











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
La Selva Baja Caducifolia (SBC) es la vegetación con mayor distribución en México, se establece en la vertiente del Pacífico y es uno de los ecosistemas más amenazados, ya que presenta pocas selvas originales. Para el estado de Oaxaca es la vegetación más emblemática y representativa, tanto por la superficie que ocupa como por su presencia en distintas regiones, donde se concentra la mayor proporción de la población. Oaxaca es considerado como uno de los once estados que cuentan con una superficie considerable de selvas tropicales secas (5.84%) con una gran diversidad vegetal. De hecho, la más representativa se ubica en la región Costa en los distritos de Jamiltepec, Pochutla y Juquila (Salas et al., 2003; Torres-Colín, 2004). Son selvas con características muy particulares debido a que se encuentran en zonas con una marcada estacionalidad en la disponibilidad de agua, lo que hace que la riqueza florística del estado varié entre 8000 a 9000 especies (Campos et al., 1992; García-Mendoza et al., 1994, 2004). Sin embargo, es considerada como la vegetación más afectada por la práctica de roza-tumba-quema, que año con año genera la pérdida de grandes superficies para la conversión en áreas agrícolas y pecuarias. De igual forma, la expansión de los asentamientos humanos que consecuentemente han generado procesos de deforestación, fragmentación y degradación de las selvas. Este cambio en el uso del suelo ha provocado diferencias estructurales, en la composición, diversidad arbórea y a la estructura físico-química del suelo de los remanentes de estas selvas. Estas perturbaciones han confinado esta vegetación primaria a pequeños manchones y como resultado a la creación de grandes áreas de vegetación secundaria. Es importante mencionar la falta de estudios específicos en la región costa de Oaxaca, sin embargo, se tienen algunos trabajos realizados en la zona de Zimatán, Municipio de San Miguel del Puerto por Salas et al. (2003) y Salas et al. (2007) en el parque Nacional Huatulco; Domínguez et al. (2008) en la microcuenca del río Cacaluta. La mayor parte de los estudios florísticos están concentrados principalmente en las provincias del Istmo de Tehuantepec (Pérez-García et al., 2001; Gallardo et al., 2005), la región de la Mixteca Alta (García-Mendoza et al., 1994) y la región del valle de Tehuacán-Cuicatlán (Acosta et al., 2003). Considerando el desconocimiento en las especies arbóreas nativas y los procesos de conservación que se dan en estas selvas, el presente estudio ofrece una descripción de la estructura, composición y diversidad arbórea de selvas bajo condiciones de desarrollo en suelos Litosoles y Regosoles eutricos degradados como consecuencia del cambio de uso de suelos a la agricultura. Con base en la información generada se contará con el conocimiento de las especies arbóreas nativas con la finalidad de desarrollar estrategias en sistemas de producción que incluyan árboles como un componente del manejo integral del recurso suelo y con esto contribuir a la productividad y conservación de las selvas bajas caducifolias.
Materiales y Métodos
Se compararon dos condiciones de desarrollo de selvas bajas caducifolias con vegetación secundaria, ubicadas en la región Costa de Oaxaca. La primera selva (SBC-M) se localizó en el municipio de San Pedro Mixtepec en la comunidad de Mandingas, con coordenadas 15° 54’ 41” N y 97° 04’ 52” O; la segunda selva (SBC-P) se ubicó en las coordenadas 15° 46’ 14” N y 96° 26’ 50” O, en la comunidad de Chepilme, Municipio de San Pedro Pochutla, ambas áreas se destinaron a la actividad de roza-tumba-quema, la SBC-M por alrededor de 34 años mediante la siembra de cultivos de temporal como maíz, frijol, calabaza y sandía; la SBC-P se destinó a la agricultura por 21 años, posteriormente fue un basurero al aire libre por cuatro meses. Desde hace 13 años dichas selvas han sido conservadas.
Establecimiento de unidades de muestreo y toma de datos dasométricos
Las Unidades de Muestreo (UM) fueron localizadas en las áreas más conservadas en las dos selvas, en cada una de ellas se establecieron cuatro UM de 50 × 50 m (0.25 ha). Cada UM se dividió en 25 cuadros de 10 × 10 m (100 m²), dentro de cada cuadro se midieron todos los individuos arbóreos con un diámetro normal > 2.5 cm (DN = 1.30 m a partir del suelo) (Zarco-Espinosa et al., 2010). Las variables dasométricas registradas para cada individuo arbóreo fueron: diámetro normal (DN) medida con cinta diamétrica, altura total con clinómetro SUUNTO® y cobertura de copa (diámetro máximo y mínimo) con cinta métrica (Villavicencio-Enríquez y Valdez-Hernández, 2003). Se realizó la colecta botánica de hojas, flores y frutos por triplicado para la identificación taxonómica de las especies.
Estructura arbórea
Estructura vertical y horizontal. La estructura de la vegetación arbórea se representó por histogramas. Para describir la estructura vertical de los individuos registrados se consideraron clases de altura: 5, 10, 15 y 20; en el caso de la estructura horizontal se agruparon los DN por categorías diamétricas de 5 cm (5 = 2.6 ‑ 7.5, 10 = 7.6 - 12.5 hasta 45 = 42.6 - 47.5) (Ortega-Baranda et al., 2017).
Índices de valor estructural. Se utilizaron dos índices de valor estructural: el Índice de Valor de Importancia (IVI), que jerarquiza la dominancia de especies en rodales mezclados, el cual revela la importancia ecológica relativa de cada especie en una comunidad vegetal (Carreón-Santos y Valdez-Hernández, 2014).
IVI = Dominancia relativa + Densidad relativa + Frecuencia relativa
El Índice de Valor Forestal (IVF), se estimó con la finalidad de evaluar la estructura bidimensional de la vegetación arbórea en zonas cálido-húmedas de México, considerando el diámetro (plano horizontal, inferior), la altura (plano vertical, superior e inferior) y la cobertura (plano horizontal, superior) (Corella et al., 2001).
IVF = diámetro relativo + altura relativa + cobertura relativa
Riqueza y diversidad de especies
La riqueza registrada se determinó por el conteo de especies presentes en las UM. Con la finalidad de estimar la riqueza de especies arbóreas posibles que pudiera haber en cada condición de desarrollo, se realizaron curvas de acumulación de especies ajustadas por un modelo matemático logarítmico y potencial, así como un estimador no-paramétrico Chao2, (López-Gómez y Williams-Linera, 2006).
donde: S obs = riqueza de especies capturada mediante el muestreo; Q 1 = número de especies que fueron registradas solamente en una muestra del muestreo; Q 2 = número de especies que fueron registradas en dos muestras del muestreo.
La diversidad de especies se estimó con el Índice de Simpson (D p) modificado por Pielou con su complemento (1-D p), que manifiesta la probabilidad de que dos individuos seleccionados al azar de una comunidad finita (muestra) sean de diferentes especies (Pielou, 1967).
donde: n i = número de individuos de la especie en la muestra; N = número total de individuos de la muestra; S = total de especies en la muestra.
El Índice de Shannon-Weaver (H´), mide el grado promedio de incertidumbre para predecir a que especie pertenecerá un individuo escogido al azar de una superf icie muestreada (Magurran, 1988).
donde: Pi = proporción de individuos de la especie i.
Se construyeron curvas de rarefacción usando re-muestreos computacionales intensivos con la aplicación del software EcoSim (Entsminger, 2014), basándose en el número de individuos con la finalidad de estimar las métricas de diversidad y obtener diferencias estadísticas.
Para determinar el grado de homogeneidad existente en las abundancias relativas de las especies, se estimó la equidad (E) (Alcolado, 1998).
donde: H´= Índice de Shannon-Weaver; S = número total de especies en el muestreo.
El análisis comparativo entre la semejanza florística de pares de muestras entre UM se realizó mediante el Coeficiente de Sorensen (IS), que relaciona el número de especies presentes-ausentes entre UM (Godínez y López, 2002; Basáñez et al., 2008).
donde: C = número de especies comunes en ambas comunidades; A = número de especies encontradas en la comunidad A; B = número de especies encontradas en la comunidad B.
Muestreo y análisis de suelos
La prospección en campo permitió distinguir un relieve accidentado en la SBC-M, pero con una geoquímica uniforme del terreno. Con estos elementos se optó por establecer ocho monolitos (dos por UM) y se determinó la tasa de infiltración (TI) de agua en el suelo, mediante cinco pozos de absorción, distribuidos en las cuatro UM, con dimensiones de 0.30 m de diámetro y 0.40 m de profundidad. La tasa de infiltración se calculó desde la superficie del suelo hasta el nivel en el cual la columna de agua detuvo su descenso en el pozo, para lo cual se utilizó la siguiente fórmula propuesta por Gavande (1979):
donde: DT 2 -DT 1 = la distancia que desciende la columna de agua en el pozo, en el periodo de tiempo t 2 - t 1 .
Cada monolito de suelo se estableció como punto centrado a la UM y a partir de ahí se realizaron transectos de 250 m en zig-zag (Palmer y Troeh, 1980) estableciendo dos monolitos en cuatro sitios (ocho monolitos edáficos). La colecta de muestras de suelo se hizo en espesores de 10 cm, hasta una profundidad de 40 cm, lo que arrojó cuatro muestras por monolito para un total de 32 muestras de suelo. En cada monolito se colectaron 2 kg de suelo por espesor; las muestras se colocaron en bolsas de plástico cerradas e identificadas y posteriormente transportadas al área de Aguas y Suelos del Laboratorio de Tecnología de la Madera de la Universidad del Mar, Campus Puerto Escondido. Las muestras fueron secadas al aire, posteriormente se les tamizó con un mortero y una malla de 2 mm de diámetro. De cada muestra se tomaron 250 g de suelo para preparar pastas de saturación con agua destilada. A cada muestra se le determinó la textura por el hidrómetro de Bouyoucos, se les aplicó 25 mL de hidróxido de sodio 1 N y se procedió a su dispersión en una batidora ® Hamilton Beach durante 10 min, se prepararon probetas graduadas de 1 L de capacidad para verter la mezcla de agua y sedimentos, la interpretación se realizó con el triángulo de texturas (Soil Survey Staff, 2014). La densidad real (ϱ a ) y la densidad aparente (ϱ r ), fueron determinadas por el método de Pignómetro hídrico y de la Parafina, respectivamente (Soil Survey Staff (2014).
El porcentaje de espacio poroso se determinó usando la Ecuación (7):
donde: ϱ a = densidad aparente; ϱ r = densidad real.
El color del suelo se determinó mediante tablas de color Münsell (Münsell, 20001). Para los análisis químicos se siguió la metodología indicada por el manual Soil Survey Staff (2014). En este caso se obtuvieron extractos de pastas de saturación, en las que se determinó pH y Conductividad eléctrica (CE) mediante un multiparámetro® Hanna Instruments modelo HI98129, sólidos totales disueltos (STD) en mg L-1 mediante el método de calcinación en mufla a 600 °C. Cationes Ca2+, Mg2+ y aniones CO3 ‑2, HCO3 ‑1 y Cl-1, mediante el método titrimétrico; SO4 -2 por turbidimetría (espectofotómetro Perkin Elmer); Na+ y K+ por flamometría (Flamómetro IL-653). Los resultados de salinidad de las muestras de suelo fueron promediados obteniendo la desviación estándar y el coeficiente de variación. Con apoyo en el sistema de análisis estadístico SAS (2006), se practicó un análisis de varianza para establecer diferencias significativas de valores de salinidad entre espesores y entre monolitos de suelo utilizando el modelo Tukey α = 0.05.
Resultados y Discusión
La composición de especies para la SBC-M fue de 18 familias con 30 géneros y 33 especies, para la SBC-P se registraron 21 familias con 44 géneros y 47 especies. Las familias más representativas para ambas selvas fueron: Fabaceae (23 y 50%), Anacardiaceae (6%), Burseraceae (4 y 6%), Malvaceae (4 y 3%) y Rubiaceae (4 y 3%) respectivamente. Las Fabaceas son representativas de este tipo de ecosistemas ya que se encuentra influenciada por las temperaturas en regiones tropicales y subtropicales, como los estados de Nayarit con 21% (Bravo et al., 2013), Michoacán con 48 y 38% (Martínez-Cruz et al., 2013; Casanova-Lugo et al., 2014), Guerrero con 30% (Pineda et al., 2007), Oaxaca con 15 y 13% (Pérez-García et al., 2001; Gallardo et al., 2005), San Luis Potosí con 31% (Alanís et al., 2010) y Yucatán con 49% (Ceccon et al., 2002).
Estructura vertical y horizontal. En lo que concierne a la distribución de los individuos arbóreos respecto a la estructura vertical se tuvo una tendencia hacia alturas entre 1‑10 m, sin embargo, la SBC-M registró un mayor número de individuos en el intervalo de 5.1‑10 m (Figura 1). La estructura horizontal se caracterizó por que la mayoría de los individuos en ambas selvas estuvieron dentro de la categoría diamétrica 5. Con una tendencia hacia la representación gráfica de una J invertida (Figura 1), lo que confirma que el desarrollo de estas selvas es inicial y con buena regeneración de la vegetación, así como con condiciones ecológicas que propician el desarrollo (Guariguata y Ostertag, 2001; Carreón-Santos y Valdez-Hernández, 2014). Similares condiciones se reportan para la región de Zimatlán en Oaxaca (Salas et al., 2003), Jalisco (Sánchez et al., 2002), Tabasco (Zarco-Espinosa et al., 2010) y Chiapas (López-Toledo et al., 2012), en donde la mayoría de los individuos se concentraron en los diámetros más pequeños y disminuyeron hacia los diámetros mayores.
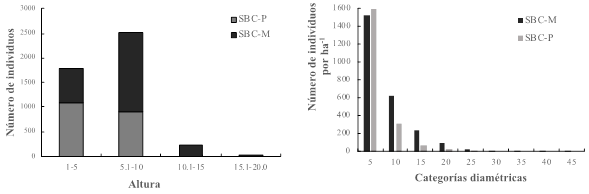
Figura 1: Estratificación vertical y horizontal de la vegetación arbórea en dos condiciones
de desarrollo de selvas bajas caducifolias en la costa de
Oaxaca.
Figure 1: Vertical and horizontal
stratification of tree vegetation in two conditions of development
of low deciduous forests on the coast of Oaxaca
Índices de valor estructural. Un resultado porcentual alto en la importancia ecológica (IVI) en la SBC-P y SBC-M, se determinó para las especies con mayor valor de dominancia relativa: Ceiba parvifolia Rose (IVI = 11.10 y 16.45%) y Heliocarpus donnell smithii Rose (IVI = 5.86 y 8.78%). De igual forma, altos valores en la densidad relativa se obtuvieron en Dalbergia congestiflora Pittier (IVI = 11.52 y 9.98%). Las especies con mayor IVF fueron las mismas que en el IVI (Cuadro 1). Tanto C. parvifolia y D. congestiflora (especies intolerantes a la sombra), se presentan en la sucesión temprana y son aptas para sistemas de enriquecimiento en bosques secos muy perturbados (Ochoa-Gaona et al., 2007; Alanís et al., 2010; Zamora-Crescencio et al., 2011). Así mismo, Amphipterygium adstringens Schiede ex Schltdl se suma a las especies con valores altos de IVI (37.75%) e IVF (38.45%) para SBC-M (Cuadro 1). Junto con D. congestiflora son especies que tienen la capacidad de regenerarse a partir de raíces o rebrotes (policaulescencia), las cuales tienden a dominar los bosques tropicales secos (Murphy y Lugo, 1986; Miller y Kauffman, 1998; Gallardo et al., 2005). La policaulescencia les permite aumentar la abundancia y formar colonias muy características de este tipo de vegetación, incrementando la densidad de individuos como se observó en este estudio. En el caso de D. congestiflora sus poblaciones se ven amenazadas por la tala clandestina de su madera que es altamente demandada para la realización de instrumentos musicales y artesanales, de similar forma, A. adstringens es amenazada por un aprovechamiento irracional de la corteza con fines medicinales en el estado de Oaxaca.
Cuadro 1: Índice de valor de importancia y valor forestal en dos condiciones de desarrollo
arbóreo en selvas bajas caducifolias en la costa de
Oaxaca.
Table 1: Index of importance value and
forest value in two conditions of tree development in low deciduous
forests on the coast of Oaxaca.
| Especie / condición de desarrollo SBC-P | IVI | Especie / condición de desarrollo SBC-P | IVF |
|---|---|---|---|
| Dalbergia congestiflora Pittier | 34.56 | Dalbergia congestiflora Pittier | 42.16 |
| Ceiba parvifolia Rose | 33.29 | Ceiba parvifolia Rose | 35.27 |
| Guapira petenensis (Lundell) Lundell | 20.49 | Bursera simaruba (L.) Sarg. | 22.23 |
| Heliocarpus donnell smithii Rose | 17.59 | Acacia farnesiana (L.) Willd. | 19.07 |
| Hintonia latiflora (Sessé & Moc. ex Dc.) Bullock | 14.41 | Heliocarpus donnell smithii Rose | 19.06 |
| Gliricidia sepium (Jacq.) Kunth ex Walp. | 13.41 | Guapira petenensis (Lundell) Lundell | 17.26 |
| Vitex hemsleyi Briq. | 13.29 | Comocladia engleriana Loes | 16.92 |
| Comocladia engleriana Loes | 9.97 | Spondias purpurea L. | 10.96 |
| Acacia farnesiana (L.) Willd. | 9.75 | Guazuma ulmifolia Lam | 8.09 |
| Ruprechtia fusca Fernald | 9.03 | Hintonia latiflora Moc &Sessé | 6.12 |
| 37 especies restantes | 37 especies restantes | ||
| Especie / condición de desarrollo SBC-M | IVI | Especie / condición de desarrollo SBC-M | IVF |
| Ceiba parvifolia Rose | 49.36 | Ceiba parvifolia Rose | 57.24 |
| Amphipterygium adstringens Schiede ex Schltdl | 37.74 | Dalbergia congestiflora Pittier | 38.91 |
| Cochlospermum vitifolium (Willd) Spreng | 31.82 | Amphipterygium adstringens Schiede ex Schltdl | 38.44 |
| Dalbergia congestifloraPittier | 29.91 | Heliocarpus donnell-smithii Rose | 25.03 |
| Heliocarpus donnell-smithii Rose | 26.35 | Cochlospermum vitifolium (Willd) Spreng | 23.27 |
| Acacia farnesiana Wall. | 14.43 | Acacia farnesiana Wall. | 17.31 |
| Comocladia engleriana Loes | 11.05 | Tabebuia chrysantha (Jacq.) G. Nicholson | 10.42 |
| Tabebuia chrysantha (Jacq.) G. Nicholson | 10.82 | Comocladia engleriana Loes | 10.23 |
| Guapira macrocarpa (Miranda) Miranda | 10.81 | Guapira macrocarpa (Miranda) Miranda | 9.79 |
| Bunchosia sp. | 10.38 | Hintonia latiflora (Sesse & Moc. Ex. DC.) Bullock | 8.26 |
| 23 especies restantes | 23 especies restantes |
Riqueza y diversidad de especies. Con base en las métricas de diversidad (Cuadro 2) y las curvas de rarefacción basadas en el número de individuos, se tiene que ambas condiciones de desarrollo presentaron diferencias significativas (P < 0.05) en la riqueza de especies, pero no en el Índice de Shannon-Weaver (Figura 2).
Cuadro 2: Riqueza registrada y estimada, índices de diversidad, equidad, dominancia y
similitud, en dos condiciones de desarrollo arbóreo en selvas bajas
caducifolias en la costa de Oaxaca.
Table 2:
Registered and estimated wealth, diversity indices, equity,
dominance and similarity, under two tree development conditions in
low deciduous forests on the Oaxaca coast.
| Condición de desarrollo | Riqueza registrada | Riqueza estimada | |||
|---|---|---|---|---|---|
| Paramétrico | No-paramétrico | ||||
| 15 | 30 | Riqueza estimada con Chao2 | Número de especies faltantes | ||
| SBC-M | 33 | 37 | 40 | 42 | 9 |
| SBC-P | 47 | 56 | 70 | 63 | 16 |
| Complemento del Índice de Simpson modificado por Pielou (I-D p ) | Índice de Shannon- Weaver (H´) | Equidad (JH’) | Dominancia | Índice de Sorensen | |
| SBC-M | 0.911 | 2.731 | 0.781 | 0.157 | 0.60 |
| SBC-P | 0.919 | 3.003 | 0.784 | 0.219 | |

Figura 2: Curvas de rarefacción de riqueza e Índice de Shannon-Weaver, con el software
EcoSim en dos condiciones de desarrollo arbóreo en selvas bajas
caducifolias en la costa de Oaxaca. Las barras de error representan
los intervalos de confianza (P <
0.05).
Figure 2: Wealth rarefaction curves and
the Shannon-Weaver Index, with the EcoSim software under two tree
development conditions in low deciduous forests on the Oaxaca coast.
The error bars represent the confidence intervals
(P < 0.05).
Sin embargo, se identificaron porciones en donde los intervalos de confianza (95%) se solapan entre comunidades, lo que indica la existencia de un tamaño muestra crítico a partir del cual estas difieren en términos de su riqueza. En el caso de las métricas de diversidad fueron menores en comparación con trabajos para el estado de Oaxaca como: Trejo y Dirzo (2002) en cuatro localidades, con registros de riqueza (52 -76 especies), H´ (3.54 - 3.98) y valor inverso de D (23.46 - 30.38); Salas et al. (2003) en Zimatán con una riqueza de 71 especies y Gallardo et al. (2005) que registraron 194 especies, con valores de H´ semejantes con la condición de desarrollo SBC-M (2.73), pero con una equidad mayor (J H´ = 0.82) a diferencia del presente estudio (J H´ = 0.78). Con lo anterior se establece que las especies en las dos condiciones de desarrollo tuvieron menor representatividad en términos de abundancia relativa. Sin embargo, López-Toledo et al. (2012) en áreas con manejo pecuario en el estado de Chiapas registraron una riqueza de 25 especies, H´= 2.64 y J H´ = 0.82; Zamora-Crescencio et al. (2011) determinaron una riqueza de 43 especies con una H´= 1.59 en un área con 10 años de abandono por agricultura y Dzib-Castillo et al. (2014) con 50 especies y un H´= 2.31 en áreas afectadas por agricultura, ambos en el estado de Campeche. De acuerdo con lo anterior expuesto, se deriva que la SBC-M y SBC-P se encuentran en un estado de desarrollo semejante con selvas perturbadas en proceso de conservación. Por consiguiente, las diferencias que se presenten en las métricas de diversidad estarán relacionadas con las condiciones ecológicas del hábitat y los cambios en el uso del suelo de cada sitio de estudio (Zamora-Crescencio et al., 2011). Considerando lo anterior y al comparar el estimador de Chao2 y el análisis de regresión para la riqueza estimada (Figura 3), se puede llegar a predecir un aproximado de 63 hasta 70 especies con 30 UM para la condición de desarrollo de SBC-P (con mayor riqueza) (Cuadro 2). Ambas condiciones de desarrollo tuvieron una disimilitud del 40% de especies.

Figura 3: Curvas de acumulación de especies en dos condiciones de desarrollo arbóreo en
selvas bajas caducifolias en la costa de
Oaxaca.
Figure 3: Species accumulation curves
under two tree development conditions in low deciduous forests from
the Oaxaca coast.
En lo que respecta a la equidad y dominancia no se tuvieron diferencias significativas (P < 0.05) entre condiciones. En este sentido las especies están más cercanas a estar igualmente representadas en ambas condiciones de desarrollo.
Características geomorfológicas, texturales y físico-químicas de los suelos de la SBC-M
Geomorfología. Un análisis anisotrópico vertical descendente de los monolitos de suelo de la SBC-M, permitió distinguir suelos poco profundos y de escaso desarrollo, los cuales se constituyen por dos horizontes identificados como A y B, formados por intemperismo de rocas metamórficas del paleozóico pertenecientes a la formación geológica Xolapa (De Cserna, 1965).
El horizonte A presentó un subhorizonte Oa con acumulación de residuos de materia orgánica fresca con descomposición rápida debido a factores ambientales como la radiación solar, la temperatura y la humedad, así como por la actividad microbiana del suelo; lo que originó una pobre acumulación de materia orgánica. Debajo de la incipiente capa húmica se localizó el sub-horizonte Oi, en el que se observó un espesor integrado de materia orgánica en descomposición y materia mineral. Las partículas que predominaron fueron las arenas en más de un 80%, seguidas de limos y arcillas; ambos sub-horizontes conforman un delgado espesor de 4 cm.
Por su parte el horizonte B con profundidad aproximada de 1 m, presentó arcillas caolinitas y óxidos e hidróxido de hierro y aluminio que se acumularon en su parte media. El horizonte C se conformó por silicatos primarios intemperizados y la roca madre. De acuerdo con López (1993), el relieve de bajo-lomeríos es resultado de levantamientos de la corteza emergida del mar, ocurridos en el Pli-pleistoceno, que derivó en la oxidación de minerales como el hierro (Fe2O3), responsable de la coloración rojiza de minerales como la hematita, la gohetita y del propio suelo (FitzPatrick, 1980).
Textura, espacio poroso y velocidad de infiltración. Se identificaron en los monolitos 1 al 6 una alternancia textural migajón arenosa a areno migajonosa presente con regularidad en el espesor 0-40 cm; debido a ligeras diferencias porcentuales en la acumulación de limos y arcillas, igualmente presente en el monolito 7. En cambio, el monolito 8, presentó una disposición bi textural en los primeros 20 cm de suelo, donde predominó una textura arena migajonosa, mientras que los 20 cm restantes, tuvo una textura migajón limosa. Esto se atribuye a la diferente posición de los suelos en el terreno, ya que las UM-I, UM-II y UM-III presentaron un relieve accidentado con pendientes > 20%, que los hace vulnerables a procesos de remoción de partículas minerales en época de lluvias, mientras que en la UM-IV, se encontró una toposecuencia baja. Esta posición permite procesos de acumulación de sedimentos provenientes de la hidroerosión en época de lluvias, que favorecen el arrastre y depósito de sedimentos aguas abajo, formando suelo inconsolidado proveniente de los materiales de arrastre de los lomeríos contiguos para la UM-IV.
La velocidad de infiltración fue de 4.5 cm h-1, dato que es consistente con valores de obtenidos por Sánchez-Bernal et al. (2013), para suelos litosoles en lomeríos de llanura costera. Caracterizados por su impermebilidad en suma con pendiente > 20% son la causa de que la escorrentía superficial se encargue de erosionar los primeros 20 cm de la capa superficial del suelo. Lo anterior se relaciona con una baja capacidad de almacenamiento de agua que los identifica como suelos hidrófobos; lo descrito tiene relevancia porque en época de estiaje las especies vegetales establecidas en estos suelos sufren estrés hídrico.
Color del suelo. Los colores 2.5 YR 4/2 y 4/3 en seco, y 7.5 YR 4/2 y 4/3 en húmedo de los espesores 0-20 cm y 20-40 cm que corresponden a los suelos de las UM‑I, UM-II y UM-III, indican procesos de lavado de cationes básicos, los cuales se pierden en el agua de drenaje, este fenómeno se conoce como latosolización y acompaña a valores de pH de suelos ácidos tropicales (Buckman y Brady, 1977). También indica que la oxidación origina en lo fundamental Fe2O3, de común ocurrencia en suelos tropicales con vegetación forestal y es constituyente de los minerales goethita, hematita y caolinita, mismos que imparten color rojo a los suelos.
pH, CE, STD y (( de soluciones de suelos. El pH de los suelos en las cuatro UM fue ligeramente ácido, el valor promedio es de 6.78 (Cuadro 3). Esto indica que en la SBC-M tienen lugar reacciones de precipitación de hidróxido de hierro y aluminio, asociados a una tipogénesis latosólica caracterizada por un pobre contenido de cationes básicos Ca2+, Mg2+, Na+ y K+ lixiviados en época de lluvias en el agua de drenaje que se acumulan en la parte baja de la microcuenca. En la UM-IV se confirmó los extractos acuosos de suelos con pequeñas concentraciones de cationes y aniones. La CE de los extractos acuosos de las pastas de saturación de los suelos analizados en los cuatro cuadrantes, resultó en un valor promedio de 0.55 dS m-1, lo que indica que no presentan problemas de salinidad. Un ligero incremento de CE se observa en el espesor 0-10 cm, que es natural, dado que las aguas capilares ascienden y se evaporan en la superficie, por lo que no existe riesgo de salinización (Sánchez-Bernal et al., 2013). La concentración de STD en la solución del suelo es muy baja, en promedio 346 mg L-1, lo que indica que la concentración iónica está presente en cantidades traza, esto se debe a que los procesos erosivos y de lixiviación iónica se encargan de remover los iones minerales aguas abajo, por lo que en época de lluvias la pérdida de cationes y aniones básicos es intensa. El potencial osmótico de las soluciones de los suelos es muy bajo, en promedio Ѱπ = -0.01 MPa, valor asociado a los diminutos valores de CE, esto indica una pobre presencia de iones minerales (Richards, 1985).
Cuadro 3: Características físico-químicas de los monolitos de suelos analizados antes de
la época de lluvias en la condición de desarrollo de SBC-M en la
costa de Oaxaca.
Table 3: Physico-chemical
characteristics of the soil monoliths analyzed before the rainy
season in the development condition of SBC-M from the coast of
Oaxaca.
| Monolito / UM | Espesor de suelo | pH | CE | STD | Ѱπ | Parámetros estadísticos | |
|---|---|---|---|---|---|---|---|
| cm | dS m-1 | mg L-1 | MPa | CE | STD | ||
| M1 UM-I | (0-10] | 6.76 | 1.33 | 851 | -0.047 | M= 0.75 | M= 479.5 |
| (10-20] | 7.03 | 0.77 | 492 | -0.027 | σ = 0.41 | σ = 266.58 | |
| (20-30] | 6.63 | 0.49 | 313 | -0.017 | CV = 0.54 | CV = 0.55 | |
| (30-40] | 7.04 | 0.41 | 262 | -0.014 | |||
| M2 UM-I | (0-10] | 6.13 | 0.78 | 499 | -0.028 | M = 0.52 | M = 335.75 |
| (10-20] | 6.27 | 0.63 | 403 | -0.022 | σ = 0.21 | Σ = 139.94 | |
| (20-30] | 6.20 | 0.38 | 243 | -0.013 | CV = 0.40 | CV = 0.41 | |
| (30-40] | 6.71 | 0.31 | 198 | -0.011 | |||
| M3 UM-II | (0-10] | 6.86 | 1.15 | 736 | -0.041 | M = 0.53 | M = 340.5 |
| (10-20] | 7.67 | 0.39 | 249 | -0.014 | Σ = 0.41 | Σ = 265.31 | |
| (20-30] | 6.44 | 0.31 | 198 | -0.011 | CV = 0.77 | CV = 0.77 | |
| (30-40] | 6.30 | 0.28 | 179 | -0.010 | |||
| M4 UM-II | (0-10] | 6.78 | 1.28 | 819 | -0.046 | M = 0.54 | M = 346.7 |
| (10-20] | 7.79 | 0.37 | 236 | -0.013 | σ = 0.48 | σ = 320.08 | |
| (20-30] | 6.84 | 0.35 | 224 | -0.012 | CV = 0.88 | CV = 0.92 | |
| (30-40] | 7.24 | 0.17 | 108 | 0.000 | |||
| M5 UM-III | (0-10] | 7.80 | 0.44 | 281 | -0.015 | M = 0.39 | M = 252.5 |
| (10-20] | 6.79 | 0.38 | 243 | -0.013 | σ = 0.03 | σ = 21.76 | |
| (20-30] | 6.55 | 0.40 | 256 | -0.144 | CV = 0.08 | CV = 0.08 | |
| (30-40] | 6.53 | 0.36 | 230 | -0.012 | |||
| M6 UM-III | (0-10] | 7.16 | 0.84 | 537 | -0.030 | M = 0.55 | M = 351.5 |
| (10-20] | 6.53 | 0.45 | 288 | -0.016 | σ = 0.19 | σ = 123.91 | |
| (20-30] | 6.98 | 0.47 | 300 | -0.016 | CV = 0.34 | CV = 0.35 | |
| (30-40] | 6.66 | 0.44 | 281 | -0.015 | |||
| M7 UM-IV | (0-10] | 6.53 | 1.03 | 659 | -0.037 | M = 0.65 | M = 418.7 |
| (10-20] | 6.52 | 0.59 | 377 | -0.021 | σ = 0.23 | σ = 162.58 | |
| (20-30] | 6.51 | 0.49 | 313 | -0.017 | CV = 0.35 | CV = 0.38 | |
| (30-40] | 7.10 | 0.51 | 326 | -0.018 | |||
| M8 UM-IV | (0-10] | 7.04 | 0.60 | 384 | -0.021 | M = 0.38 | M = 246 |
| (10-20] | 6.94 | 0.40 | 256 | -0.014 | σ= 0.16 | Σ = 100.15 | |
| (20-30] | 6.11 | 0.27 | 172 | -0.009 | CV = 0.42 | CV = 0.40 | |
| (30-40] | 6.68 | 0.27 | 172 | -0.009 | |||
M = media de parámetros CE y Ѱπ; σ = desviación estándar de parámetros CE y Ѱπ; CV = coeficiente de variación de parámetros CE y Ѱπ.
M = mean of CE and Ѱπ parameters; σ = standard deviation of parameters CE and Ѱπ; CV = parameter variation coefficient CE and Ѱπ.
Composición iónica de pastas de saturación del suelo. La composición geoquímica de los extractos de pastas de saturación de los suelos analizados es típica de suelos formados por intemperismo de rocas metamórficas cálcico-magnésicas que forman parte de la estructura geológica de la Sierra Madre del Sur (López, 1993).
En este caso el orden catiónico fue de Ca2+ > Mg2+ > Na+ > K+ y el aniónico HCO3 - > Cl- > SO4 -2. De hecho, este orden guarda consistencia con la capacidad de migración de los iones en solución, pues los iones Na+ y K+, así como los aniones Cl- y SO4 -2, migran más rápido en solución que los de Ca2+ y Mg2+, lo cual es congruente con lo señalado por Szabolcs (1994) y se constata en los datos del Cuadro 4. De acuerdo con la clasificación de suelos de la FAO (2015), en las UM- I, UM-II y UM-III de SBC-M los suelos son de tipo litosol; mientras que para la UM-IV dada la transición de lomerío a piedemonte, donde se presentan incipientes procesos de depósito y acumulación, está ocurriendo en el tiempo una transición hacia un suelo regosol eutrico.
Cuadro 4: Composición química de extractos acuosos de pastas de saturación de suelos
analizados en la condición de desarrollo SBC-M, en la costa de
Oaxaca.
Table 4: Chemical composition of
aqueous extracts of soil saturation pastes analyzed in the SBC-M
development condition, from the coast of Oaxaca.
| Monolito/UM | Espesor | Cationes | Total | Aniones | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | Mg2+ | Na+ | K+ | CO3 -2 | HCO3 -1 | Cl-1 | SO4 -2 | ||||
| cm | - - - - - - - - - - - - - - - - - - - - - mEq 100 g-1 de suelo - - - - - - - - - - - - - - - - - - - - - | ||||||||||
| M1 UM-I | (0-10] | 0.0016 | 0.0002 | 0.0014 | 0.0003 | 0.0035 | 0.0000 | 0.0003 | 0.0004 | 0.0000 | 0.0007 |
| (10-20] | 0.0007 | 0.0002 | 0.0005 | 0.0001 | 0.0015 | 0.0000 | 0.0002 | 0.0004 | 0.0000 | 0.0006 | |
| (20-30] | 0.0009 | 0.0004 | 0.0007 | 0.0001 | 0.0021 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (30-40] | 0.0001 | 0.0002 | 0.0001 | 0.0001 | 0.0005 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| M2 UM-I | (0-10] | 0.0012 | 0.0010 | 0.0002 | 0.0000 | 0.0024 | 0.0000 | 0.0003 | 0.0001 | 0.0000 | 0.0004 |
| (10-20] | 0.0009 | 0.0003 | 0.0005 | 0.0005 | 0.0022 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (20-30] | 0.0005 | 0.0011 | 0.0003 | 0.0000 | 0.0019 | 0.0000 | 0.0003 | 0.0000 | 0.0000 | 0.0003 | |
| (30-40] | 0.0007 | 0.0002 | 0.0004 | 0.0000 | 0.0013 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| M3 UM-II | (0-10] | 0.0021 | 0.0003 | 0.0002 | 0.0002 | 0.0028 | 0.0000 | 0.0002 | 0.0000 | 0.0003 | 0.0005 |
| (10-20] | 0.0004 | 0.0012 | 0.0001 | 0.0001 | 0.0018 | 0.0000 | 0.0002 | 0.0001 | 0.0002 | 0.0005 | |
| (20-30] | 0.0004 | 0.0013 | 0.0001 | 0.0001 | 0.0019 | 0.0000 | 0.0002 | 0.0001 | 0.0002 | 0.0005 | |
| (30-40] | 0.0014 | 0.0011 | 0.0002 | 0.0001 | 0.0028 | 0.0000 | 0.0002 | 0.0001 | 0.0002 | 0.0005 | |
| M4 UM-II | (0-10] | 0.0009 | 0.0003 | 0.0000 | 0.0002 | 0.0014 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 |
| (10-20] | 0.0007 | 0.0004 | 0.0000 | 0.0000 | 0.0011 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (20-30] | 0.0010 | 0.0003 | 0.0000 | 0.0000 | 0.0013 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (30-40] | 0.0014 | 0.0002 | 0.0000 | 0.0000 | 0.0016 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| M5 UM-III | (0-10] | 0.0010 | 0.0004 | 0.0005 | 0.0002 | 0.0021 | 0.0000 | 0.0002 | 0.0000 | 0.0005 | 0.0007 |
| (10-20] | 0.0012 | 0.0003 | 0.0007 | 0.0002 | 0.0024 | 0.0000 | 0.0002 | 0.0000 | 0.0004 | 0.0006 | |
| (20-30] | 0.0015 | 0.0004 | 0.0003 | 0.0001 | 0.0023 | 0.0000 | 0.0002 | 0.0000 | 0.0001 | 0.0003 | |
| (30-40] | 0.0010 | 0.0006 | 0.0003 | 0.0001 | 0.0020 | 0.0000 | 0.0002 | 0.0000 | 0.0001 | 0.0003 | |
| M6 UM-III | (0-10] | 0.0019 | 0.0001 | 0.0004 | 0.0001 | 0.0026 | 0.0000 | 0.0002 | 0.0001 | 0.0000 | 0.0003 |
| (10-20] | 0.0011 | 0.0002 | 0.0002 | 0.0000 | 0.0015 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (20-30] | 0.0010 | 0.0001 | 0.0001 | 0.0000 | 0.0012 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (30-40] | 0.0008 | 0.0002 | 0.0001 | 0.0000 | 0.0011 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| M7 UM-IV | (0-10] | 0.0002 | 0.0008 | 0.0002 | 0.0022 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (10-20] | 0.0008 | 0.0003 | 0.0005 | 0.0001 | 0.0017 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (20-30] | 0.0006 | 0.0003 | 0.0001 | 0.0000 | 0.0010 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (30-40] | 0.0015 | 0.0003 | 0.0006 | 0.0001 | 0.0025 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| M8 UM-IV | (0-10] | 0.0015 | 0.0002 | 0.0003 | 0.0001 | 0.0021 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 |
| (10-20] | 0.0008 | 0.0000 | 0.0004 | 0.0000 | 0.0012 | 0.0000 | 0.0002 | 0.0001 | 0.0000 | 0.0003 | |
| (20-30] | 0.0010 | 0.0004 | 0.0007 | 0.0001 | 0.0022 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
| (30-40] | 0.0011 | 0.0003 | 0.0002 | 0.0000 | 0.0016 | 0.0000 | 0.0002 | 0.0000 | 0.0000 | 0.0002 | |
Vocación de uso del suelo. La poca profundidad de los suelos litosoles de las UM-I, UM-II y UM-III, los hace intolerantes a la erosión hídrica, al respecto Sánchez-Bernal et al. (2013), determinaron una erosión de 16 Mg ha-1, para suelos de la ranchería “El Guarumbo”, en San Pedro Mixtepec, Oaxaca, próximos a la condición de desarrollo de SBC-M. De igual manera la poca capacidad de almacenamiento hídrica de estos suelos está asociada con la presencia de una vegetación de SBC.
Precisamente la pérdida de suelo y las condiciones semiáridas del estiaje caracterizan su bajo valor económico. De acuerdo con lo anterior, es necesario implementar más trabajos de repoblación, sobre todo en la UM-IV de la SBC-M, ya que la reforestación puede favorecer una mayor cubierta vegetal, la capacidad de captar y almacenar agua de lluvia y de controlar la erosión del suelo. En cuanto a técnicas de manejo y conservación del suelo, es recomendable establecer en los conos de arroyada más propicios (por tamaño y volumen de captación hídrica), tinas de captación y almacenamiento de agua y canales que provean del vital líquido a los árboles en épocas de secano. Así mismo en los terrenos con pendientes > 20%, se sugiere el establecimiento de vegetación en curvas de nivel, para favorecer el control de la erosión hídrica.
Conclusiones
- Las condiciones de desarrollo de la vegetación arbórea en la SBC-M y SBC-P presentaron diferencias significativas (P < 0.05) en la riqueza de especies. En tanto que las métricas de diversidad de especies fueron bajas en comparación con selvas en estados más avanzado de conservación, sin embargo, se encuentran dentro de los intervalos promedio reportados para varios sitios de México con las mismas condiciones de desarrollo estudiadas. De modo que, la estructura vertical y horizontal también mostraron condiciones de crecimiento que confirman un buen desarrollo inicial con buena regeneración propios de una SBC con vegetación secundaria.
- Los suelos de la condición de desarrollo SBC-M, se conforman por litosoles de lomerío en la UM-I, UM-II y UM-III, mientras que en la UM-IV, se tiene una transición edáfica hacia regosoles eutricos. Ambos tipos de suelo se caracterizan por su escaso desarrollo, procesos de latosolización y de erosión que los empobrece de nutrientes en época de lluvias. Se trata de suelos intolerantes a la erosión, hidrófobos y de relieve accidentado. La vocación de uso de estos suelos es eminentemente forestal y por ningún motivo se deben considerar para un uso distinto ya que originaría su degradación y la pérdida de su potencial biológico.

