes edades y el C los tratamientos con RCV. Los datos de brotación obtenidos en este ensayo presentaron una distribución no normal acorde a la Prueba Kolmogorov-Smirnov, por lo que se realizaron análisis de comparación múltiple mediante la prueba de Kruskal-Wallis con la opción de Wilcoxon con el valor P aproximado (chi-cuadrado), y prueba de comparaciones múltiples de Bonferroni. Debido a que los tubérculos de 149 DDS no mostraron signos de brotación, éstos no fueron considerados para los análisis estadísticos. A los datos de cosecha de los tubérculos en campo (longitud, peso fresco, diámetro, número de tubérculos por planta), se les realizó análisis de varianza y en los casos en que se detectaron diferencias estadísticas significativas usando nivel de significancia α=0.05, se realizaron pruebas de separación de medias utilizando la prueba HSD de Tukey. El análisis de los datos se realizó utilizando el paquete estadístico SAS v.9.4.
Resultados y Discusión
Tubérculos cosechados a las diferentes edades
La longitud, diámetro y peso de los tubérculos de ñame cosechados a los 149, 179, 269 y 326 días después de la siembra (DDS) en este experimento se muestran en el Cuadro 3. Los tubérculos cosechados a 149 y 179 DDS presentaron menor longitud, diámetro y peso fresco que los tubérculos cosechados a 269 y 326 DDS. Acorde a las observaciones, los tubérculos cosechados a 149 DDS presentaban menor tamaño, se encontraban apenas iniciando su formación, presentaban yemas inmaduras, y se encontraban en un estado que es comúnmente conocido como “cormo”. Por su parte los tubérculos cosechados a 179 DDS, presentaban mayor tamaño, se encontraban en proceso de acumulación activa de materia fresca y, al igual que los anteriores, mostraban algunos signos que indicaban inmadurez del tubérculo, debido a que el extremo distal presentaba color blanquecino, característico de crecimiento activo. Por el contrario, los tubérculos cosechados a los 269 y 326 DDS presentaron mayor tamaño y peso, con yemas desarrollada y coloración que indicaba que habían alcanzado su madurez fisiológica (Figura 1). Estos resultados en general coinciden con lo reportado por Ile et al. (2006), en cuanto al grado de desarrollo de los tubérculos.
Cuadro 3: Características de los tubérculos de dos variedades de ñame colectados a cuatro edades de cosecha.
Table 3: Tuber characteristics of two yam varieties collected at four harvesting ages.
Variedad |
Época de cosecha |
Longitud‡ |
Peso fresco |
Diámetro |
Tubérculos por planta |
cm |
g |
cm |
|||
0307-49SB |
149 |
3.72 c |
18.14 c |
2.55 d |
1.23 |
179 |
24.47 b |
1243.06 b |
7.79 c |
1.40 |
|
269 |
28.50 a |
2351.43 a |
11.48 a |
1.30 |
|
326 |
27.83 a |
2381.33 a |
10.09 b |
1.23 |
|
0307-50CB |
149 |
3.39 c |
13.77 c |
2.14 d |
1.13 |
179 |
20.12 b |
822.46 b |
6.30 c |
1.13 |
|
269 |
30.70 a |
2109.33 a |
10.75 a |
1.20 |
|
326 |
27.70 a |
1324.66 a |
7.98 b |
1.16 |
|
Media |
20.80 |
1283.02 |
7.3875 |
1.225 |
|
Dev. estándar |
2.7810 |
358.63 |
0.8517 |
0.1354 |
|
CV% |
13.37 |
27.96 |
11.53 |
11.05 |
|
Significancia |
† DDS = días después de siembra. ‡ Promedios con distinta letra dentro de cada columna indican diferencias a un nivel de significancia α = 0.05. **Altamente significativo (P ≤ 0.01); *significativo (P ≤ 0.05); NS = no significativo (P ≥ 0.05).
† DDS = Days after sowing. ‡ Averages with different letters within each column indicate differences at a significance level α = 0.05. ** Highly significant (P ≤ 0.01); * significant (P ≤ 0.05); NS = no significant (P ≥ 0.05).

Acorde a lo anterior, para este estudio se consideró que los tubérculos cosechados a 149 y 179 DDS corresponden a tubérculos inmaduros, en proceso de crecimiento, mientras que los colectados a 269 y 326 DDS, se consideran maduros, con yemas completamente desarrolladas, aptos para la cosecha. Las plantas de las cuales se obtuvieron los dos últimos tipos de tubérculo mostraron, en campo, síntomas de amarillamiento y secado del follaje, lo cual es considerado indicativo de madurez en el cultivo.
Brotación de las secciones del tubérculo
La evaluación de la brotación de las secciones de los tubérculos se realizó mediante muestreos destructivos realizados a 7, 14 y 21 días después de los tratamientos (DDT). Los resultados mostraron brotación solamente en los tubérculos de 179, 269 y 326 DDS. No se observó brotación en los tubérculos cosechados a 149 DDS (Figura 2), lo que se atribuyó a un insuficiente desarrollo e inmadurez de los tejidos meristemáticos, lo cual fue corroborado por las observaciones histológicas.
Figura 2: Número promedio de yemas con brotación en secciones de tubérculos cosechados a diferentes edades de desarrollo. Promedio de 10 tratamientos, dos variedades, tres evaluaciones y tres repeticiones (n = 180). La brotación en tubérculos de 149 días después de sembrados (DDS) fue nula y no se incluyó en los análisis. Las barras sobre columnas corresponden a error estándar y las letras encima de cada corresponden a la prueba de comparaciones múltiples de Bonferroni.

Figure 2: Average number of buds with sprouting in tubers harvested at different development ages. Average of 10 treatments, two varieties, three evaluations and three replications (n = 180). Sprouting in tubers 149 days after planting (DDS) was zero and was not included in the analyses. The bars over columns correspond to standard error and the letters above each correspond to the Bonferroni multiple comparison test.
Por el contrario, en los tubérculos cosechados a los 179, 269 y 326 DDS se observaron signos que confirmaron la brotación de las yemas (Figura 3).

Estos resultados confirman el reporte de Ile et al. (2006), en cuanto que los tubérculos de 149 DDS son aún inmaduros, con ausencia de tejidos meristemáticos y como consecuencia no muestran sensibilidad a la aplicación exógena de los RCV. Según Ile et al. (2006), este tipo de tubérculos se encuentran en una fase latente que es controlada por factores endógenos. En otras especies como papa, otros autores (Rodriguez y Moreno, 2010), han reportado que la latencia durante la formación temprana de los tubérculos está asociada con concentración de ácido abscísico (ABA) en el tejido vegetal, que se acumula activamente durante la formación de los tubérculos. Según estudios realizados por Sonnewald (2001) y Alcantara-Cortes et al. (2019), el ABA es una fitohormona considerada como el principal regulador del inicio de la formación de los tubérculos, la cual tiene la capacidad de inhibir y controlar la brotación de los tubérculos que aún se encuentran en estado de formación.
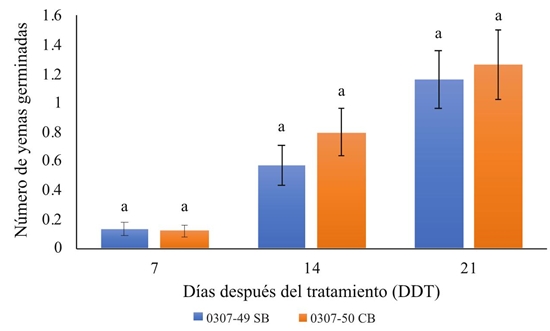
Con relación a los genotipos, no se presentaron diferencias estadísticas en la brotación, entre las dos variedades evaluadas en los muestreos realizados (Figura 4).
Figura 4: Número promedio de yemas con brotación en secciones de tubérculos cosechados a diferente edad de desarrollo en dos variedades de ñame. Promedio de 10 tratamientos, tres edades de los tubérculos y tres repeticiones (n = 90). Las barras sobre columnas corresponden al error estándar y las letras encima de cada corresponden a la prueba de comparaciones múltiples de Bonferroni.
Con relación a la aplicación RCV, se observó que los mayores efectos se observaron en los tubérculos de 269 y 326 DDS, indicando mayor sensibilidad de estos a su acción (Figura 5). Según estos resultados, aunque se observan algunos efectos sobre los tubérculos de 179 DDS, la respuesta a la aplicación exógena de RCV fue menor, posiblemente debido a que estos tubérculos presentan tejidos muy jóvenes, no suficientemente maduros para responder a las aplicaciones. Lo anterior coincide con resultados reportados por Hamadina y Craufurd (2015), quienes indican que la edad del tubérculo es un factor importante para considerar la aplicación exógena de RCV con fines de inducir la brotación, toda vez que su estado de desarrollo modera o regula el grado del efecto que se pueda obtener. Probablemente, la concentración de hormonas como el ABA en los tubérculos aún en desarrollo, impide la brotación de las yemas antes de completar el proceso de formación del tubérculo. A este respecto, los reportes de la literatura indican que se requiere la degradación paulatina del ABA, para que otras hormonas (como auxinas o giberelinas), puedan ejercer su función de incentivar la brotación en yemas y semillas en general (Kermode, 2005; Cheng et al., 2013).
Figura 5: Número promedio de yemas con brotación en secciones de tubérculos de tres edades de cosecha por efecto de la aplicación de Reguladores de Crecimiento Vegetal (RCV). CM corresponde a cloruro de mepiquat; ET corresponde a etefón; THIO corresponde a thiourea. Los números al lado de cada RCV corresponden a las concentraciones evaluadas, así: thiourea a 10, 20 y 30 g L-1 (TH10, TH20 y TH30), cloruro de mepiquat a 1.0, 2.0 y 3.0 g L-1 (CM1; CM2; CM3); etefón a 0.5, 1.0 y 2.0 g L-1 (ET05; ET1; ET2). Las barras muestran el promedio de dos variedades, tres repeticiones y tres muestreos realizados a los 7, 14 y 21 DDS. Las barras sobre columnas corresponden al error estándar (n = 18).
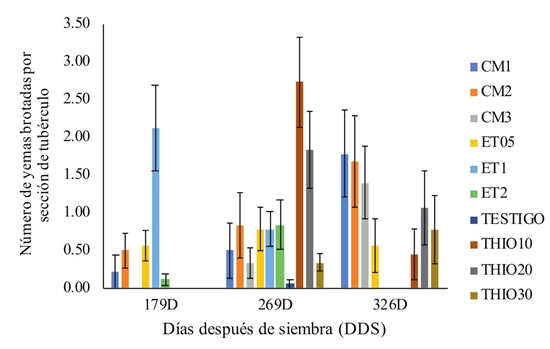
Figure 5: Average number of buds with sprouting in sections of tubers of three harvest ages because of plant growth regulator (RCV) application. CM corresponds to mepiquat chloride; ET corresponds to ethephon; THIO corresponds to thiourea. The numbers next to each RCV correspond to the concentrations evaluated, as follows: thiourea at 10, 20 and 30 g L-1 (TH10, TH20 and TH30), mepiquat chloride at 1.0, 2.0 and 3.0 g L-1 (CM1; CM2; CM3); ethephon at 0.5, 1.0 and 2.0 g L-1 (ET05; ET1; ET2). The bars show the average of two varieties, three repetitions and three samplings carried out at 7, 14, and 21 days after sowing. The bars on columns correspond to standard error (n = 18).
Con relación a los diferentes productos utilizados, se presentó interacción significativa (P ≤ 0.05) entre la edad de los tubérculos y los productos, indicando acción diferencial de los mismos, dependiendo del estado de desarrollo o edad de los tubérculos. En tubérculos de 179 DDS, los mayores efectos se obtuvieron con el etefón, mientras que en los tubérculos de 269 DDS, los mayores efectos se observaron con la thiourea. En tubérculos de mayor grado de madurez (326 DDS), el cloruro de mepiquat se muestra como el mejor RCV para inducir la brotación de los tubérculos (Figura 5).
A través de contrastes ortogonales se compararon los efectos de las diferentes dosificaciones de los RCV, encontrando mayor efecto de la dosis de 1.0 g L-1 de etefón (en tubérculos de 179 DDS), 10 g L-1 y 20 g L‑1 de thiourea (en tubérculos de 269 DDS) y 1.0 g L-1 de cloruro de mepiquat (en tubérculos de 369 DDS). Por otro lado, el tratamiento testigo (sin aplicación de RCV), no mostró ningún signo de brotación en los muestreos realizados, corroborando el estado de reposo natural en que se encontraban los tubérculos. Tan solo se observó cierto grado de brotación en los tubérculos de 269 DDS (Figura 5).
Los anteriores resultados coinciden con estudios realizados por Hamadina y Craufurd (2015), quienes mostraron que la aplicación del etefón solo o en combinación con thiourea acortó la duración de la brotación del ñame en aproximadamente 32 días. El etefón es un producto ampliamente utilizado en la agricultura para acelerar la maduración de los frutos, pero también se ha demostrado su efectividad en la ruptura de la dormancia en semillas y tubérculos como la papa (Degebasa, 2020). Una vez aplicado a las plantas, el etefón es hidrolizado y de este proceso se libera etileno, el cual es efectivo en dicho proceso (Weaver, 1972).
El cloruro de mepiquat es reportado como fitorregulador que altera la concentración de ácido giberélico (AG) en las células (Rademacher, 1991), debido a su inhibición parcial de una de las enzimas involucradas en su biosíntesis. El producto ha sido muy usado en algodón (Cadena y Orozco, 2002) para el control del crecimiento de las plantas. En el caso del ñame, se ha reportado que las giberelinas tienen un efecto inhibidor de la brotación en tubérculos de ñame (Tanno et al., 1992), por lo cual la aplicación exógena de inhibidores de su biosíntesis se considera eficaces en la ruptura de la dormancia (Shiwachi et al., 2003). Por lo anterior, en el presente estudio el cloruro de mepiquat posiblemente alteró las enzimas involucradas en la biosíntesis de giberelinas, actuando de manera eficaz en el rompimiento de la dormancia. La sensibilidad a este producto fue mayor en los tubérculos de ñame de 326 días después de la siembra.
La thiourea por su parte, es un compuesto órgano-sulfurado de estructura muy similar a la urea, excepto que el átomo de oxígeno ha sido reemplazado por el azufre, y ha sido reportado en plantas como un producto que a bajas concentraciones tiene la capacidad para romper la dormancia (Srivastava et al., 2016). Este producto ha sido usado ampliamente para acelerar la brotación de tubérculos de papa, en donde el periodo de brotación se ha reducido hasta en 12 días (Germchi et al., 2011). Bajo las condiciones del presente estudio, la thiourea fue efectiva en romper la dormancia e inducir la brotación de las secciones de tubérculos de ñame de 269 días después de la siembra.
Según estos resultados, el estado de reposo de los tubérculos de ñame puede ser interrumpido mediante la aplicación exógena de RCV, en cuyo caso algunos tratamientos resultaron ser más efectivos, dependiendo de la edad de la cosecha o madurez de los mismos. Desde el punto de vista práctico, podría ser posible obtener cultivos de ñame más uniformes mediante la siembra de tubérculos o secciones de ñame tratados con RCV, que aceleren la brotación de las yemas que se encuentran en reposo natural. Esto resolvería los problemas que enfrentan los agricultores en campo, en donde la brotación de los tubérculos sembrados es desuniforme, ocasionando cultivos con plantas con diferente grado de desarrollo, lo que ocasiona dificultades en el manejo y cosechas con alto grado de desuniformidad.
Conclusiones
Los reguladores de crecimiento vegetal (RCV) evaluados fueron efectivos en inducir la brotación de las yemas en los tubérculos de ñame espino, independientemente la variedad utilizada. Los efectos se observaron a partir de 14 días después del tratamiento y su acción fue efectiva solamente en tubérculos de 179, 269 y 326 días después de la siembra (DDS), lo que indica que se requiere de cierto grado de madurez en los tubérculos. No se encontró efectividad de los tratamientos en tubérculos de 146 días probablemente debido a su estado de inmadurez y pobre desarrollo. De lo anterior, se puede concluir que con el objeto de obtener semillas que respondan a la aplicación externa de RCV, se requiere de al menos un periodo de desarrollo de los tubérculos en campo ≥179 días después de la siembra.
En tubérculos de 179 días, el tratamiento más efectivo fue la aplicación de etefón en dosis de 1.0 g L‑1, mientras que en los tubérculos de 269 DDS, el tratamiento más efectivo fue la aplicación de thiourea en dosis de 10 y 20 g L-1. En tubérculos de mayor grado de madurez (326 DDS), el cloruro de mepiquat fue el tratamiento más efectivo, y en este último caso la dosis de mayor efectividad fue 1.0 g L-1.
Con estos resultados se puede concluir que es posible acelerar o inducir la brotación de las yemas del ñame que se encuentran en reposo a través de la aplicación de RCV, lo cual podría tener significancia, desde el punto de vista práctico, en la obtención de cultivos más uniformes y con tubérculos de mayor calidad (tamaño y uniformidad). Se requiere validar estos resultados bajo condiciones de campo en próximos estudios.
Fondos
Este estudio fue financiado por el Ministerio de Agricultura y Desarrollo Rural Colombiano bajo el convenio número TV18 y TV19 con AGROSAVIA. Este trabajo formó parte del proyecto “Desarrollo de tecnologías para el manejo y producción de semillas de calidad para mejorar la uniformidad y productividad del cultivo de ñame de la región Caribe”.
Contribución de los Autores
Para el presente artículo los autores han contribuido así: Conceptualización: J.C.T. y D.B.S.L. Metodología: D.B.S.L. Análisis: J.C.T. Validación: S.M.R.H. Laboratorio: L.L.L.C. Análisis formal: J.C.T. Investigación: S.M.R.H. y L.L.L.C. Recursos: D.B.S.L. Curación de datos: J.C.T. Preparación del borrador original: J.C.T. y D.B.S.L. Revisión y Edición: J.C.T. Visualización: J.C.T. y D.B.S.L. Supervisión: D.B.S.L. Administración del proyecto: D.B.S.L. Adquisición de fondos: D.B.S.L. y J.C.T.

