











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En México, los cereales de grano pequeño se siembran bajo riego y en temporal o secano; la cebada, el trigo y el triticale, se cultivan en un área aproximada de 322 000, 713 000 y 4800 ha, con un rendimiento de grano promedio nacional de 2.7, 5.2 y 2.4 Mg ha-1, respectivamente (SAGARPA, 2014). En condiciones de temporal en el ciclo primavera-verano el rendimiento de cebada (2 Mg ha-1), trigo (2.1 Mg ha-1) y triticale (2.4 Mg ha-1) es más bajo que en riego en el ciclo otoño-invierno (cebada = 5.5; trigo = 5.8 y triticale = 3.2 Mg ha‑1), debido a las deficiencias hídricas del suelo que con frecuencia coinciden con las etapas de floración y llenado de grano, al reducir severamente la producción y calidad del grano (López-Castañeda, 2013).
La reducción en el rendimiento y calidad del grano puede ser mayor en áreas en las que el trigo se siembra en el ciclo otoño-invierno (aproximadamente 20 000 ha) (SAGARPA, 2014), donde el cultivo depende de la humedad almacenada en el suelo después de la temporada de lluvias y las plantas experimentan altos niveles de estrés hídrico que se reflejan en rendimientos aún más bajos (0.6 Mg ha-1) (SAGARPA, 2014), como suele suceder en la región Mixteca Alta de Oaxaca y en otros estados como Nuevo León, Michoacán y Baja California. En estas condiciones ecológicas tan drásticas, el rendimiento se podría incrementar significativamente, al seleccionar variedades con caracteres agronómicos que les permitan adaptarse mejor a los ambientes con sequía, sobre todo en las etapas de floración y formación del grano.
Al considerar que los cultivos de temporal con frecuencia enfrentan serias limitaciones en la cantidad de agua disponible en el suelo, para su crecimiento, desarrollo y rendimiento, la selección debe estar enfocada a obtener genotipos resistentes o tolerantes a sequía, que posean atributos que le permitan a la planta: (i) capturar mayor cantidad de humedad disponible, (ii) intercambiar agua transpirada por moléculas de CO2 más efectivamente para producir biomasa y (iii) convertir mayor proporción de biomasa en grano (p. ej.: la proporción de materia seca que es grano o índice de cosecha) (Passioura y Angus, 2010). Aunque estos tres componentes del rendimiento bajo condiciones limitantes de humedad con frecuencia interactúan entre sí, son suficientemente independientes para evaluarlos en forma separada, de tal manera que la selección en alguno de ellos resultará en un aumento en el rendimiento (Passioura, 2006).
Un incremento en la captura de agua por las raíces, puede lograrse al seleccionar genotipos con sistemas radicales de mayor densidad y profundidad que absorban mayor cantidad de agua y nutrientes; el mejoramiento de la habilidad de las raíces para absorber agua y nitrógeno del suelo más pronto y más rápido es una estrategia eficiente para la sincronización entre la disponibilidad de nitrógeno y agua en el perfil del suelo y su demanda por la planta; esta estrategia dicta que las raíces crezcan rápido, inicien su proliferación temprana y profusamente para interceptar y capturar el nitrógeno, y el agua antes de que estos se muevan fuera del alcance de las raíces en el perfil del suelo (Palta et al., 2007). Las raíces juegan un papel muy importante para mantener el balance hídrico de la planta bajo condiciones de déficit hídrico edáfico y su importancia como característica de resistencia a sequía en cereales de grano pequeño, se ha reconocido a través de los años (Salim et al., 1965). Sin embargo, a la fecha son pocos los estudios realizados, para determinar los atributos de crecimiento de las raíces que puedan contribuir a un mejor comportamiento agronómico y mayor rendimiento de grano bajo condiciones de déficit hídrico en el suelo; Kramer (1969) y Richards (2008) sugirieron que las características de crecimiento de las raíces que una planta resistente a sequía debe tener: son un sistema radical profundo, extenso y muy ramificado como el del sorgo, no obstante, la conexión entre el rendimiento de grano y la cantidad de humedad utilizada por la planta apenas es perceptible, particularmente en un cultivo de grano, cuyo rendimiento depende de la cantidad y la etapa fenológica en que es utilizada.
El trigo ‘Venturero’ cultivado en la Mixteca Alta Oaxaqueña, es un buen ejemplo de las plantas adaptadas a condiciones de humedad residual que experimentan sequía, al disminuir el contenido de humedad en el suelo conforme transcurre el ciclo biológico; en estas condiciones, las plantas desarrollan sistemas de raíces que continúan en expansión a pesar del déficit hídrico edáfico, confiriendo a la planta, la capacidad de sobrevivir a la deshidratación y producir grano; el trigo ‘Venturero’ proviene del germoplasma que fue introducido a México por los españoles, en la época de la colonia y que ha sido cultivado en condiciones marginales de humedad y temperatura, y baja fertilidad del suelo en el ciclo otoño-invierno bajo condiciones de humedad residual en el suelo, desarrollando una alta capacidad de adaptación a este tipo de ambientes; las poblaciones de trigo ‘Venturero’ se caracterizan por presentar una amplia variabilidad genética en precocidad, altura de planta y espigas con y sin aristas. El rendimiento de grano de este germoplasma ‘criollo’, conocido también como ‘Venturero’, es bajo comparado con el que se obtiene en las siembras convencionales de trigo en temporal, sin embargo, el grano es apreciado por su calidad de gluten que permite su uso en la elaboración de tortillas para la alimentación humana.
Otra posibilidad de aumentar el rendimiento de grano en cereales de grano pequeño bajo condiciones limitantes de humedad, es a través de un intercambio gaseoso más efectivo entre el área foliar de la planta y la atmosfera (moléculas de H2O por moléculas de CO2), de tal manera que esto se traduzca en una mayor acumulación de biomasa en la planta; no obstante que el intercambio gaseoso de las plantas es afectado por el estrés hídrico, al reducirse la concentración de CO2 en el interior de las hojas (Passioura y Angus, 2010); un rápido déficit hídrico de la planta conlleva un rápido cierre de estomas, pero un déficit hídrico lento como ocurre típicamente en condiciones de campo, cuando el suelo se está deshidratando puede tener un efecto mayor en el desarrollo del área foliar con efectos más graduales en la conductancia estomática (Condon et al., 1992); en estas condiciones el cierre estomático usualmente resulta en un aumento en la temperatura de las hojas y consecuentemente en un mayor déficit de presión de vapor, de tal manera que el impacto de la baja concentración interna de CO2 en el eficiencia transpiratoria de la planta puede ser menor (Condon et al., 2002). El estrés hídrico también puede afectar otras características de la planta como la altura que disminuye a consecuencia de una reducción en la acumulación de materia seca, tasa de asimilación neta, tasa relativa de crecimiento, tasa de aparición de hojas y macollos, y la tasa de expansión del área foliar; y el índice de cosecha que decrece debido a la muerte de florecillas durante la emergencia de la hoja bandera, al iniciarse el estrés hídrico severo típico de ambientes con sequía terminal y que se refleja en una reducción en el número de granos por espiga y granos m-2 (Siddique et al., 1989). Además, la sequía también puede reducir significativamente la expansión del área foliar y la acumulación de materia seca de las plantas, y puede afectar la duración de las etapas críticas de desarrollo en el periodo de pre-antesis (iniciación floral, espiguilla terminal y espigamiento) y la duración de las etapas reproductivas en periodo post-antesis (floración o antesis, periodo de llenado del grano y madurez fisiológica) (López-Castañeda y Richards, 1994b); el déficit hídrico tiene efectos más severos en el rendimiento, cuando coincide con la antesis y el llenado del grano (Kirby y Appleyard, 1984). Wang et al. (2015) observaron que un déficit hídrico moderado en la etapa de crecimiento vegetativo en plantas de trigo, disminuyó el efecto de foto-inhibición en la hoja bandera causado por la sequía y el calor durante el periodo de llenado del grano; adicionalmente determinaron que en las plantas tratadas con déficit hídrico moderado, la sequía inhibió la fotosíntesis, principalmente a través de una disminución en la tasa fotosintética máxima del transporte de electrones, mientras que la disminución en la eficiencia de la carboxilación limitó la fotosíntesis bajo condiciones de estrés por calor; la sequía durante la etapa vegetativa mejoró la tolerancia a ambos, la sequía y el estrés por calor, ocurridos durante el periodo de llenado del grano.
La conversión de la biomasa en grano, representada por el índice de cosecha (p. ej.: el cociente entre el rendimiento de grano y la biomasa aérea de la planta) ha aumentado substancialmente en muchos cultivos durante las últimas décadas; la introducción de los genes de enanismo en los cereales de grano pequeño para resolver el problema de acame tuvo beneficios inesperados al aumentar el índice de cosecha; en ambientes favorables de humedad pasó de 0.33 a más de 0.50 (Passioura y Angus, 2010). Este incremento ha sido asociado con un aumento en el número de granos m-2, con un aumento pequeño en el peso del grano o producción de biomasa (Evans, 1993). La selección de genotipos mejor adaptados al patrón de distribución de la humedad durante la estación de crecimiento, puede resultar en un rápido desarrollo del área foliar que reduzca significativamente las pérdidas de humedad por evaporación directa del suelo debajo del dosel del cultivo y favorezca un buen balance de la evapotranspiración antes y después de la floración, lo que se reflejará en un mayor índice de cosecha (López-Castañeda y Richards, 1994c; Passioura, 2006).
En un futuro se requerirán sistemas integradores para el estudio de caracteres cuantitativos complejos como la estabilidad del rendimiento de plantas bajo estrés hídrico y la investigación deberá utilizar los recursos genómicos más modernos, combinando las nuevas tecnologías en genética cuantitativa, genómica y biomatemáticas, con un amplio conocimiento ecofisiológico de las interacciones entre el genotipo y el ambiente de crecimiento, y finalmente, el mejoramiento genético del cultivo (Parry et al., 2012).
El presente trabajo de investigación tuvo como objetivo estudiar la variabilidad genética en el rendimiento de grano y sus componentes, y la densidad, y profundidad de raíces, características morfológicas que pueden contribuir a un mejor comportamiento agronómico de la cebada, el trigo (antiguas y modernas variedades y una criolla denominado ‘Venturero’) y el triticale, en condiciones limitantes de humedad residual del suelo en el ciclo otoño-invierno, en la Mixteca Alta Oaxaqueña.
Materiales y Métodos
Sitio experimental
Se estableció un experimento en campo bajo condiciones de humedad edáfica residual en el ciclo otoño-invierno 2013-2014, en la comunidad de San Cristóbal Suchixtlahuaca, Distrito de Coixtlahuaca, Oaxaca, México (17° 44’ N, 97° 19’ O y 2130 m de altura). Esta localidad se caracteriza por presentar clima cálido semiseco (BS1kw(w)(i’)gw’’), con verano cálido y seco; temperatura media anual de 15.8 °C y precipitación anual de 556 mm, con lluvias en verano y escasa precipitación en invierno, y poca oscilación climática (García, 1988). El suelo donde se estableció el experimento es de textura arcillosa en el perfil de 0 a 80 cm; en el estrato de 0-20 cm mostró valores de densidad aparente ligeramente más altos que en los otros estratos y valores de humedad ligeramente más bajos en presiones equivalentes a la capacidad de campo (CC) y porcentaje de marchitamiento permanente (PMP). El pH en el estrato de 0-20 cm fue más bajo que en los otros estratos, mientras que el contenido de materia orgánica, nitrógeno total, fósforo y potasio fue mayor en el estrato de 0-20 cm que en los estratos de 20-40, 40-60 y 60-80 cm; la conductividad eléctrica del suelo tendió a presentar valores más altos en los estratos de 40-60 y 60-80 cm que en los estratos de 0-20 y 20-40 cm (Cuadro 1).
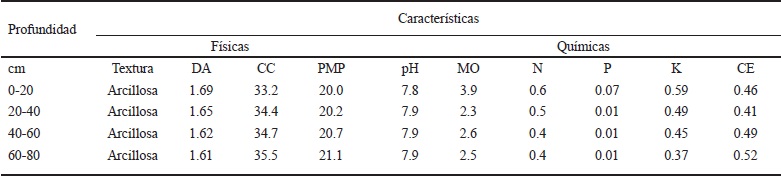
Textura, método del hidrómetro de Bouyoucos; DA = densidad aparente, método de la probeta (g cm-3); CC = capacidad de campo (%), olla de presión a 0.1 kPa; PMP = porcentaje de marchitamiento permanente (%), membrana de presión a 15 kPa; pH, medidor electrónico de pH; MO = materia orgánica (%), método de Walkley y Black; N = nitrógeno total (%), método de Kjeldahl, semi-micro; P = fósforo (ppm), método de Olsen; K = potasio (ppm), método de Olsen; C.E. = conductividad eléctrica (mmhos cm-1), método del puente de conductividad eléctrica.
Cuadro 1 Características físicas y químicas del suelo utilizado para el experimento.
Material genético
Se utilizaron 20 genotipos de trigos harineros antiguos y modernos, y un criollo ’Venturero’, tres genotipos de cebada y uno de triticale (Cuadro 2). Los genotipos de trigo incluyeron materiales criollos antiguos como ‘Gabo’ y ‘Marroqui’, genotipos considerados como progenitores de la ‘Revolución Verde’; ‘Yaqui 50’ un material criollo originario del Valle del Yaqui, Sonora, México y otros genotipos representativos de diferentes décadas de la ‘Revolución Verde’; además de dos genotipos de cebada de dos hileras (Betzes y Lenetah) y una de seis hileras (Cleopatra), y un triticale experimental; Betzes, Lenetah y Triticale 118 se introdujeron de la Universidad de California, Riverside, CA, Estados Unidos de Norte América (Cuadro 2).

† La multiplicación del germoplasma y los datos del peso de 100 granos se obtuvieron en el ciclo de verano 2010 en condiciones de secano, en el Colegio de Postgraduados, Montecillo, Texcoco, México; ‡ Germoplasma obtenido de la Colección de Cereales de Grano Pequeño (Small Cereals Collection, Idaho, U.S.A); ¶ Material genético obtenido del INIFAP; Chapingo, México; †† Variedades obtenidas del CIMMYT, El Batán, Texcoco, Estado de México; § Germoplasma proveniente de la Universidad de California, Riverside, CA, U.S.A.; # Material genético colectado en la Comunidad de San Cristóbal Suchixtlahuaca, Oaxaca, en Septiembre de 2013; ‡‡ Línea avanzada procedente del ICARDA, Alepo, Siria; §§ Genotipos de trigo obtenidos por el método genealógico, Dr. Cándido López Castañeda, Profesor Investigador Titular, Colegio de Postgraduados.
Cuadro 2 Genotipos de trigo, triticale y cebada utilizados en el experimento.
Manejo del trabajo experimental
Se utilizó un diseño experimental de bloques completos al azar con cuatro repeticiones; la unidad experimental consistió de cuatro surcos de 4 m de longitud, separados a 0.40 m. La siembra se realizó el día 5 de octubre de 2013 en suelo seco, se depositó la semilla en el fondo del surco y cubrió con azadón. Se utilizó una densidad de siembra de 100 kg de semilla ha‑1 y se aplicó una dosis de fertilización de 40‑40-00 al momento de la siembra, con urea como fuente de nitrógeno y superfosfato de calcio triple como fuente de fósforo. Al observar que no llovía cinco días después de la siembra, se procedió a dar un riego para favorecer la germinación de la semilla en el suelo; el agua se consiguió de una pequeña presa localizada a 300 m del lugar en el que se estableció el experimento, el agua se aplicó con una manguera de 3” de diámetro, en una cantidad suficiente para humedecer los 5 cm más cercanos a la superficie del suelo. De la misma forma se aplicó un segundo riego a los 20 dds; tres días después, se presentaron las primeras lluvias y el experimento se dejó a merced de la precipitación hasta la madurez fisiológica. Las malezas de hoja ancha y de hoja angosta se eliminaron mediante deshierbes manuales entre los 30 y 60 dds. No se observó la incidencia de plagas.
Variables de estudio
Días a antesis (DA); se determinó cuando el 50% de las espigas mostraban las anteras expuestas en dehiscencia.
Días a madurez fisiológica (MF); se determinó cuando el 80% de los órganos aéreos de las plantas presentaban el color paja característico de cada genotipo.
Biomasa aérea final (BM, g m-2); se obtuvo al cosechar los tallos completos presentes en los surcos centrales de cada unidad experimental, después de eliminar los extremos, para determinar su peso seco total y dividirlo entre el área cosechada.
Rendimiento de grano (RG, g m-2); se obtuvo al determinar el peso del grano después del desgrane de la muestra utilizada para determinar la biomasa aérea final.
Índice de cosecha (IC); se calculó al dividir el rendimiento de grano entre la biomasa área final (RG/BM).
Espigas M-2 (E M-2); se determinó al contar el número total de tallos fértiles (NTF) con espiga y dividirlo entre el área cosechada.
Peso de 100 granos (P100g, g); se determinó al pesar 100 granos elegidos al azar en cada unidad experimental.
Granos por espiga (G E-1); se determinó al contar y promediar el número de granos por espiga en 10 espigas completas de la muestra utilizada, para determinar la biomasa aérea final.
Granos M-2 (G M-2); se calculó al multiplicar el número de espigas m-2 por el número de granos por espiga (E M-2 × G E-1).
Altura de planta (AP, cm); se midió la longitud del tallo desde la superficie del suelo hasta la última espiguilla en la espiga en cinco plantas en cada unidad experimental.
Peso hectolítrico o densidad del grano (PH, g mL‑1); se calculó como el cociente entre el peso (g) y el volumen (mL) de una muestra de 10 g de grano, tomada al azar del grano cosechado en cada unidad experimental.
Densidad de raíces (DR, cm cm-3); se obtuvieron muestras de suelo del experimento para calcular la densidad de raíces. Las muestras se tomaron de todas las parcelas de Triticale 118, Cleopatra y Lenetah (cebada) y Criollo ‘Venturero’, Yaqui 50, Marroquí, Tlaxcala F-2000 y Náhuatl F-2000 (trigo). Se extrajeron cuatro muestras de suelo de cada parcela con una barrena del tipo Veihmeyer, con boquilla de 2.1 cm de diámetro en los estratos de 0-20, 20‑40, 40-60 y 60-80 cm; dos sobre la línea de plantas y dos entre líneas de plantas. Las raíces se separaron del suelo completamente seco con pinzas de depilar y se colocaron en viales de plástico con 50 ml de agua destilada; posteriormente, los segmentos de raíz se dispersaron en 500 ml de agua en una charola de plástico transparente de 30 × 40 cm, colocada sobre una cuadrícula de 0.5 × 0.5 cm (Tennant, 1975). Se contó el número de intersectos entre los segmentos de raíz y las líneas verticales y horizontales de la cuadrícula (Tennant, 1975). La densidad de raíces (cm de raíz cm-3 de suelo) se estimó con el producto del número de intersectos × el factor de conversión de longitud (0.3928), para una cuadrícula de 0.5 × 0.5 cm (Tennant, 1975; López-Castañeda y Richards, 1994a).
Datos meteorológicos
Los datos diarios de temperatura máxima y mínima del aire, y la precipitación acumulada semanalmente se registraron con un termómetro de máxima y mínima de columna de mercurio (Marca Taylor) y un pluviómetro de acumulación semanal en el lugar en el que se estableció el experimento en condiciones de campo, respectivamente.
Contenido de humedad edáfica
Se extrajeron muestras de suelo con una barrena del tipo Veihmeyer, para determinar el contenido hídrico en los estratos de 0-20, 20-40, 40-60 y 60-80 cm, semanalmente durante el experimento. Las muestras de suelo se secaron en una estufa con circulación de aire forzado marca GCA Corporation, U.S.A., modelo 17. El contenido de humedad aprovechable (HA) se calculó de acuerdo al método gravimétrico [HA (%) = ((Peso del suelo húmedo - Peso del suelo seco) / Peso del suelo seco) 100].
Análisis estadístico
Se realizó un análisis de varianza para las variables estudiadas, utilizando el programa SAS (2009) (Statistical Analysis System, Versión 9.1, para Windows). Para la comparación de medias, se calculó la diferencia mínima significativa (DMS, P ≤ 0.05).
Resultados y Discusión
Temperatura, lluvia y contenido hídrico del suelo
Las temperaturas mínimas y máximas promedio registradas durante el ciclo fueron 7 y 24 °C (Figura 1); estos valores de temperatura se consideran favorables para el crecimiento y desarrollo del trigo y la cebada, considerando que la temperatura base para el crecimiento de la cebada y el trigo tiene un valor de 0º C (Cao y Moss, 1989; López-Castañeda et al., 1996) y un valor de 30-32 °C, para la temperatura máxima a la que el crecimiento de las plantas empieza a disminuir rápidamente (Vincent, 1989).
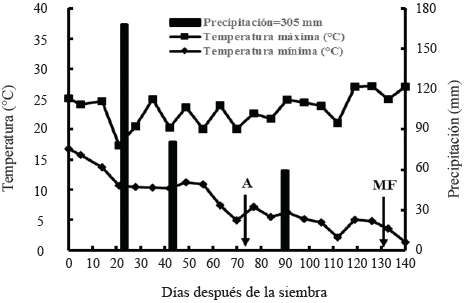
Figura 1 Temperatura máxima y mínima promedio semanal, y precipitación semanal acumulada durante el ciclo otoño-invierno 2013-2014. San Cristóbal Suchixtlahuaca, Oaxaca. A = antesis o floración y MF = madurez fisiológica.
La cantidad de lluvia registrada durante el experimento fue 305 mm de los cuales, el 81% ocurrió en las primeras tres semanas después de la siembra y el 19% en la etapa de elongación del tallo (56 dds) y al inicio del periodo de llenado de grano (84 dds) (Figura 1). La distribución de la lluvia durante el experimento, mostró un patrón de sequía intermitente durante el periodo de preantesis y un patrón de sequía terminal durante el periodo de postantesis (Figura 1); las deficiencias hídricas en ésta etapa reducen considerablemente el contenido hídrico en las células de la planta, mostrando síntomas de marchitamiento y perdida de turgencia, que repercuten en una disminución del intercambio gaseoso y fotosíntesis de la planta y finalmente en el rendimiento de grano (Blum, 2011).
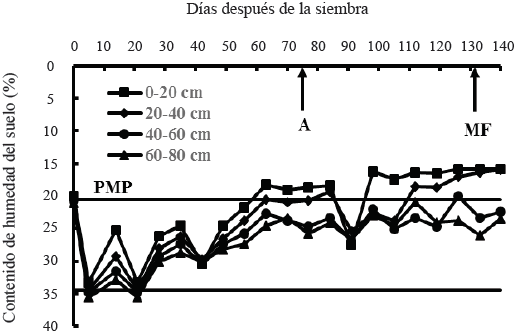
El contenido hídrico en todos los estratos del suelo se mantuvo cercano a capacidad de campo (CC) entre los cinco (aplicación del riego de siembra) y 21 dds (168 mm de lluvia); se observó un segundo y tercer pico de alta humedad aprovechable en todos los estratos del suelo, debido a las precipitaciones ocurridas a los 42 (80.5 mm de lluvia) y 91 dds (59 mm de precipitación). No obstante, estos periodos de alta humedad aprovechable en el suelo, se observaron periodos de sequía intermitente en el suelo; el contenido hídrico edáfico disminuyó por debajo del porcentaje de marchitamiento permanente (PMP) en los estratos de 0-20 y 20-40 cm entre los 63 y 84 dds, y entre los 98 y 140 dds en el estrato de 0-20 cm, y entre los 112 y 140 dds en el estrato de 20-40 cm (Figura 2). El primer periodo de sequía en el suelo coincidió con las etapas de espigamiento, antesis e inicio de la formación del grano, y el segundo periodo de déficit hídrico edáfico se presentó durante la mayor parte del periodo de llenado del grano hasta la madurez fisiológica (Figura 2). La mayor disminución en el contenido hídrico del suelo en los estratos de 0-20 y 20-40 cm, se debió a una mayor extracción de agua, al considerar que el 50% de las raíces, se concentran en estos estratos del suelo en trigo, cebada y triticale en condiciones de secano (López-Castañeda y Richards, 1994a).
Comparación del rendimiento de grano y sus componentes entre especies
Las mayores diferencias en rendimiento de grano y sus componentes, se observaron entre las especies estudiadas; la cebada produjo 47, 12, 37, 16 y 32% mayor rendimiento de grano, biomasa aérea, índice de cosecha, número de granos espiga-1 y peso de 100 granos que el trigo, y 47, 47 y 32% mayor rendimiento de grano, índice de cosecha y peso de 100 granos que el triticale (Cuadro 3). La cebaba tuvo mayor rendimiento de grano en menor número de días a antesis y madurez f isiológica que el trigo y el triticale, y tuvo menor peso hectolítrico del grano y altura de planta que el trigo y el triticale. El triticale produjo 27, 15 y 18% mayor número de espigas m-2, número de granos m-2 y peso hectolítrico que la cebada, y 18, 31, 25 y 7% mayor biomasa aérea, número de espigas m-2, número de granos m-2 y peso hectolítrico que el trigo. El trigo tuvo mayor altura de planta y mayor número de días a antesis que la cebada y el triticale (Cuadro 3).
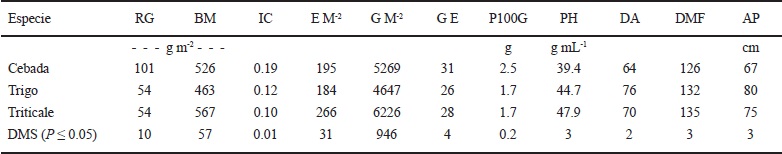
RG = rendimiento de grano; BM = biomasa aérea final; IC = índice de cosecha; E M-2 = espigas m-2; G M-2 = granos m-2; G E = granos por espiga; P100G = peso de 100 granos; PH = peso hectolítrico; DA = días a antesis; DMF = días a madurez fisiológica; AP = altura de planta.
Cuadro 3 Rendimiento y características fenológicas y fisiológicas de los cultivos de cebada, trigo y triticale en condiciones de secano, en el ciclo otoño-invierno 2013-2014. San Cristóbal Suchixtlahuaca, Oaxaca.
La cebada fue menos afectada por la sequía que el trigo y el triticale, debido a que completó sus procesos de desarrollo fenológico en menor tiempo y de ésta forma, evadió en mayor grado el severo estrés hídrico terminal durante el periodo de llenado del grano; otros estudios en condiciones de secano con sequía terminal han determinado que la cebada supera en rendimiento de grano al trigo y al triticale, debido a su más rápido desarrollo del área foliar, mayor acumulación de materia seca y mayor precocidad (López Castañeda y Richards, 1994b).
Un estudio conducido bajo condiciones de secano con líneas de porte alto, intermedio y bajo de avena, cebada de primavera e invierno, trigo de invierno y centeno de invierno, mostró que las variedades de cebada de invierno y primavera tuvieron mayor rendimiento de grano que las variedades de trigo, avena y centeno invernales, debido a su mayor índice de cosecha (Singh y Stoskopf, 1971); se observó también que la altura de planta estuvo negativamente relacionada con el índice de cosecha y que una reducción en la altura de planta, la cual reduce el tamaño de la demanda del tallo, pudo haber disminuido la capacidad de almacenamiento de asimilados en el tallo; una correlación negativa y significativa entre la altura de planta y el índice de cosecha en trigos de invierno, sugiere que el índice de cosecha puede mejorar, reduciendo el tamaño de la planta (Singh y Stoskopf, 1971; Fischer, 1989).
El índice de cosecha fue el componente que más contribuyó al alto rendimiento de grano de cebada posiblemente, debido a una mejor distribución de los asimilados entre los órganos vegetativos y el grano (Singh y Stoskopf, 1971). Estos autores señalan también que un mayor índice de cosecha y rendimiento de grano bajo sequía, pueden lograrse, al seleccionar plantas de menor porte, entrenudos más cortos o menor número de entrenudos y un periodo vegetativo más corto; la cebada reúne algunos de estos caracteres, lo que puede explicar su mejor comportamiento que el trigo y el triticale, en condiciones de sequía terminal (López-Castañeda y Richards, 1994b). Otros caracteres de la planta que pueden contribuir a mayor índice de cosecha y rendimiento de grano, bajo condiciones de estrés hídrico terminal en cereales de grano pequeño: son una mayor tasa de llenado de grano y mayor capacidad de removilización de reservas del tallo al grano; en condiciones de sequía terminal en el sureste de Australia, se determinó que la removilización de reservas del tallo al grano puede contribuir al rendimiento de grano de cebada hasta en 70% y en trigo 57% (López-Castañeda y Richards, 1994b).
Comparación del sistema radical entre especies
La mayor densidad radical se concentró cerca de la superficie del suelo (0-20 cm) en la cebada (69%), el trigo (56%) y el triticale (71%) (Figura 3); la densidad de raíces en promedio de los diferentes genotipos dentro de especies, mostró que el trigo tuvo 39, 63 y 33% mayor densidad radical que la cebada en los estratos de 40-60, 60-80 y 80-100 cm, y 50, 69, 63 y 100% mayor densidad radical que el triticale, en los estratos de 20‑40, 40-60, 60-80 y 80‑100 cm (Cuadro 4). En el estrato de 0-20 cm no se observaron diferencias significativas entre especies y en el estrato de 20-40 cm, no se observaron diferencias significativas entre la cebada y el trigo (Cuadro 4). Los valores de la densidad de raíces observados en el presente estudio son más bajos en promedio de las tres especies que los determinados para las mismas especies en el sureste de Australia (López-Castañeda y Richards, 1994a) y Australia Occidental (Siddique et al., 1989) bajo en condiciones de sequía terminal; estas diferencias pueden estar relacionadas con el número de días requeridos para alcanzar la antesis que el presente estudio fue 70 días y en el sureste de Australia varió de 118 a 134 dds (López-Castañeda y Richards, 1994a) y de 100-125 dds en Australia Occidental (Siddique et al., 1989); se ha determinado que el crecimiento del sistema radical en trigo culmina cuando las plantas alcanzan la etapa de antesis o floración (Barraclough, 1984).
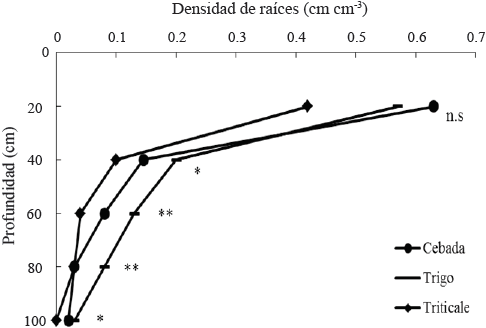
Figura 3 Densidad radical en diferentes estratos del suelo, de los cultivos de cebada, trigo y triticale en condiciones de secano, en el ciclo otoño-invierno 2013‑2014. San Cristóbal Suchixtlahuaca, Oaxaca. **(P ≤ 0.01); *(P ≤ 0.05); n.s. = no significativo.

Cuadro 4 Densidad radical en los estratos de 0-20, 20-40, 40-60, 60-80 y 80-100 cm, de los cultivos de cebada, trigo y triticale en condiciones de secano, en el ciclo otoño-invierno 2013-2014. San Cristóbal Suchixtlahuaca, Oaxaca.
La superioridad en densidad del sistema radical de la cebada sobre el trigo y el triticale, en condiciones de sequía terminal se ha observado también en otros trabajos de investigación; López-Castañeda y Richards (1994a) determinaron que la cebada produjo 40% mayor densidad de raíces que trigo y triticale en los 15 cm más cercanos a la superficie del suelo, manteniendo significativamente mayor densidad radical hasta los 90 cm de profundidad; Atta et al. (2013) confirmaron que la mayor densidad de raíces en cebada y trigo bajo estrés hídrico, se concentró cerca de la superficie del suelo (15 cm de profundidad). También, Gregory et al. (1992) y Atta et al. (2013) observaron que la cebada produjo mayor densidad radical que trigo entre los 0 y 75 cm de profundidad del suelo en condiciones de secano con estrés hídrico terminal en Australia Occidental.
Las divergencias en la densidad radical entre el trigo y la cebada, y el triticale, observadas en los estratos de 40-100 cm, pueden deberse a diferencias en el número de días a floración y consecuentemente, la duración del crecimiento y cambios en la intercepción de luz y arquitectura del dosel; estos factores son importantes para el crecimiento del sistema radical, en virtud de que la duración del funcionamiento del área foliar y la oferta de carbono para el sistema radical tienen influencia en el crecimiento y exploración de las raíces (Watt et al., 2013).
Comparación del rendimiento de grano y sus componentes entre genotipos
La variabilidad genética en rendimiento de grano y sus componentes entre genotipos individuales fue substancial; la variedad de cebada Cleopatra produjo mayor rendimiento de grano, índice de cosecha y granos espiga-1 que todos los demás genotipos; además, Cleopatra registró el menor número de días a madurez fisiológica (Cuadro 5). También, se observaron diferencias entre genotipos para la biomasa aérea final, espigas m-2, número de granos m-2 y altura de planta; Cleopatra, Lenetah , Betzes, Triticale 118, criollo ‘Venturero’, Yaqui 50, Marroqui, Nainari 60, Huamantla, Salamanca S-75, Verano S-91, Romoga F-96, Borlaug M-95 y la línea F6 de ICARDA, tuvieron mayor biomasa aérea final que los demás genotipos; Betzes y el Triticale 118 produjeron mayor número de espigas m-2 que los otros genotipos; Cleopatra, Lenetah, Betzes, Triticale 118, criollo ‘Venturero’, Yaqui 50, Nainari 60, Huamantla, Salamanca S-75, Verano S-91, Romoga F-96, Borlaug M-95, Blanca Grande, Rebeca F-2000, Náhuatl F-2000 y la línea F6 de ICARDA, tuvieron mayor número de granos m-2 que los demás genotipos; Cleopatra y el criollo ‘Venturero’ obtuvieron mayor peso de 100 granos que los demás genotipos; y Marroquí tuvo la mayor altura de planta (Cuadro 5).
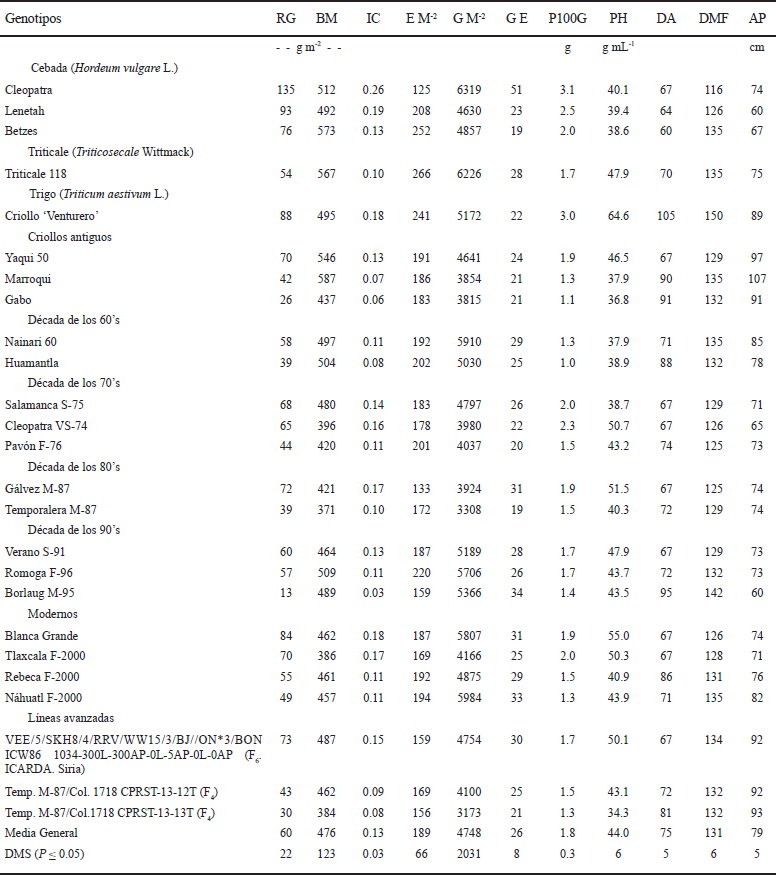
RG = rendimiento de grano; BM = biomasa aérea final; IC = índice de cosecha; E M-2 = espigas m-2; G M-2 = granos m-2; G E = granos por espiga; P100G = peso de 100 granos; PH = peso hectolítrico; DA = días a antesis; DMF = días a madurez fisiológica; AP = altura de planta.
Cuadro 5 Rendimiento y características fenológicas y fisiológicas de los diferentes genotipos de cebada, trigo y triticale en condiciones de secano, en el ciclo otoño-invierno 2013-2014. San Cristóbal Suchixtlahuaca, Oaxaca.
En otros estudios en cereales de grano pequeño bajo condiciones de estrés hídrico terminal, también se ha confirmado que la cebada tiene un mejor comportamiento agronómico que el trigo, el triticale y la avena; la mejor variedad de cebada O’Connor produjo 39% más grano que la mejor variedad de trigo Kulin, debido a que su biomasa aérea final e índice de cosecha fueron 25 y 4% mayores que los de Kulin, en dos localidades de secano en Nueva Gales del Sur, Australia (López-Castañeda y Richards, 1994a). En otro trabajo de investigación con cebada bajo condiciones de riego, sequía y temporal se determinó que el rendimiento de grano estuvo positivamente relacionado con la biomasa aérea final y el número de granos m-2; las líneas que produjeron mayor rendimiento de grano también tuvieron mayor biomasa aérea final y granos m-2 (López-Castañeda, 2011).
Comparación del sistema radical entre genotipos
Al comparar en forma individual las líneas utilizadas para determinar la densidad de raíces, se observó que la mayor proporción de raíces se concentró cerca de la superficie del suelo (0-20 cm), sin presentar diferencias significativas entre genotipos (Cuadro 6). El criollo ‘Venturero’ fue el genotipo que produjo más raíces en las profundidades de 20-100 cm (Figura 4). En el estrato de 60-80 cm también se observaron diferencias significativas en la densidad de raíces entre genotipos; Lenetah, Triticale 118, Yaqui 50, Marroqui, y Náhuatl F-2000 tuvieron mayor densidad de raíces que Cleopatra y Tlaxcala F-2000 (Figura 4).
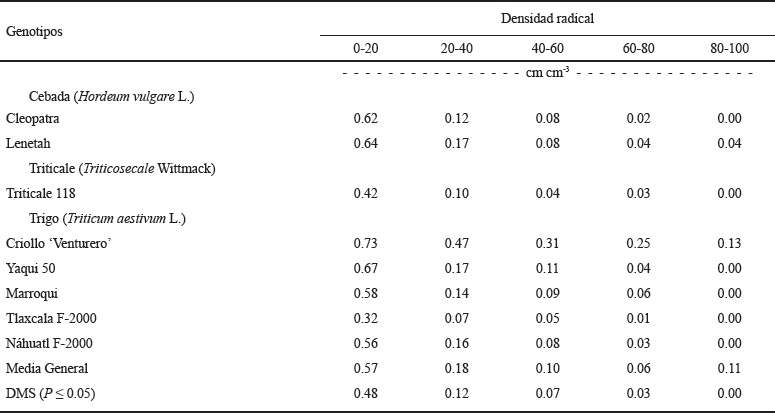
Cuadro 6 Densidad radical en los estratos de 0-20, 20-40, 40-60, 60-80 y 80-100 cm, de los cultivos de cebada, trigo y triticale en condiciones de secano, en el ciclo otoño-invierno, 2013-2014. San Cristóbal Suchixtlahuaca, Oaxaca.
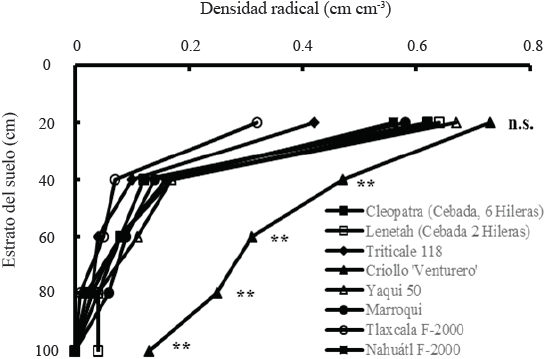
Figura 4 Densidad radical determinada en diferentes estratos del suelo en distintos genotipos de cebada, trigo y triticale en condiciones de secano, en el ciclo otoño-invierno 2013-2014. San Cristóbal Suchixtlahuaca, Oaxaca. **(P ≤ 0.01); n.s. = no significativo.
La presencia de una alta proporción de raíces cerca de la superficie del suelo se ha observado también en otros trabajos de investigación; Gregory et al. (1992) determinaron que tanto la variedad de trigo Gutha como las variedades de cebada O’Connor y Beecher, produjeron la mayor proporción de raíces cerca de la superficie del suelo (15 cm) y que el sistema radical de Gutha estuvo confinado a una profundidad menor de 60 cm, mientras que las raíces de O’Connor y Beecher penetraron hasta 80 cm de profundidad, en condiciones de secano con sequía terminal en Australia Occidental. En otro estudio de raíces bajo condiciones de estrés hídrico terminal en el sureste de Australia, se determinó que la cebada O’Connor tuvo 23% mayor densidad de raíces que la variedad Meteor de trigo cerca de la superficie del suelo (10 cm) y que las diferencias en densidad radical entre estos genotipos, se mantuvieron hasta los 90 cm de profundidad del suelo (López-Castañeda y Richards, 1994a).
El mayor tamaño de las raíces en el trigo ‘Venturero’, le confiere una mayor capacidad de adaptación a condiciones de déficit hídrico; estudios previos en la comparación del tamaño del sistema radical entre trigos antiguos y modernos, indican que los cultivares antiguos tienen sistemas de raíces más grandes que los cultivares modernos; un sistema radical más grande, extenso y profundo, puede optimizar la absorción de agua y nutrientes, y ayudar a maximizar el rendimiento de grano (Waines y Ehdaie, 2007). En el presente estudio, el trigo ‘Venturero´ tuvo la más alta densidad de raíces y produjo el rendimiento de grano más alto entre los genotipos de trigo, características indicativas de un mejor comportamiento de éste material en comparación con los otros genotipos de trigo, bajo condiciones de humedad residual con sequía terminal. La relación entre las características del sistema radical y el rendimiento de grano bajo condiciones de estrés hídrico también se ha demostrado en variedades de cebada maltera en Želešice, República Checa, donde se observó que las variedades con mayor tamaño del sistema radical produjeron mayor rendimiento de grano bajo sequía y tuvieron mayor contenido de almidón, sacáridos, extracto de malta y rendimiento de proteína (Chloupek et al., 2010).
Las variedades de cebada produjeron mayor rendimiento de grano que los genotipos de trigo y triticale gracias a su mayor índice de cosecha, peso o tamaño del grano y mayor densidad de raíces, así como la menor estatura de planta y ciclo biológico más corto; estos atributos hacen a la cebada uno de los cereales más populares en áreas de secano con déficit hídrico terminal durante la antesis y el periodo de llenado de grano, pues generalmente produce grano a pesar del estrés hídrico, debido principalmente a su rápido crecimiento y precocidad (López-Castañeda y Richards, 1994b), y sistema de raíces más profundo y profuso (López-Castañeda y Richards, 1994a). El trigo y el triticale presentaron menor rendimiento de grano que la cebada y tuvieron un comportamiento similar entre sí, sin embargo, el trigo ‘Venturero’ produjo un rendimiento de grano similar a las variedades de cebada Lenetah y Betzes, y a las variedades de trigo Gálvez M-87 y Blanca Grande; a diferencia de estos genotipos, el trigo ‘Venturero’ tuvo mayor número de espigas m-2 y tamaño del grano, y el más alto peso hectolítrico o densidad del grano, la mayor densidad de raíces, y el ciclo biológico más largo, corroborando su gran adaptación a condiciones de baja humedad residual y estrés hídrico terminal severo.
Los atributos de la planta que se deben considerar en los criterios de selección de nuevos genotipos de cereales de grano pequeño, adaptados a condiciones de humedad residual y sequía terminal, como las que se tuvieron durante el experimento, son un sistema radical profundo y profuso siendo el carácter genético más importante para lograr mejorar la resistencia a sequía y el rendimiento.
Conclusiones
-. La cebada fue la especie sobresaliente al tener mayor rendimiento de grano, biomasa aérea, índice de cosecha, número de granos por espiga y peso de 100 granos que el trigo, y mayor rendimiento de grano, índice de cosecha y peso de 100 granos que el triticale; la cebada produjo su alto rendimiento en menor número de días a antesis y madurez fisiológica que el trigo y el triticale.
-. El triticale tuvo un comportamiento interesante al producir mayor número de espigas m-2, número de granos m-2 y peso hectolítrico que la cebada, y mayor biomasa aérea, número de espigas m-2, número de granos m-2 y peso hectolítrico que el trigo; el trigo tuvo mayor altura de planta y número de días a antesis que la cebada y el triticale.
-. La mayor proporción de raíces en la cebada, el trigo y el triticale se concentró cerca de la superficie del suelo (20 cm), sin presentar diferencias significativas entre especies; el trigo produjo mayor densidad de raíces que la cebada entre los 40 y 100 cm, y mayor densidad radical que el triticale entre los 20 y 100 cm.
-. La variabilidad entre todos los genotipos en conjunto mostró que la variedad de cebada Cleopatra, fue la que tuvo el más alto rendimiento de grano, índice de cosecha y granos espiga-1; este mayor rendimiento de grano se obtuvo con menor número de días a madurez fisiológica que los demás genotipos.
-. El trigo ‘Venturero’ produjo mayor rendimiento de grano, índice de cosecha, número de espigas m-2, peso de 100 granos, peso hectolítrico, número de días a antesis y madurez fisiológica, densidad radical y tuvo una mejor distribución de raíces que los demás genotipos de trigo entre los 20 y 100 cm de profundidad.
-. Los genotipos de cebada y trigo con mayor densidad de raíces, produjeron mayor rendimiento de grano, corroborando su mayor resistencia a sequía o adaptación a condiciones de déficit hídrico terminal.

