











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El cambio climático global asociado al aumento potencial de la temperatura superficial del planeta es un problema que se acentúa por el rápido incremento actual en las emisiones de Gases de Efecto Invernadero (GEI) (Bolin et al., 1989) y por las dificultades de reducir en forma sustantiva el incremento de los mismos en el futuro próximo (Houghton et al., 1995); así, por ejemplo, se calcula un ingreso antropogénico a la atmósfera de aproximadamente 8.9 Pg C año-1 a nivel mundial (Ciais et al., 2013). Diferentes autores: Schneider (1989), Houghton y Woodwell (1989), Lashof y Ahuja (1990), Dixon et al. (1994), Masera (1995), Schimel (1995), (Le Quéré et al., 2009), Houghton et al. (2012), afirman que el dióxido de carbono (CO2) es uno de los gases de efecto invernadero (GEI) más importantes y que su emisión a la atmósfera por el cambio de uso del suelo ocupa el segundo lugar a nivel mundial que, de acuerdo a las últimas estimaciones disponibles, se calcula 1.1-1.14 Pg C año-1 (Ciais et al., 2013, Houghton et al., 2012), con una fuerte contribución de las zonas tropicales.
En México, de acuerdo al Inventario Nacional de emisiones de GEI, la categoría Uso del Suelo, Cambio de Uso del Suelo y Silvicultura (USCUSS) fue en 2006 el tercero en liberación de GEI a la atmósfera con un 9.9% del total estimado en el país (SEMARNAT e INECC, 2009); aunque en 2010 se estimó en 6.3%, siendo el cuarto en importancia (SEMARNAT e INECC, 2012), lo que representó una disminución en términos absolutos de 23 310 Gg de CO2 eq, debidos principalmente a cambios en la metodología e interpretación, dado que se utilizó la misma información base (SEMARNAT e INECC, 2012). Debido a lo anterior, los ecosistemas forestales tienen un papel prioritario en el ciclo global del carbono y en el cambio climático global, por lo que resulta de gran importancia su conservación; ya que a pesar de ello, no existen mediciones forestales colectadas en forma regular y sistemática utilizando metodologías consistentes a lo largo del tiempo y manteniendo series históricas, debido a lo cual existe un alto grado de incertidumbre en estas estimaciones (De Jong et al., 2009).
De acuerdo al Grupo Intergubernamental de Expertos sobre el Cambio Climático (IPCC, 2003 y 2006), en los ecosistemas forestales el C se distribuye en cinco depósitos: biomasa viva por encima del suelo (biomasa aérea), biomasa viva por debajo del suelo (biomasa subterránea), madera muerta sobre el suelo, mantillo y carbono orgánico del suelo (COS). La biomasa aérea y subterránea se definen como la cantidad total de material orgánico vivo existente, por arriba y por abajo del suelo, respectivamente, expresada como peso anhidro por unidad de área (FAO, 2004). La capacidad de los ecosistemas forestales para almacenar carbono en forma de biomasa varía en función de la composición florística, la edad y la densidad de población de cada estrato por comunidad vegetal (Schulze et al., 2000). Así, Prentice et al. (2001), estimaron que la mitad de la producción primaria bruta (PPB) se incorpora en los tejidos vegetales como hojas, raíces y tejidos leñosos (producción primaria neta o PPN), calculando globalmente con datos de los ochentas, que la PPB era de 120 Pg año-1 de C. Posteriormente, Beer et al. (2010) estimaron la PPB en 123±8 Pg año-1.
El COS es el producto de la descomposición de plantas, el metabolismo de organismos vivos y el crecimiento bacteriano (Gardi et al., 2014). Se estima que el depósito de COS a 1 m de profundidad es de 1500-2400 Pg (Batjes, 1996; Lal, 2003; Ciais et al., 2013), tres a cuatro veces mayor que el depósito de C en la vegetación (550 Pg C) y dos veces mayor que el atmosférico (760 Pg) (Eswaran et al., 1993; Lal, 2003).
El mantillo incluye desde materia orgánica fresca hasta materiales orgánicos altamente descompuestos y parcialmente estabilizados y es la fuente principal de C y de nutrientes disponibles para las plantas (Swift et al., 1979; Caldato et al., 2010), teniendo en gran medida responsabilidad en el funcionamiento de los ecosistemas forestales. La calidad del mantillo (relación carbono: nitrógeno) está determinada en gran medida por las especies de árboles presentes, que a su vez dependen de la elevación, el clima y la historia de uso (Berg, 2000).
De acuerdo al IPCC (2006), la madera muerta comprende toda la biomasa boscosa no viva no contenida en el mantillo, ya sea en pie, superficial o en el suelo. Es un componente natural importante en los bosques, que además de ser un elemento clave de la biodiversidad forestal, también influencia el microclima de éstos, actúa como un almacén de agua en períodos secos (Harmon et al., 1986) y como un almacén de C y nutrientes a largo plazo (Harmon et al., 1986; Schaetzl et al., 1989; Keenan et al., 1993).
El Monte Tláloc forma parte de la Sierra Nevada, una de las regiones terrestres prioritarias para la Comisión Nacional de Áreas Naturales Protegidas (CONANP), y representa un área de gran importancia desde el punto de vista biogeográfico puesto que ahí convergen las regiones neártica y neotropical, además de valores altos de riqueza y endemismos (Arriaga, et al., 2000) y como principal zona de recarga y amortiguamiento hidrológico del Valle de México (Moctezuma y Carreón, 2005). Adicionalmente, la Sierra Nevada se sitúa al oriente de uno de los mayores complejos urbanos del mundo: la zona metropolitana de la Ciudad de México, la cual ejerce una enorme presión sobre la vegetación circundante y en general sobre los recursos naturales (Sánchez y López, 2003), misma que, se prevé, se incremente con la construcción del Nuevo Aeropuerto Internacional de la Ciudad de México (OECD, 2015), en especial en el municipio de Texcoco, Estado de México. Debido a esta importancia geográfica estratégica, a la falta de información específica de factores de emisión del sector agrícola, silvícola y otros usos del suelo (AFOLU, por sus siglas en inglés) y a la capacidad de almacenamiento de carbono de los ecosistemas forestales, descrita anteriormente; en el presente estudio se seleccionaron dos sitios de muestreo ubicados en el municipio de Texcoco, utilizando metodologías de muestreo y laboratorio estandarizadas con los criterios definidos por el IPCC (IPCC, 2003 y 2006) para la estimación de los depósitos de carbono de un bosque de oyamel y un bosque de ciprés; aportando información para reducir el grado de incertidumbre en las estimaciones de emisiones de GEI que a la fecha se tienen en este sector, por lo que este trabajo contribuye con este objetivo.
Materiales y Métodos
Sitios de muestreo
El área de muestreo se ubicó entre las coordenadas 19° 26’ 42.36” y 19° 28’ 20.09” N y 98° 44’ 59.74” y 98° 46’ 42.78” O e integra parte de los ejidos de Santa Catarina del Monte, San Pablo Ixayoc, Santa María Nativitas y San Miguel Tlaixpan, todos en el municipio de Texcoco. Se seleccionó un paisaje de 9 km2 espacialmente representativo de la vegetación y de los tipos de suelos de los bosques templados del oriente del Estado de México, dentro de esta área se delimitaron dos zonas denominadas Bosques de Referencia (BR), con rasgos homogéneos en términos de almacenes de carbono: el primero fue un bosque monoespecíf ico de oyamel (Abies religiosa) y el segundo un bosque monoespecífico de ciprés (Cupressus lusitanica). Cada BR (Conglomerado) se integró por cuatro sitios de muestreo (Figura 1). Algunas de las variables de densidad de ambos BR se muestran en el Cuadro 1, en el que se agregó como referencia el número de árboles muertos en pie y tocones medidos en ellos. Este último dato muestra que ambos BR han estado sujetos a intervención humana, dado que en la mayoría de sitios que conforman cada BR el número de tocones encontrados fue superior al de árboles vivos. Destaca también el hecho de que el área basal de los sitios 3 y 4 del BR1 es mayor que la de los sitios 1 y 2 del mismo BR a pesar de tener una cantidad similar de individuos.

Fuente: Elaboración propia, adaptado de CONAFOR 2011.
Figura 1: Diseño del conglomerado (Bosque de Referencia).

† NAV = número de árboles vivos de la especie dominante con DN ≥ 7.5cm. ‡ NAMP = número de árboles muertos en pie.
Cuadro 1: Variables de densidad de los bosques de referencia.
Respecto a las clases diamétricas, el BR1 (oyamel) corresponde a una masa forestal irregular, con un rango de clases muy amplio. En el BR2 (ciprés) los individuos se concentran en las clases más pequeñas (10-50 cm), por lo que a pesar de tener una mayor cantidad de individuos con respecto a los sitios 3 y 4 del BR1, su área basal es sensiblemente inferior a éstos (Cuadro 2).
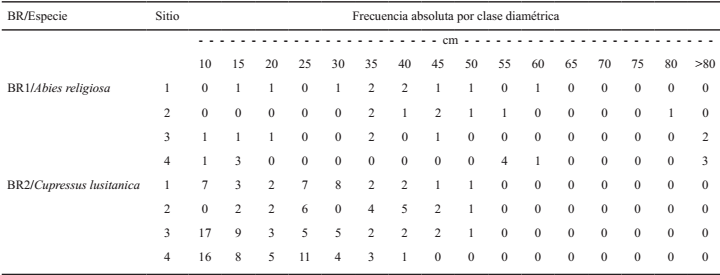
Diseño del muestreo de campo
El diseño del muestreo de campo se orientó para obtener datos cuantitativos de COS, mantillo, madera muerta, biomasa área y biomasa subterránea. Los sitios cuantitativos se distribuyeron en los BR formando un conglomerado tipo CONAFOR integrado por cuatro sitios de muestreo circulares de 400m2 de área en forma de “Y” invertida (CONAFOR, 2011) (Figura 1).
En cada uno de los cuatro sitios cuantitativos del conglomerado se midieron todos los árboles con diámetro normal (DN) ≥ 7.5cm, los arbustos y se tomaron cuatro muestras de biomasa de herbáceas (0.25 m2), mismas que se colectaron en áreas representativas de cada sitio. Se colectaron cinco muestras de mantillo (hojarasca y capa de fermentación) y suelo, cuatro de ellas distribuidas en la periferia del círculo que delimitó el sitio, orientadas a cada uno de los puntos cardinales, y una en el centro del mismo, denominada DAP (donde se midió densidad aparente del suelo). Finalmente, se trazaron dos transectos perpendiculares entre sí para medir el material leñoso caído (combustibles), tal como se muestra en la Figura 2.
Estimación de depósitos de carbono
Los depósitos considerados fueron los siguientes:
Biomasa aérea. Se refiere a toda la biomasa viva por encima del suelo incluyendo el fuste, las ramas, la corteza, las semillas y las hojas; su cuantificación es de gran importancia debido a que aproximadamente el 50 % de ella es C (Brown, 1997; Chojnacky y Milton, 2008). En el caso de la vegetación arbórea se midieron los datos de diámetro normal (DN) de cada árbol registrado por sitio de muestreo. Con estos datos se realizó el cálculo de biomasa para Abies religiosa utilizando ecuaciones alométricas y factores de expansión de biomasa encontrados en la literatura revisada (Cuadro 3).

† FEB = factores de expansión de biomasa.
Cuadro 3: Métodos/modelos para estimación de biomasa de Abies religiosa.
Basado en los resultados obtenidos en el cálculo de biomasa de Abies religiosa, para cuantificar el contenido de carbono se seleccionaron dos ecuaciones alométricas para esta especie, tomando como criterio de selección el que se explica en la sección denominada Estimación de contenido de biomasa de Abies religiosa. Adicionalmente se utilizó una ecuación para el cálculo de biomasa y carbono de Cupressus lusitanica y otra para Quercus laurina (se encontraron 2 árboles de esta especie en el BR2 y uno en el BR1), las cuales corresponden a las especies arbóreas encontradas en los sitios de muestreo (Cuadro 4), tomando como criterios de selección el que corresponden a la misma especie y que fueron obtenidas en sitios próximos geográficamente al área de estudio.

Cuadro 4: Ecuaciones alométricas utilizadas para la estimación de contenido de carbono en biomasa aérea arbórea.
En lo que respecta a la vegetación arbustiva se realizó un procedimiento similar al usado para la vegetación arbórea, pero no se encontraron en la literatura ecuaciones alométricas desarrolladas para las especies encontradas en los sitios de muestreo, por lo que se utilizó la ecuación desarrollada por Búrquez et al. (2010), ya que, aunque fue desarrollada para especies de zonas desérticas (la gran mayoría de ecuaciones encontradas fueron desarrolladas para especies de éstas zonas, donde el aporte del estrato arbustivo a la biomasa total del sitio es muy importante) mostró consistencia en las estimaciones de biomasa de muchas especies con diferentes formas de crecimiento, tamaños, condiciones edáficas y de disponibilidad de humedad.
Así, el modelo utilizado considera el área proyectada de la copa como una elipse, de tal forma que el volumen de ésta corresponde a un cilindro elíptico, Ecuación 1.
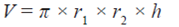 (1)
(1)
De los tres modelos desarrollados por Búrquez et al. (2010), en este trabajo se seleccionó el no lineal, mismo que se generó en condiciones de mayor disponibilidad de agua en el suelo y estima la cantidad de biomasa seca sobre el mismo (Bm, g m-2) a partir del volumen de copa (V, en m3 m-2) con la Ecuación 2.
 (2)
(2)
El contenido de carbono en el estrato herbáceo se calculó a partir del peso seco de la muestra (1 m2) y se obtuvo la densidad de la biomasa (Mg ha-1). Posteriormente el valor de densidad de biomasa se multiplicó por el factor de contenido de carbono recomendado por el IPCC (2003) de 0.5 para obtener la densidad de carbono en este estrato.
Biomasa subterránea. Este reservorio incluye toda la biomasa de raíces vivas excluyendo raíces finas de menos de 2 mm de diámetro debido a que con frecuencia no se pueden distinguir de la materia orgánica del suelo o mantillo (IPCC, 2006). En el presente estudio este depósito se estimó únicamente para vegetación arbórea viva, ya que se consideró que la de la vegetación herbácea es poco significativa y la de la vegetación arbustiva requiere realizar muestreos destructivos, que implican una gran cantidad de tiempo y recursos económicos (no son aplicables las ecuaciones alométricas existentes en la literatura), además de provocar una perturbación mayor en los sitios de medición, por lo que se optó por no considerarlos en la cuantificación de éste depósito. Basado en el trabajo realizado por Cairns et al. (1997), el IPCC (2006) recomienda la utilización de la ecuación alométrica (Ecuación 3) para estimar la biomasa por debajo del suelo en bosques templados, en función de la biomasa aérea seca (BSS) calculada previamente.
 (3)
(3)
Carbono orgánico del suelo (COS). El COS se estimó en función de la densidad aparente, profundidad, fracción gruesa del suelo (partículas mayores a 2mm) y porcentaje de carbono medido con un analizador automático Shimadzu 5050A. Para obtener el valor del COS se utilizó la Ecuación 4.
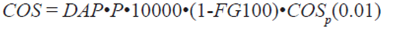 (4)
(4)
dónde:
COS = carbono orgánico del suelo (Mg ha-1)
DAP = densidad aparente del suelo (Mg m-3)
P = profundidad del suelo (m)
FG = fragmentos gruesos en el suelo (> 2 mm), en %
COSp = contenido de carbono orgánico, en % 0.01 factor de conversión de unidades (porcentaje a fracción).
El valor del factor de escala (10 000) tiene como unidades m2 ha-1.
El valor de la DAP se obtuvo tomando una muestra de suelo en campo, misma que se llevó al laboratorio para secado en estufa y determinar su peso seco. La profundidad que se consideró en el cálculo del contenido de carbono para el presente estudio fue de 0.3 m debido a que, de acuerdo al IPCC (2003), es una buena práctica medir el depósito de carbono en el suelo mínimamente a esta profundidad, ya que probablemente donde se producirán las mayores variaciones es en este depósito en el tiempo. El porcentaje de FG se obtuvo secando, moliendo y tamizando el suelo de las muestras en una malla 10, con abertura de 2 mm, calculando el peso de ambas fracciones (gruesa y fina) y su respectivo porcentaje en relación al total de la muestra. Posteriormente se obtuvo una submuestra de 5 g, la cual se tamizó en malla 100 para estimar el COSp.
Mantillo. Incluyó la hojarasca capa de fermentación y material leñoso caído fino con diámetro menor de 7.5 cm. El cálculo de contenido de C se realizó determinando el peso seco de cada compartimento y midiendo el porcentaje de carbono de las muestras mediante el analizador automático Shimadzu 5050A.
Madera muerta. Dentro de este depósito se consideró árboles muertos en pie, tocones, trozas o material leñoso caído con un diámetro ≥ 7.5 cm. Para los árboles se estimó el contenido de carbono utilizando las mismas ecuaciones alométricas que para biomasa aérea, para los tocones se estimó su volumen con el diámetro y altura medido (asumiendo que tienen forma cilíndrica) y la densidad de madera de acuerdo a su grado de putrefacción y para el caso del material leñoso caído (MLC) se utilizó el método de líneas de intersección recomendado por el IPCC (Van Wagner, 1968; IPCC, 2006), Ecuación 5, y la densidad de madera de acuerdo a su grado de putrefacción.
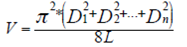 (5)
(5)
dónde:
V = volumen de MLC (m3 ha-1)
D = diámetro de cada una de las n trozas (cm)
L = largo de la línea (suma de todos los transectos) en sitio de muestreo (m)
Posteriormente se estimó la biomasa, para ello se utilizó la Ecuación 6.
 (6)
(6)
dónde:
Resultados y Discusión
Estimación de contenido de biomasa de Abies religiosa
En el BR1 se encontraron árboles de Abies religiosa con Diámetro Normal (DN) >70 cm, de grandes dimensiones de acuerdo a la definición de Brown et al. (1997), mismos que estaban fuera del rango de estimación de todas las ecuaciones alométricas encontradas inicialmente para esta especie en la literatura revisada (Avendaño et al., 2009; Rodríguez y Calva, 2013; Flores et al., 2011), por lo que las estimaciones de biomasa y contenido de carbono con las ecuaciones mencionadas resultaron ser mucho mayores a todo lo reportado en la literatura para esta especie. Por lo anterior, se utilizó el método de Factores de Expansión de Biomasa (FEB), que es uno de los dos que recomienda el IPCC (2003), y la ecuación alométrica propuesta por Brown et al. (1997) utilizada en la cuantificación de contenido de carbono en bosques inalterados del este de Estados Unidos, en donde se encontraron árboles de grandes dimensiones, similares a los encontrados en el BR1. En los sitios 1 y 2, donde todos los árboles medidos tuvieron un DN<70 cm, la cantidad de biomasa estimada con todos los métodos utilizados es relativamente similar, dado que el coeficiente de variación (CV) para el sitio 1 es de 32.6% y 31.5% para el sitio 2 (Cuadro 5), siendo la ecuación que calcula el valor menor la propuesta por Flores et al. (2011) y el mayor valor al utilizar el método de factores de expansión de biomasa (IPCC, 2003). Las otras tres ecuaciones alométricas propuestas (Brown et al., 1997; Avendaño et al., 2009; Rodríguez y Calva, 2013) estiman valores muy similares de biomasa, por lo que, tomando en cuenta únicamente éstas ecuaciones, el CV se reduce a 4.32% para el sitio 1 y 1.91% para el sitio 2 (Cuadro 5).

† = desviación estándar; ‡ = coeficiente de variación.
Cuadro 5 Biomasa aérea de Abies religiosa calculada con diferentes métodos/ecuaciones alométricas.
En cambio, en los sitios 3 y 4 del BR1, donde se midieron los árboles con DN>80 cm, existió una diferencia notable entre las estimaciones con los métodos utilizados. Así, con excepción de la ecuación alométrica propuesta por Brown et al. (1997), el resto calculó cantidades superiores a lo reportado en la literatura revisada para este tipo de vegetación y especie (De Jong et al., 1999; Ordóñez et al., 2008; Avendaño et al., 2009; Rodríguez y Calva, 2013; Flores et al., 2011). Lo anterior se debe a que los DN de los árboles de grandes dimensiones están fuera de los rangos utilizados en la construcción de estas ecuaciones alométricas, por lo que no consideran esta condición. Así, por ejemplo, Avendaño et al. (2009) utilizaron rangos de DN entre 5.7 y 79 cm; pero en el BR1 se midieron tres árboles en el Sitio 4 con DN>79cm (84, 159 y 262 cm) y en el Sitio 3 dos árboles (132 y 210 cm), por lo que el modelo utilizado no es representativo de estas clases diamétricas y, aunque son solo cinco especímenes de un total de 38 medidos, dadas sus dimensiones, es suficiente para sobreestimar el contenido de carbono en estos sitios.
Adicionalmente, en la Figura 3 se muestra una comparación de los valores de biomasa por árbol estimada con las ecuaciones alométricas propuestas por Avendaño et al. (2009) y Brown et al. (1997), donde se observó que para árboles de Abies religiosa con DN<80 cm de los cuatro sitios que integran el conglomerado (BR) los valores estimados son muy similares con ambas ecuaciones. En contraste, para árboles de esta misma especie, pero con DN>80 cm, se observó una diferencia amplia en las estimaciones, misma que es mayor a medida que crece el DN (Figura 4), lo cual se debe principalmente a que la ecuación propuesta por Avendaño et al. (2009) es de tipo exponencial, por lo que los valores estimados crecen rápidamente conforme se incrementa el valor de la variable independiente (DN).
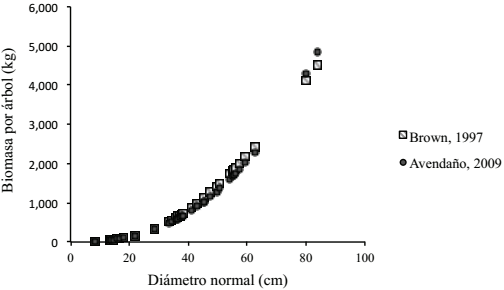
Figura 3: Biomasa aérea arbórea de Abies religiosa, utilizando las ecuaciones alométricas propuestas por Avendaño et al. (2009) y Brown et al. (1997), especímenes con DN < 85cm.
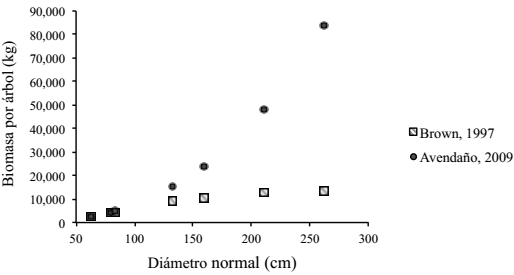
Figura 4: Biomasa aérea arbórea de Abies religiosa, utilizando las ecuaciones alométricas propuestas por Avendaño et al. (2009) y Brown et al. (1997), especímenes con DN > 60cm.
Como resultado del análisis anterior, para estimar el contenido de C en el BR1 se utilizó la ecuación alométrica propuesta por Avendaño et al. (2009) para árboles de A. religiosa con DN ≤ 80cm, considerada más precisa debido a que es específica para esta especie y se generó a partir de un sitio geográficamente próximo y con condiciones similares al del presente estudio; y la propuesta por Brown et al. (1997) para los especímenes con DN superior a este valor: dos en el caso de sitio 3 y tres en el caso del sitio 4, como se mencionó anteriormente.
Cálculo de contenido de carbono
La densidad de C total fue mayor en el BR1 con respecto al BR2, siendo superior en todos los depósitos considerados con excepción del COS. Así, el promedio de la densidad de carbono total para los sitios del BR 1 fue de 376 Mg ha-1, siendo el depósito más importante el de la biomasa aérea (59%), seguido del COS (22%); en tanto que en el BR 2 fue de 205 Mg ha-1, siendo el depósito con mayor densidad el COS (50%), seguido de biomasa aérea (36%). En el Cuadro 6 se muestran las densidades de carbono para los cinco depósitos considerados en el presente estudio, así como los promedios para cada uno de los BR medidos.

Para el caso de densidad de carbono en biomasa aérea, el contenido mayor en el BR1 con respecto al BR2 se debe principalmente a que las ecuaciones alométricas utilizadas están en función de la especie considerando como variable independiente el DN de los individuos, por lo que al ser mayores los DN de los especímenes de Abies religiosa medidos en el BR1, a pesar de ser menos individuos, con respecto a los especímenes de Cupressus lusitanica medidos en el BR2, la biomasa aérea es mayor en el primero.
En el BR1, el valor calculado de C en biomasa aérea fue superior al reportado en otros trabajos con tipos de vegetación similares, por ejemplo De Jong et al. (1999) reportó densidades de carbono en biomasa de 145.7 Mg C ha-1; Ordóñez et al. (2008) 169.7 Mg C ha‑1; Galeana et al. (2013) 163 Mg C ha‑1 (bosque cerrado); incluyendo a un bosque mono específico de A. religiosa de 75 años de edad ubicado en el Cofre de Perote, Veracruz en el que se estimó una densidad de C en biomasa aérea arbórea de 157 Mg C ha‑1 (Mendoza y Galicia, 2010). Lo anterior se debe, como ya se mencionó, a que se encontraron árboles de grandes dimensiones en dos de los sitios de muestreo que integraron el BR1. En contraste, las densidades medias de carbono estimadas en el BR2 para biomasa (73 Mg ha‑1) son inferiores a las reportadas por otros autores en tipos de vegetación similares (bosques de pino-encino), por ejemplo, De Jong et al., 1999, reportó densidades de carbono en biomasa de 166.3 Mg ha-1 y Ordóñez et al. (2008) 115.7 Mg ha‑1), debido principalmente a que este bosque tuvo especímenes con DN pequeño (10‑40 cm) y a que ha estado sujeto a tala, debido a la gran cantidad de tocones que se midieron, como se reportó en el Cuadro 1.
La densidad del COS de ambos BR se encuentra por debajo de lo reportado por otros autores para vegetación similar en México, tales como De Jong et al. (1999) quienes encontraron densidades de 172.6 Mg C ha‑1 (Bosque de pino) y 174.4 Mg C ha-1 (bosque de pino-encino) en los altos de Chiapas; Cairns et al. (1997) reportan una densidad promedio de 118 Mg C ha-1 para bosques de México. El contenido de C en este depósito fue mayor en el BR2 que en el BR1, lo que se debe principalmente a que el porcentaje de la fracción gruesa del BR1 es mayor al del BR2, por lo que esta fracción se resta de la masa del suelo, de acuerdo a la Ecuación 4.
En el caso del contenido de C en Mantillo, los valores encontrados (11 y 5 Mg ha-1) son inferiores al reportado por Woodall (2012), quién encontró que la densidad promedio de C en este depósito en los ecosistemas forestales de Estados Unidos fue de 25.6 Mg ha‑1, lo cual puede deberse a que los sitios medidos en este estudio están sujetos a intervención humana, por lo que es probable que parte del mantillo se haya retirado para utilizarse como combustible (extracción de leña) o materia orgánica para plantas (tierra de monte), prácticas comunes en la zona; sin embargo, esto no fue cuantificado.
Conclusiones
- Los resultados obtenidos en el presente estudio mostraron que los depósitos de carbono mayores fueron la biomasa viva sobre el suelo (biomasa aérea) y el carbono orgánico del suelo (COS). El primero fue más importante en el bosque de Abies religiosa con 237 Mg ha-1 (representó el 59% del total de C almacenado) y el segundo fue el mayor depósito en el bosque de Cupressus lusitanica con 102 Mg ha-1 (representó el 50% del C total estimado). En ambos bosques la suma de estos dos depósitos fue superior al 80% del total del C almacenado. Asimismo, dentro del depósito correspondiente a la biomasa aérea, el estrato arbóreo resultó ser mucho mayor que el arbustivo y herbáceo (representó más del 98% en ambos bosques con respecto al total estimado en biomasa aérea).
- De acuerdo a los resultados obtenidos con las ecuaciones alométricas desarrolladas para Abies religiosa, éstas sobreestiman la biomasa aérea y, por lo tanto, su contenido de carbono, con excepción de la desarrollada por Brown et al. (1997). Dado lo anterior, es necesario explorar funciones que representen de manera más adecuada la acumulación de biomasa y carbono en la vegetación arbórea, ya que existen indicios de que las funciones exponenciales representan adecuadamente solo una parte del ciclo de crecimiento, por lo que se concluye que es necesario construir ecuaciones alométricas que representen el ciclo de crecimiento completo, incluyendo árboles de grandes dimensiones.
- Es importante medir los depósitos de carbono que la mayoría de los estudios no estiman: biomasa subterránea, madera muerta sobre el suelo y mantillo; ya que su aporte como depósito de C aunque no es tan grande como el de biomasa aérea y COS, en conjunto se estimó en este estudio entre 14% (bosque de Cupressus lusitanica) y 19% (bosque de Abies religiosa), pero dada la extensión de los bosques en nuestro país, representan un depósito de carbono importante, mismo que debe ser considerado como parte del ciclo global del C. Adicionalmente, la estimación del contenido de C en estos depósitos representa un avance en el conocimiento del ciclo de este elemento en el sector silvícola; además de que juegan un rol muy importante en la salud del ecosistema, ya que son fuentes primarias de nutrientes y almacén de combustibles para potenciales incendios forestales.
- Finalmente, se encontró que el contenido de C en el bosque de oyamel fue superior a lo reportado en otros sitios con vegetación similar, por lo que resulta importante conservar este tipo de vegetación como reservorio de carbono en el área de estudio y nos da una idea precisa del potencial como almacén de C que este tipo de vegetación tiene cuando llega a estados maduros.

