











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En las regiones áridas y semiáridas, la infiltración puede causar la acumulación superficial del agua o el almacenamiento del agua en el suelo, por lo que puede modificar distintos procesos ecológicos (Eldridge et al., 2000; Coyne y Thompson, 2006). El agua que entra en el suelo y es almacenada en éste promueve la germinación de las semillas y el crecimiento de las plantas. Por el contrario, el agua que permanece en la superficie del suelo genera escorrentía superficial que causa erosión y pérdida de nutrientes (Eldridge et al., 2000; Coyne y Thompson, 2006). Por lo anterior, el estudio de los factores físicos y biológicos que alteran la inf iltración es necesario para entender el funcionamiento de las regiones áridas y semiáridas.
Uno de los factores biológicos que alteran la infiltración en los ecosistemas áridos y semiáridos son las biocostras. Las biocostras son comunidades de cianobacterias, líquenes y musgos que crecen en la superficie del suelo (Belnap y Lange, 2003; Castillo-Monroy y Maestre, 2011). Estas comunidades forman una capa entre la atmósfera y el suelo que influye en la distribución y abundancia de las plantas y los microorganismos, y participa en el ciclo de los nutrientes y la dinámica del agua (Belnap, 2006; Maestre et al., 2011; Chamizo et al., 2012b). El impacto de las biocostras en la infiltración es contradictorio porque puede ser positivo, negativo o neutro. Así, por ejemplo, las biocostras compuestas por cianobacterias y musgos incrementan la infiltración debido a que absorben cantidades de agua relativamente grandes (Eldridge et al., 1997; Eldridge, 2001; Belnap, 2006). Además, estas biocostras incrementan la rugosidad del suelo, por lo que disminuyen la velocidad del agua e incrementan el tiempo de contacto con el suelo (Belnap, 2006; Belnap et al., 2013). Por el contrario, las biocostras compuestas por cianobacterias y líquenes disminuyen la infiltración debido a que las vainas, el talo y las rizinas obstruyen los poros del suelo o el talo produce sustancias hidrófobas (Eldridge, 1993; Eldridge y Rosentreter, 1999; Gao et al., 2010; Rodríguez-Caballero et al., 2013). Por último, las biocostras no alteran la inf iltración debido a que la textura, porosidad, estabilidad de agregados y densidad aparente controlan la cantidad de agua que entra en el suelo (Quiñones-Vera et al., 2009; Gao et al., 2010; Castillo-Monroy y Maestre, 2011). El impacto de las biocostras en la infiltración ha sido poco estudiado en las regiones áridas y semiáridas de nuestro país. Hasta donde sabemos, solo un trabajo ha evaluado la infiltración en biocostras de pastizales de Durango (Quiñones-Vera et al., 2009). En estos pastizales, los suelos con biocostra no modificaron la infiltración en comparación con los suelos sin biocostra. Debido a la escasa información sobre el impacto de las biocostras en la infiltración, es necesario realizar más estudios para entender la importancia de estas comunidades de microorganismos en la dinámica del agua en las regiones áridas y semiáridas mexicanas.
El Valle de Zapotitlán de las Salinas es una región semiárida del centro de México, en donde las biocostras son abundantes en laderas de cerros, salineras, campos de cultivo, tierras malas y terrazas fluviales (Rivera-Aguilar et al., 2006). En las terrazas fluviales las biocostras crecen en mezquitales con dosel abierto, en donde existen amplias áreas desprovistas de vegetación (> 5m de longitud). Las biocostras en estas áreas cubren hasta 77% de la superficie del suelo, por lo que contribuyen a estabilizarlo y protegerlo del efecto erosivo de la lluvia. Sin embargo, se desconoce si las biocostras modifican la infiltración. En este trabajo se evaluó la infiltración en suelos con biocostras en las terrazas fluviales del Valle de Zapotitlán de las Salinas. Para cumplir este objetivo, se determinaron las propiedades físicas y químicas del suelo, la composición y cobertura de las biocostras, y la infiltración en suelos con y sin costra.
Materiales y Métodos
El estudio fue realizado en las terrazas fluviales del río El Salado en el Valle de Zapotitlán de las Salinas, Puebla, México (18º 20’ N, 97º 28’ O, 1550 m de altitud). El clima es seco semi-cálido, y la temperatura y precipitación promedio anuales son 21 ºC y 420 mm, respectivamente (Muñoz et al., 2009). Los suelos son Fluvisoles calcáricos con textura franco arcillosa. La vegetación dominante es mezquital de Prosopis laevigata mezclado con Parkinsonia praecox, Myrtillocactus geometrizans, Castela tortuosa, Opuntia puberula y Stenocereus stellatus (Osorio et al., 1996). Se seleccionaron dos terrazas fluviales donde se determinaron las características físicas y químicas del suelo, cobertura y composición de las biocostras, y la infiltración. Ambas terrazas tienen mezquital con dosel abierto, pero distinta textura del suelo. La primera terraza tiene una superficie de 20 ha y el suelo tiene una distribución de partículas de 36.6 ± 3.6% de arena, 38.8 ± 2.2% de limo y 24.6 ± 2.1% de arcilla (se denomina terraza fina, de aquí en adelante). La segunda terraza tiene una superficie de 13 ha y el suelo tiene 69.8 ± 0.5% de arena, 16.0 ± 0.3% de limo y 14.2 ± 0.3% de arcilla (terraza gruesa, de aquí en adelante). En ambas terrazas, el muestreo fue realizado en áreas desprovistas de vegetación en donde las biocostras tienen 60% de cobertura. Allí se seleccionaron sitios donde la cobertura de las costras fuera homogénea.
Para medir las propiedades físicas y químicas del suelo, en cada terraza se tomaron 7 pares de muestras, separadas cada una ca. 30 cm, en los primeros 10 cm de profundidad. Las muestras fueron tomadas justo a un lado de los sitios en donde se realizaron las pruebas de infiltración (ver más adelante). Cada par de muestras de suelo se mezcló para obtener 7 muestras compuestas. En el laboratorio, las muestras compuestas fueron secadas y usadas para evaluar: (1) textura con el método del hidrómetro (Bouyoucos, 1962), (2) densidad aparente con el método de la probeta (Beaver, 1963), (3) densidad real con el método del picnómetro, (4) estabilidad de agregados con el método modificado de Yoder (USDA, 1999), (5) materia orgánica con el método de oxidación con ácido crómico y sulfúrico (Walkley-Black, 1947), y (6) pH con potenciómetro digital y relación suelo-agua 1:2.5 (p/v).
Para estimar la cobertura y composición de las biocostras, en cada terraza se seleccionaron siete pares de sitios de 6 × 6 cm, separados cada uno ca. 30 cm, con costra intacta. El tamaño de estos sitios fue igual al área de contacto de los inf iltrómetros usados en campo (ver más adelante). En cada sitio se colocó una hoja de acetato de vinilo dividida en 36 cuadros de 1 cm2 para contar el número de cuadros ocupados por cianobacterias, musgos y líquenes. Con el número de cuadros ocupados y el número total de cuadros se calculó el porcentaje del área ocupado por cada grupo de organismos. Las biocostras fueron colectadas con una espátula después de medir la infiltración y transportadas al laboratorio en bolsas de papel para su identificación.
La infiltración fue medida en los siete pares de sitios de cada terraza, en donde previamente se estimó la cobertura y composición de las biocostras. En uno de los sitios, la costra fue removida de la superficie del suelo con una espátula, mientras que en el otro sitio la costra permaneció intacta. En ambos sitios se midió la sortividad y la infiltración estable con infiltrómetros de mini-disco (marca Decagon Devices) con tensión de succión de -0.5 cm en un sitio y -6.0 cm en el otro. La sortividad es la capacidad del suelo seco para absorber agua por capilaridad. La infiltración estable es cuando el flujo del agua es constante y depende de la capilaridad, gravedad, tamaño del disco y tensión de succión (Eldridge et al., 2000). Cuando la tensión de succión es de -0.5 cm, el agua fluye por macroporos (> 1000 μ), mientras que cuando es de -6.0 cm, fluye por mesoporos (10-1000 μ; Filgueira et al., 2006). Los infiltrómetros fueron colocados sobre una capa delgada de arena de sílice para incrementar el contacto con la costra (Eldridge et al., 2000). Las mediciones fueron hechas simultáneamente en ambos sitios durante 30 min. De 0-10 min, las mediciones fueron registradas cada 30 s, de 11-15 min, cada 60 s y de 16-30 min, cada 300 s. Para cada tensión de succión, la sortividad y la infiltración fueron calculadas con el método de Zhang (1997), de acuerdo con las especificaciones del fabricante de los infiltrómetros.
Las propiedades físicas y químicas del suelo, y la cobertura de cianobacterias, musgos y líquenes fueron comparadas entre la terraza fina y gruesa. Además, en cada terraza la sortividad e infiltración con tensiones de succión de -0.5 y -6.0 cm fueron comparadas entre sitios con y sin costra. Las comparaciones fueron hechas con pruebas de t de Student, en el programa estadístico SPSS, versión 15.0.
Los datos expresados en porcentaje como algunas propiedades del suelo y la cobertura de las costras fueron transformados con la función arcoseno para cumplir los supuestos de normalidad e igualdad de varianzas. Sin embargo, cuando los datos transformados no cumplieron estos supuestos, los valores de t, grados de libertad y significancia reportados fueron calculados por el programa para varianzas desiguales.
Resultados y Discusión
Los suelos de las terrazas fina (42%) y gruesa (39%) tuvieron altos porcentajes de limo (Cuadro 1). La estabilidad de los agregados fue baja (18-19%) y el contenido de materia orgánica moderadamente pobre (1.4-1.5%) en ambas terrazas. La densidad real fue ca. 2.5 g cm-3. Sin embargo, la densidad aparente fue alta en la terraza fina (1.3 g cm-3) y media en la gruesa (1.2 g cm‑3). La porosidad fue media en la terraza fina (47%) y ligeramente alta en la gruesa (50%). El pH fue menor en la terraza fina (7.8) que en la gruesa (7.9), pero ambos fueron ligeramente alcalinos. Los altos porcentajes de limo de las terrazas fina y gruesa concuerdan con los encontrados en otras terrazas fluviales del Valle de Zapotitlán de las Salinas (Muñoz et al., 2009). En las terrazas fluviales de este valle, las áreas desprovistas de vegetación tienen altos porcentajes de limo debido a que estas partículas son transportadas y depositadas en estos sitios por escurrimiento superficial (Muñoz et al., 2009). A pesar de que la terraza fina y gruesa tuvieron altos porcentajes de limo, el suelo de la terraza fina fue más compacto y tuvo menor espacio poroso que el suelo de la terraza gruesa.
Cuadro 1: Propiedades físicas y químicas (promedio ± error estándar) del suelo en la terraza fina y gruesa.
Propiedad |
Terraza |
|
Fina |
Gruesa |
|
Arenas (%) |
36.97 ± 4.25 |
35.37 ± 1.55 |
Limos (%) |
41.66 ± 2.92 |
39.57 ± 0.54 |
Arcillas (%) |
21.37 ± 1.59 |
25.06 ± 1.65 |
Densidad aparente (g cm-3) |
1.31 ± 0.02 a |
1.24 ± 0.01 b |
Densidad real (g cm-3) |
2.49 ± 0.02 |
2.51 ± 0.02 |
Porosidad (%) |
47.32 ± 0.76 a |
50.39 ± 0.35 b |
Estabilidad de agregados (%) |
18.54 ± 2.01 |
17.84 ± 1.20 |
Materia orgánica (%) |
1.37 ± 0.10 |
1.47 ± 0.14 |
pH |
7.78 ± 0.04 a |
7.92 ± 0.02 b |
Las letras en las propiedades del suelo indican diferencias significativas entre las terrazas (P < 0.05).
Las biocostras de ambas terrazas estuvieron compuestas por cinco géneros de cianobacterias y tres de líquenes. No tuvieron ningún musgo. Las cianobacterias fueron Chroococcidiopsis sp., Chroococcus sp., Nostoc sp., Schyzothrix sp. y Scytonema javanicum. Su cobertura fue similar en la terraza f ina (24 ± 0.046%, promedio ± error estándar, N = 14) y gruesa (14 ± 0.035%, N = 13; t = 1.9; g. l. = 25, p = 0.07). Los líquenes fueron Enchylium sp., Endocarpon sp., Placidium squamulosum, morfo especie 1 y morfo especie 2. Su cobertura fue menor en la terraza fina (65 ± 0.045%, N = 14) que en la gruesa (78 ± 0.043%, N = 14; t = 2.6, g. l. = 26, p = 0.02). Los géneros de cianobacterias y líquenes en las biocostras de las terrazas son similares a los géneros encontrados en las biocostras de otros sitios como laderas de cerros, salineras, campos de cultivo y tierras malas en el Valle de Zapotitlán de las Salinas (Rivera-Aguilar et al., 2006, 2009). Además, la presencia de los líquenes en las biocostras de las terrazas concuerda con la observación de que los líquenes crecen en sitios con baja cobertura vegetal o completamente expuestos a la radiación solar (Belnap y Lange, 2003; Rivera-Aguilar et al., 2006; Castillo-Monroy y Maestre, 2011). La alta cobertura de los líquenes en las terrazas podría estar relacionada con la densidad aparente del suelo (1.2-1.3 g cm-3) y el pH (7.8-7.9). En este sentido, Rivera-Aguilar et al. (2009)también encontraron que los líquenes tienen alta frecuencia relativa en suelos con densidad aparente de 1.2 g cm-3 y pH 8. Por último, la ausencia de musgos en las biocostras de las terrazas podría deberse a que las áreas desprovistas de vegetación no tienen las condiciones adecuadas de sombra y humedad para el crecimiento de estos organismos (Belnap y Lange, 2003).
La sortividad e infiltración estable fueron diferentes en cada terraza (Figura 1). En la terraza fina, la sortividad (t = 2.4, g. l. = 8, p = 0.04) e infiltración (t = 3.1, g. l. = 7, p = 0.02) con tensión de -0.5 cm fueron menores en los sitios con costra que sin costra. La sortividad (t = 0.9, g. l. = 12, p = 0.4) e infiltración (t = 2.4, g. l. = 7, p = 0.05) con tensión de -6.0 cm fueron similares en sitios con y sin costra. En la terraza gruesa, la sortividad con tensión de -0.5 cm fue mayor en sitios con costra que sin costra, aunque estas diferencias fueron marginalmente no significativas (t = 2.1, g. l. = 12, p = 0.06). La infiltración con tensión de -0.5 cm fue mayor en sitios con costra que sin costra (t = 5.6, g. l. = 12, p = 0.0001). La sortividad (t = 1.2, g.l. = 12, p = 0.3) e infiltración (t = 0.6, g. l. = 1, p = 0.7) con tensión de -6.0 cm fueron similares en sitios con y sin costra. Las diferencias en la sortividad e infiltración entre las terrazas sugieren que las biocostras alteran el movimiento del agua dependiendo de las propiedades físicas del suelo. Sin embargo, la infiltración solo fue evaluada en las áreas desprovistas de vegetación de las terrazas, por lo que es necesario evaluarla en áreas con plantas perennes y en otros sitios como laderas para entender mejor el impacto de las costras en la infiltración en esta región.
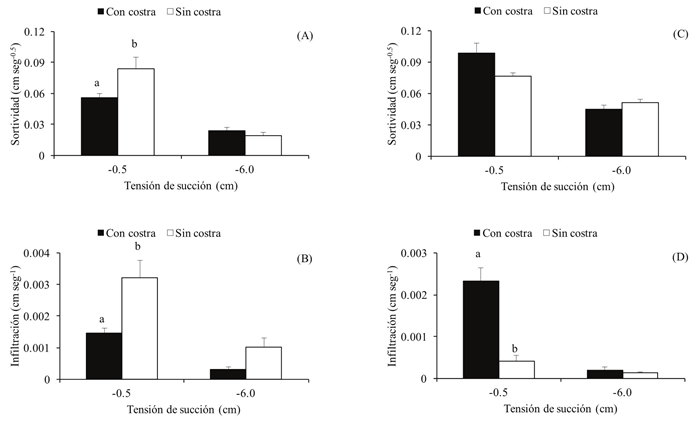
Figura 1: Sortividad e infiltración (promedio ± error estándar) con tensiones de succión de -0.5 cm y -6.0 cm, en suelos con y sin biocostra en las terrazas fina (A y B) y gruesa (C y D). En cada panel, las letras indican diferencias significativas entre los suelos con y sin costra (P < 0.05)
Las biocostras en ambas terrazas estuvieron compuestas por líquenes y cianobacterias. Algunos autores han sugerido que los líquenes son hidrófobos por lo que pueden impedir la inf iltración (Eldridge, 1993; Eldridge y Rosentreter, 1999; Rodríguez-Caballero et al., 2013; Chamizo et al., 2016). Sin embargo, nuestros resultados no apoyan esta observación debido a que la infiltración fue registrada en ambas terrazas. La infiltración inclusive fue mayor en la terraza gruesa, en donde los líquenes tuvieron el mayor porcentaje de cobertura. Belnap et al. (2013) sugieren que las costras compuestas en su mayoría por líquenes y musgos tienen mayor volumen y profundidad de infiltración que las costras incipientes de cianobacterias y los suelos sin costra. Por su parte, las cianobacterias absorben cantidades de agua relativamente grandes (Eldridge et al., 1997; Eldridge, 2001; Belnap, 2006; Belnap et al., 2013), por lo que incrementan el tiempo de contacto del agua con el suelo e incrementan su infiltración.
El suelo en la terraza fina tuvo alto porcentaje de limo, alta densidad aparente y baja porosidad, por lo que el agua absorbida por las biocostras pudo disgregar los limos, los cuales taparon los poros del suelo y disminuyeron la infiltración. Los limos son partículas pequeñas, sin carga, que son fácilmente transportadas por el agua. Estas partículas obstruyen los poros y sellan la superficie del suelo (Muñoz et al., 2009). La infiltración también pudo disminuir por los filamentos de las cianobacterias y las rizinas de los líquenes que podrían obstruir los poros del suelo. Por el contrario, el suelo en la terraza gruesa tuvo menor porcentaje de limo, menor densidad aparente y mayor porosidad por lo que los limos probablemente no taparon los poros y el agua absorbida por las costras biológicas pudo entrar en el suelo. La modificación de la infiltración por las biocostras en ambas terrazas fue mayor cuando el agua fluyó por macroporos (tensión de succión de -0.5 cm) que por mesoporos (tensión de succión de -6.0 cm). Los resultados del presente trabajo coinciden con los reportados para el desierto de Negev, Israel, en donde la infiltración en suelos limosos fue menor en sitios con costra que en sitios sin costra (Eldridge et al., 2000). Además, son similares a los reportados para una región semiárida en Almería, España, donde la infiltración en suelos de textura fina fue similar en sitios con costra de líquenes y sitios con costra física, mientras que en suelos de textura gruesa fue mayor en sitios con costra de líquenes que en sitios con costra física (Chamizo et al., 2012a). Por el contrario, nuestros resultados difieren de otros reportados para pastizales en Durango, México, en donde la infiltración fue similar en sitios con y sin costra (Quiñones-Vera et al., 2009). Sin embargo, las comparaciones con estos trabajos deben interpretarse cuidadosamente debido a que existen diferencias en los métodos y en el volumen de agua usados para medir la infiltración, lo cual altera la capacidad de infiltración de las biocostras.
Conclusiones
Las biocostras modificaron la infiltración dependiendo del contenido de limo, la densidad aparente y porosidad del suelo de las terrazas fluviales. La infiltración fue menor en los suelos de la terraza fina, los cuales tuvieron alto porcentaje de limo, mayor densidad y menor porosidad que los suelos de la terraza gruesa. Estos resultados muestran que las biocostras modifican la infiltración en las terrazas fluviales del Valle de Zapotitlán de las Salinas. Es necesario medir la infiltración en otros sitios como áreas debajo de plantas perennes y laderas de cerros para entender el impacto de las costras en la infiltración en esta región semiárida.

