











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La salinidad es uno de los procesos de degradación de los suelos que más daños causan a la agricultura, transformando tierras productivas y fértiles en estériles y frecuentemente conducen a perdidas en hábitat y reducción de biodiversidad (Clavero y Razz, 2002).
En las últimas décadas, las investigaciones sobre los efectos de la salinidad de los suelos en los cultivos, reflejan la importancia de este problema para la agricultura mundial. El estrés salino es uno de los factores ambientales adversos que influyen sobre aspectos de la fisiología de las plantas, lo que a su vez limita la productividad de los cultivos de interés económico. La salinidad reduce la capacidad de las plantas para absorber agua, ocasionando una reducción en el crecimiento (Munns, 2002). Altas concentraciones de sales en la solución externa de las células vegetales ocasiona efectos, que pueden resumirse fundamentalmente en tres tipos: sequía osmótica, toxicidad debida a la excesiva absorción de cloro y sodio y un desbalance nutrimental (Trinchant et al., 2004; Karimi et al., 2005).
A nivel mundial un bajo número de hortalizas presentan una alta demanda de consumo. Un cultivo representativo es el tomate, su importancia radica en sus cualidades para integrarse en la preparación de alimentos. En los últimos años, la producción mundial de ésta hortaliza, se ha mantenido estable con un nivel promedio anual de 100 millones de toneladas (Mujica et al., 2014). Según datos de la SAGARPA (2010), los principales productores de tomate son China, Estados Unidos, Turquía, India, Italia, Irán y Egipto, países que conjuntamente han producido durante los últimos 10 años el 70% de la producción mundial.
En México, el tomate es una de las especies hortícolas de mayor importancia económica y social, en virtud del valor de su producción y por la demanda de mano de obra que genera. Es el principal producto hortícola de exportación, representa el 35% del valor total de las exportaciones de legumbres y hortalizas y el 18% del valor total de las exportaciones agropecuarias, solo superadas por el ganado vacuno (SAGARPA, 2010).
El Estado de Baja California Sur es el cuarto Estado de la república Mexicana con mayor producción, ocupa el 7.5% de la producción anual, el tomate es una especie que se cultiva en aproximadamente 14 186 ha, con una producción promedio anual de 39.5 Mg ha-1, con un valor promedio anual cercano a los 150 millones de pesos (Lucero et al., 2012).
En este mismo sentido Navejas (2002), menciona que la producción de tomate orgánico, en México se lleva a cabo en Baja California Sur, pero uno de los problemas esenciales en la producción es la salinidad del suelo y del agua, además de la lucha contra los insectos y enfermedades.
En este mismo contexto FIRA (2011), establece que la producción de tomate en Baja California Sur mediante la agricultura protegida es de suma importancia, en México representa al tercer lugar con mayor superficie de producción de tomate en invernaderos y casas sombra con el 11.28%. La principal problemática relacionada con los aspectos de producción y prácticas agronómicas que enfrentan los productores en las principales regiones productoras del estado de Baja California Sur, es la escasa o casi nula información sobre los cultivares, híbridos o variedades que deben sembrarse para obtener rendimientos altos y de calidad de esta especie, incluyendo al mismo tiempo la interacción de los factores bióticos y abióticos.
Uno de los factores abióticos considerado como el principal problema es la calidad del agua para riego, la cual en la mayoría de las regiones, presenta altos contenidos de sales (principalmente cloruro de sodio) que es altamente perjudicial para el buen desarrollo y producción de las principales especies cultivadas (Goykovic y Saavedra, 2007). El productor no cuenta con un programa de mejoramiento genético para las condiciones agroecológicas prevalecientes en la entidad y falta realizar estudios con tomate en los aspectos de tolerancia a salinidad.
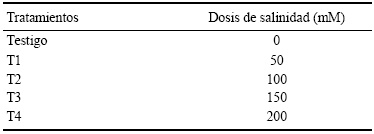
Por lo anterior, se realizó el trabajo de investigación cuyo objetivo fue determinar la respuesta diferencial en cuanto a su capacidad de germinación y características morfológicas de ocho genotipos de tomate, que fueron sometidos a diferentes concentraciones salinas (0, 50, 100, 150 y 200 mM de NaCl) en sus primeras etapas fenológicas (germinación y crecimiento vegetativo inicial).
Materiales y métodos
Área de Estudio
El presente trabajo de investigación se realizó en el Centro de Investigaciones Biológicas del Noroeste, S.C. (CIBNOR), el cual se localiza en los terrenos costeros de El Comitán, en la porción meridional de la península de Baja California Sur, a 24° 08' N y 110° 24' O, 17 km al oeste del puerto de La Paz, capital del estado de Baja California Sur, México.
Material Genético Utilizado
Para ambas etapas fenológicas en estudio (germinación y crecimiento vegetativo inicial), se utilizaron ocho cultivares comerciales de tomate tipo "Saladette: Missouri, Super, Río Grande, Yaqui y tipo bola: Tropic, Feroz, Ace, Vita y Floradade".
Etapa de Germinación
La evaluación de los genotipos en la etapa de germinación se desarrolló previa a la aplicación de los tratamientos salinos, el experimento inició sometiendo a los genotipos mencionados a los tratamientos con NaCl (Cuadro 1) con cuatro repeticiones de 100 semillas cada una, en un diseño experimental completamente al azar con arreglo factorial, considerando a los genotipos como el factor A y a los tratamientos salinos como el factor B. Posterior a esta prueba y una vez que se determinó la viabilidad y el porcentaje de germinación de todos los genotipos, las semillas se colocaron en cajas Petri utilizando papel filtro como sustrato. En la primera aplicación de los tratamientos, a cada caja Petri se le suministraron 25 mL de solución salina correspondiente a cada tratamiento, manteniendo así la humedad del sustrato. Este experimento se realizó en una cámara de germinación marca Lumistell, modelo IES-OS, SERIE 1408-88-01 en condiciones controladas de temperatura (25±1 °C), humedad (80%) y con luz (12 horas continuas) durante los 14 días del periodo de evaluación.
Variables de Respuesta Considerados en la Etapa de Germinación
Porcentaje de germinación. Se realizó una revisión diaria de cada una de las cajas Petri, a partir del tercer día, realizando el conteo diario por un periodo de 14 días, tiempo límite para la germinación de semillas de tomate (Moreno, 1984). Las variables morfométricas medidas en las plántulas se realizaron inmediatamente al cumplir los 14 días de evaluación de la germinación.
Peso fresco de parte aérea (tallos + hojas). Se determinó al dividir cada plántula en tallos y hojas y pesarlas por separado, utilizando para ello una balanza analítica (Marca Mettler Toledo, modelo AG204). Posteriormente se sumaron ambos pesos, mismos que fueron expresados en gramos de materia vegetal fresca. Cabe señalar que para las variables peso fresco y seco de parte aérea y peso fresco y seco de raíz se realizó con base en la metodología propuesta por Murillo et al. (2007).
Peso seco de parte aérea (tallos + hojas). Una vez que se obtuvo el peso fresco de tallos y hojas de las plántulas, ambos tejidos se colocaron en una estufa de secado (Marca Shel-Lab, modelo FX-5, serie-1000203) a una temperatura de 80 °C durante 72 h hasta obtener su deshidratación completa. Posteriormente se pesaron en balanza analítica (Marca Mettler Toledo, modelo AG204) y el peso fue expresado en gramos de materia vegetal seca (Murillo et al., 2007).
Peso fresco y seco de raíces. Esta variable fue evaluada tomando el peso de la masa radicular y el resultado se expresó en gramos, utilizando para ello una balanza analítica (Marca Mettler Toledo, modelo AG204). Una vez que se obtuvo el peso fresco de raíces, éstas se colocaron en bolsas de papel y se introdujeron en una estufa de secado (Marca Shel-Lab, modelo FX-5, serie-1000203) a una temperatura de 80 °C durante 72 h hasta obtener su deshidratación completa. Posteriormente se pesaron en balanza analítica (Marca Mettler Toledo, modelo AG204), expresando el peso en gramos de materia vegetal seca (Murillo et al., 2007).
Plántulas normales. En la prueba de germinación se considerarón plántulas normales después de los 14 días de la prueba establecida según Moreno (1984) y fueron aquellas que presentaron las siguientes estructuras esenciales:
sistema radicular bien desarrollado, incluyendo raíz primaria y
las que presentaron una plúmula intacta con desarrollo vigoroso y balanceado.
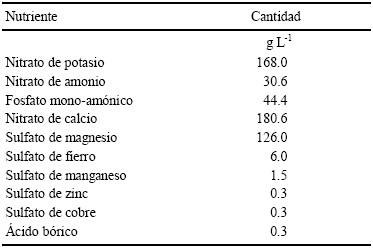
Etapa de crecimiento vegetativo inicial. Esta segunda etapa se realizó en condiciones de malla sombra. Las semillas se sembraron en charolas de poliestireno de 200 cavidades, las cuales contenían sunshineMR (sustrato comercial INERTE). El riego aplicado a las charolas se realizó diariamente con el fin de lograr una emergencia homogénea de las plantas. El trasplante se realizó cuando las plantas presentaron una altura promedio entre 10 y 15 cm en macetas de aproximadamente 4 kg, mismas que contenían como sustrato una mezcla en una proporción de 1:1 de arena gruesa de arroyo más sunshineMR. En cada maceta se colocaron tres plantas con el fin de asegurar el éxito del trasplante. Una vez que se trasplantaron, se inició con la aplicación diaria del riego, se utilizó agua potable, la cual contenía una solución nutritiva (Cuadro 2) que fue preparada para tomate según Samperio (1997). Después de 8 días del trasplante se inició con la aplicación de los tratamientos. Cada vez que se aplicaba, se tomaba la lectura del pH y de la conductividad eléctrica de cada tratamiento, de la solución drenada por las macetas para comparar el pH y la conductividad eléctrica de cada solución. La solución nutritiva se ajustó a un pH de 6.5, adicionando H2SO4 o KOH, para su uso en el riego.
Cuadro 2 Solución nutritiva utilizada en el experimento de crecimiento vegetativo inicial de plantas de tomate (Tomado de Samperio, 1997).
Aplicación del riego. Los tratamientos 1 y 2 (0 y 50 mM de NaCl) continuaron con su respectiva solución, a los tratamientos 3, 4 y 5, se les aplicó el tratamiento salino correspondiente a 100 mM de NaCl. Los siguientes tres días a esta aplicación, los tratamientos 1, 2 y 3, mantuvieron la concentración salina asignada para cada uno de ellos, mientras que a los tratamientos 4 y 5 se les aplicó el tratamiento de 150 mM de NaCl. Los tres días siguientes los tratamientos 1, 2, 3 y 4 mantuvieron las concentraciones de NaCl asignadas al experimento y por último en los siguientes tres días, al tratamiento 5 se le aplicó la concentración de 200 mM de NaCl, acorde con la metodología propuesta por Murillo et al. (2007). La cantidad aplicada en cada riego fue de 500 mL, con lo que la solución aplicada drenó a través de los orificios de las macetas, con la finalidad de evitar la acumulación de sales en el sustrato.
Longitud del tallo, peso fresco y seco. Esta variable se midió después de que las plantas se cosecharon al cumplirse el período de 35 días de aplicación de los tratamientos salinos. Se les separó la raíz, tallo y hojas. La longitud del tallo consistió en medirlo desde su base hasta la parte apical. Esta variable está expresada en centímetros (Murillo et al., 2007).
Posteriormente se determinó su peso fresco, expresando los resultados en gramos. Cada tallo se introdujo en bolsas de papel y se colocaron en una estufa de secado (Shel-Lab, modelo FX-5, serie-1000203) a una temperatura de 80 °C durante 72 h, con el fin de deshidratar completamente el material vegetativo. Después de sustraerlos de la estufa, se pesaron en una balanza de precisión (Mettler Toledo PR2002), expresando los datos en gramos.
Número de hojas. Se contabilizaron todas las hojas verdaderas de cada planta cosechada.
Área foliar. Después de separar las hojas de los tallos, se determinó el área foliar total utilizando un integrador de área foliar (LI-COR, modelo-LI-3000A, serie PAM 1701), expresando los datos de esta variable en centímetros cuadrados.
Peso fresco de hojas. Se determinó utilizando una balanza de precisión (Mettler Toledo PR2002). Los datos se expresaron en gramos.
Análisis estadístico. Se realizaron análisis de varianza para todas las variables medidas en las dos etapas fenológicas, utilizando el paquete estadístico STATISTICA (StatSoft Inc., 2001), cuando se encontraron diferencias significativas entre tratamientos (genotipos y salinidad), se realizaron pruebas de comparación de medias basadas en Tukey (P ≤ 0.05). Con el fin de cumplir con los supuestos de homogeneidad de varianza, se realizaron transformaciones de datos cuando fue necesario. La variable porcentaje de germinación se transformó utilizando arcoseno, la variable número de plantas anormales se transformó por el método de la raíz cuadrada de X (√X) y la variable número de semillas muertas se transformó por el método de log (Y+1) (Sokal y Rohl, 1988). Además se realizó análisis de regresión lineal entre los niveles de salinidad y las variables en estudio.
Resultados y discusión
Etapa de Germinación
La salinidad afecta negativamente la germinación de las semillas de tomate de plantas cultivadas. Estos efectos inciden en el porcentaje de germinación y el tiempo en que este proceso se lleva a cabo (Goykovic y Saavedra, 2007). Según Cuartero et al. (2006) en la evaluación de la heredabilidad de diferentes caracteres en plantas de Solanum lycopersicum x S. pimpinellifolium expuestas a condiciones salinas, encontraron que al interior del género Lycopersicon, especies con un grado de tolerancia a salinidad mayor que otras, incluyendo a L. esculentum, hecho que demuestra que no sólo al interior de las especies silvestres es posible detectar germoplasma con mayor tolerancia a salinidad. Los resultados obtenidos en la investigación dan cuenta de la existencia de variabilidad genética en tomate y por tanto de germoplasma importante de considerar para el mejoramiento genético a estrés salino, de modo que los esfuerzos en identificar germoplasma resistente a este estrés son válidos, especialmente en aquellas especies silvestres que presenten compatibilidad genética con los cultivares comerciales, en este sentido Casierra et al. (2013) resaltan que no sólo los caracteres relacionados con el rendimiento son importantes en los estudios de mejoramiento de plantas tolerantes a la salinidad, debido a que la salinidad afecta a casi todos los aspectos fisiológicos y bioquímicos de la planta, por lo que el incremento de la tolerancia de los cultivos a la salinidad requiere la combinación de algunos o muchos rasgos fisiológicos, y no simplemente aquellos que influyen directamente en el rendimiento.
Para el presente estudio, los resultados indican que, la mayoría de las variables medidas en la etapa de germinación, mostraron una tendencia lineal al disminuir conforme los niveles de salinidad se incrementaron (Figura 1 y 2).
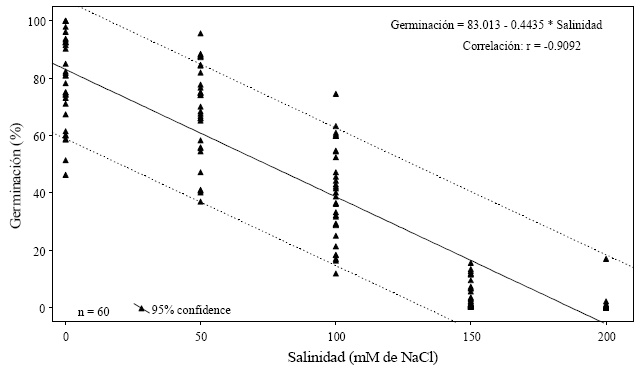
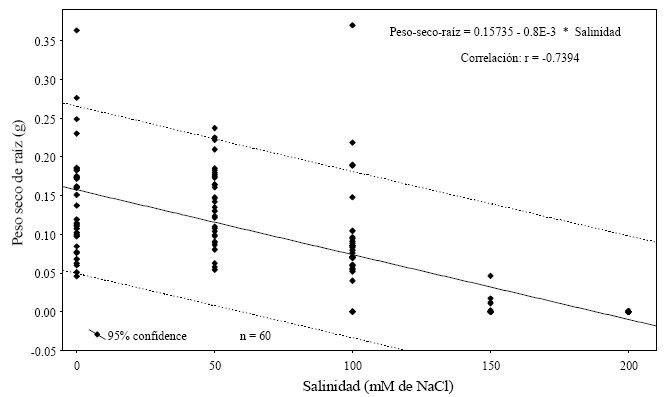
Figura 2 Efecto de la salinidad en el peso seco de raíces de plántulas de ocho genotipos de tomate en la etapa de germinación.
Así mismo, los resultados concuerdan con lo mencionado por Jones (1986), quien reporta que la tolerancia a la salinidad de las semillas en su germinación es una medida de la habilidad de éstas para soportar los efectos de altas concentraciones de sales solubles en el medio, ya que la presencia de las mismas disminuye el potencial hídrico, lo que provoca una menor disponibilidad de agua para las semillas. De manera que deben generar suficiente potencial osmótico para mejorar el estatus hídrico de los embriones y permitir su crecimiento, se observó que los genotipos "Tropic y Yaqui" tuvieron valores superiores en germinación, incluso en las concentraciones de 50 y 100 mM de NaCl y en 150 para "Tropic", en comparación con los genotipos Ace y Vita, esto se debe a que la salinidad origina reducción del crecimiento de los cultivos al afectar negativamente la germinación o la capacidad de emerger de las plántulas. En el Cuadro 3 se muestra el genotipo "Missouri" que mostró el valor superior en 200 mM de NaCl, aunque este valor no llegó al 6%.
Acorde a la etapa de desarrollo del cultivo, los resultados obtenidos concuerdan con los encontrados por Blanco et al. (1991) en tomate al aplicar Na Cl en el agua de riego, mencionan que la germinación disminuye a medida que se incrementan los niveles de salinidad, por su parte Camejo y Torres (2000) reportan que al aplicar dosis de 150 mM de sales de sodio la germinación en semillas de tomate disminuyó, en este mismo sentido Cristo et al. (2001) mencionan que en cultivares de arroz al incrementar los niveles de salinidad su germinación decrece.
Cuadro 3 Efecto de la salinidad en la germinación, peso fresco y seco de parte aérea, peso fresco de raíz y peso fresco de hojas de ocho genotipos de tomate.
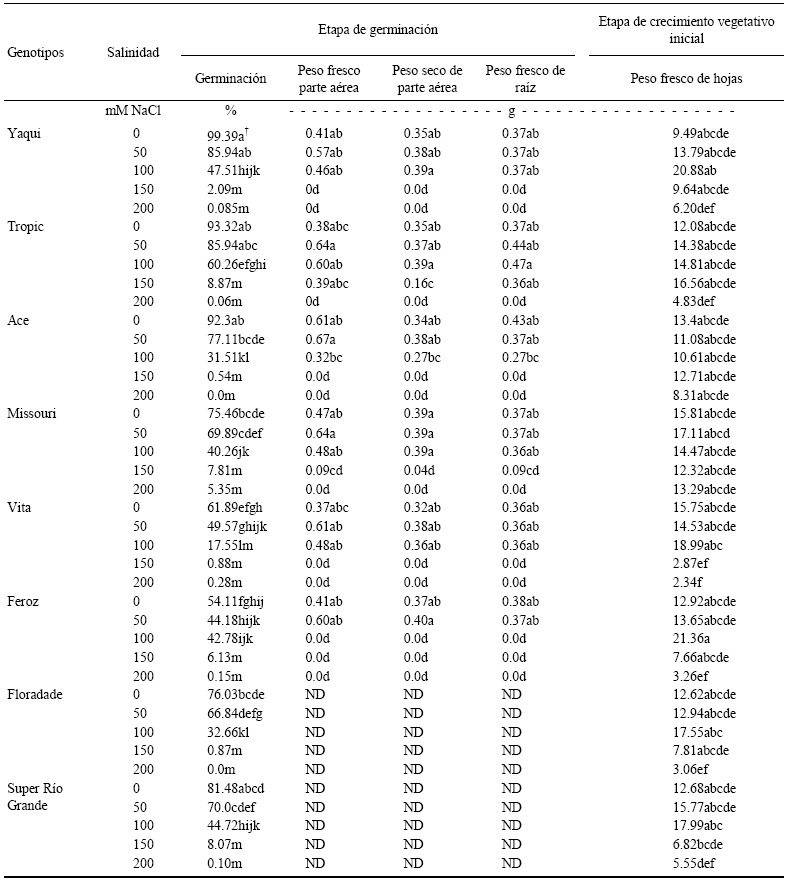
† Literales diferentes entre columnas indican desigualdad (P ≤ 0.05). ND = no determinado por problema técnico del experimento.
La variable peso fresco de parte aérea se incrementó en todos los genotipos de 0 a 50 mM de NaCl, para disminuir a partir de 100 mM conforme los niveles de salinidad se incrementaron. Asimismo, la variable peso seco de parte aérea incrementó en todos los genotipos de 0 a 50 mM y en algunos genotipos "Yaqui y Tropic" se incrementó de 50 a 100 mM o se mantuvo con el mismo valor en 50 y 100 mM "Missouri", para disminuir al incrementarse los niveles de salinidad (Cuadro 3). El peso fresco de raíz, se mantuvo con valores similares en las concentraciones 0, 50 y 100 mM en los genotipos "Yaqui y Vita"; en el genotipo "Tropic" se incrementó al aplicar 50 mM; en el genotipo "Ace", disminuyó a 50 mM, mientras que en el genotipo Missouri se mantuvo con el mismo valor en 0 y 50 mM pero disminuyó al cambiar la concentración de sales de 50 a 100 mM. Los resultados anteriores indican que a medida que la salinidad aumenta, se afecta la absorción de agua por las semillas y su germinación, así como el crecimiento de las plántulas. Esta respuesta coincide con lo determinado por Fraschina y Chiesa (1993); González y Ramírez (1996), quienes mencionan que tales resultados se deben al efecto osmótico que causa el exceso de sales en la solución y que dificulta la absorción de agua, así mismo, al efecto tóxico de los iones de Na y Cl, que causan desbalance nutricional y afectan el metabolismo o la combinación de ambos efectos.
Etapa de Crecimiento Vegetativo Inicial
Todos los órganos de la parte aérea de las plantas de tomates se alteraron con la salinidad. Los tallos alcanzaron menor altura, las hojas disminuyeron en número, el área foliar igualmente se redujo, se registró un menor peso fresco y seco del área foliar, tallo y raíces (Cuadro 3).
Los resultados coinciden con lo mencionado por Maas y Hoffman (1976); Aceves (1979); Rowell (1989); Alvarado et al. (1991); Escalante (1995); Carvajal et al. (1999); y Lombardo y Saladino (1997) quienes reportan disminuciones en el área foliar, altura de la planta, peso fresco y seco del área foliar, tallo y raíz y número de hojas al incrementarse la concentración de la salinidad en el agua de riego. Sin embargo, en el presente trabajo el peso fresco de las hojas se incrementó al aumentar la concentración de sales de 0 a 50 y de 50 a 100 mM en las variedades "Yaqui, Tropic, Feroz, Floradade y Super Río Grande" y posteriormente disminuyó en los niveles de 150 y 200 mM de NaCl, estos resultados posiblemente se deben a que estos genotipos tienen cierta tolerancia en las primeras etapas de crecimiento, tal y como lo señalan González et al. (2002) quienes mencionan que en la evolución de los mecanismos de tolerancia y adaptación de las plantas a los diferentes agentes estresantes, puede observarse la existencia de grados de sensibilidad y de tolerancia muy diferentes entre las diversas especies e incluso variedades o ecotipos dentro de una misma especie, lo que evidencia la diversidad de estrategias que han desarrollado las plantas, para mantener una respuesta altamente refinada ante una amplia gama de estrés, a las cuales se encuentran rutinariamente expuestas. En el Cuadro 3 se muestran otros genotipos (Ace y Vita) que disminuyeron en peso fresco de hojas, al variar la concentración de 0 a 50 mM pero se incrementó al pasar de 50 a 100 mM, mientras que en el genotipo "Missouri", se incrementó ante el cambio de 0 a 50 y disminuyó de 50 a 100 mM de NaCl. Estos resultados coinciden a los reportados por Akran et al. (2010) quienes establecen que en presencia de salinidad, los cultivos responden de manera particular, algunos producen rendimientos aceptables a altas concentraciones de salinidad, mientras que otros son sensibles a bajas concentraciones. Este incremento en los valores de biomasa seca a concentraciones moderadas y relativamente elevadas puede ser debido a un incremento en la síntesis de solutos orgánicos (azúcares, prolinas, amino ácidos) para contrarrestar los efectos osmóticos de la salinidad en esta etapa del desarrollo, lo que puede estar asociado con la presencia de mecanismos de tolerancia a la salinidad en el cultivar.
En el mismo contexto se ha señalado que las plantas en condiciones de salinidad (Balibrea et al., 1996), para ajustarse osmóticamente e incrementar su potencial osmótico interno utilizan una porción de sus fotosintatos. Por su parte Morales et al. (2010), encontraron que las plantas de tomate tratadas con niveles moderados de salinidad (50 mM) presentaron un crecimiento de biomasa similar o superior al control en correspondencia con un incremento en el contenido de azúcares reductores, totales y de prolina en tallo y raíz.
Estos últimos resultados señalan que algunos genotipos son fuentes de genes de gran importancia para el mejoramiento de esta hortaliza para resistir a condiciones salinas. Al igual que en la etapa de germinación, algunas variables medidas en la presente etapa, mostraron una tendencia lineal al disminuir sus valores conforme los niveles de salinidad se incrementaron (Figura 3). Los resultados obtenidos coinciden con Goykovic y Saavedra, (2007) que mencionan que la salinidad produce efectos negativos en las plantas de tomates, sean estas cultivadas o silvestres. La mayoría de los efectos son negativos, y se expresan desde los primeros estados fenológicos de la planta. La germinación se reduce y se prolonga el tiempo de esta etapa. El crecimiento de las raíces disminuye y por tanto se reduce la capacidad de absorción de agua y nutrimentos.

Figura 3 Efecto de la salinidad en la longitud de tallo de plántulas de ocho genotipos de tomate en la etapa de crecimiento vegetativo inicial.
Por otro lado, la Figura 4 muestra el efecto de las diferentes concentraciones salinas sobre el peso seco de tallo, observándose que los valores promedio mayores para esta variable, se presentaron en la concentración de 0 mM de sales, seguido en orden descendente por 50, 100, 150 y 200 mM de NaCl. Es decir, la tendencia de esta variable fue disminuir conforme los niveles de salinidad se incrementaron, esta respuesta puede ser explicada mediante una ecuación de tipo lineal donde se obtuvieron valores de significancia (r = -0.47, R2 = 0.22, n = 120, P = 0.000001) que revelan el ajuste de los datos a este modelo.

Figura 4 Relación entre la salinidad (NaCl) y el peso seco de tallo de plantas de ocho genotipos de tomate en la etapa de crecimiento vegetativo inicial.
La disminución de las variables relacionadas con el tallo y su morfometría en respuesta a la salinidad coincide con lo expuesto por García y Jáuregui (2008) y Madueño et al. (2006), donde establecen que uno de los primeros efectos fisiológicos que provoca el estrés salino en las plantas es la reducción del crecimiento debido a una disminución en la capacidad de absorción de agua de estas; en este mismo contexto Cruz y Cuartero (1990), establecen que la habilidad de adaptarse a la salinidad parece ser mayor en plantas más maduras o viejas que en plantas jóvenes o plántulas, se ha demostrado que los tomates desarrollados mediante la exposición al agua salina a través de su vida, disminuyen, en peso seco de parte aérea, en la floración y en el tiempo de inicio de cosecha con respecto al control. De acuerdo con estos mismos autores, tanto el tallo como la hoja de tomate disminuyen en condiciones salinas, aunque en cultivares de L. esculentum, el tallo, expresado en peso seco (biomasa) disminuye en menor proporción que el peso seco de la hoja.
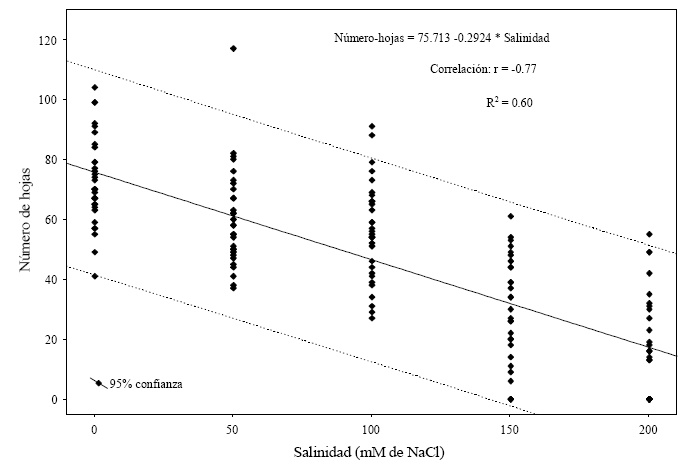
El efecto de las concentraciones salinas en el número de hojas en plantas de tomate durante la etapa de crecimiento vegetativo inicial (Figura 5), fue significativamente diferente entre la concentración de 0 mM de NaCl y las concentraciones de 50, 100, 150 y 200 mM de NaCl. En relación a las concentraciones de 50 y 100 mM de NaCl estas no difieren estadísticamente entre sí, pero en comparación con las concentraciones de 0, 150 y 200 Mm si se presentó diferencias significativas en el número de hojas. Acorde con lo reportado por Wilches et al. (2008), la cantidad de fotosíntesis que una planta realiza depende de la superficie de la hoja u órganos fotosintéticos que posea y de la actividad fotosintética por unidad de área de estos tejidos. Al mismo tiempo, el área foliar depende del número de hojas, de su velocidad de crecimiento y de su tamaño final (Lovelli et al., 2012). La respuesta de esta variable al efecto de la salinidad, mostró una tendencia decreciente conforme los niveles de salinidad se incrementaron (Figura 5), dicha respuesta se puede explicar mediante una ecuación de tipo lineal donde se obtuvieron valores de significancia (r = -0.77, R2 = 0.60, n = 120, P = 0.000001) que revelan el ajuste de los datos a este modelo.
Área Foliar
En la respuesta diferencial del área foliar de plantas se encontró que el genotipo Missouri presentó un valor promedio superior en comparación a los demás, seguido en orden descendente por los genotipos "Feroz, Yaqui, Ace, Tropic y Río Grande", los cuales se agruparon con valores estadísticos similares. Los genotipos "Vita y Floradade" por su parte, se agruparon con los valores menores para esta variable (Figura 6). Los resultados obtenidos en la presente investigación demostraron que la respuesta diferencial de los genotipos evaluados, es una característica de los cambios que se observan en condiciones de salinidad en el área foliar, la cual se reduce, afectando también la relación parte aérea/raíz (Maurel y Chrispeels, 2001). Estos resultados coinciden a lo mencionado anteriormente.

Figura 6: Respuesta diferencial del área foliar de plantas de ocho genotipos de tomate sometidas a estrés por salinidad (NaCl) en la etapa de crecimiento vegetativo inicial. Barras con misma literal no difieren estadísticamente (Tukey P ≤ 0.05).
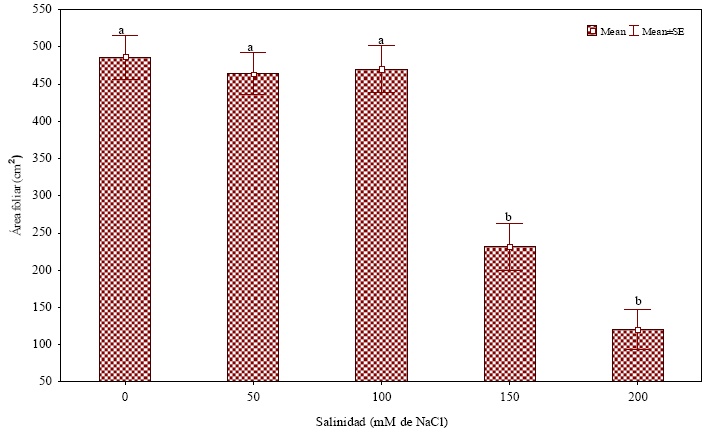
El promedio de área foliar de las plantas sometidas a las concentraciones de 0, 50 y 100 mM de NaCl no difieren estadísticamente (Figura 7). Por su parte, las plantas sometidas a concentraciones de 150 y 200 mM de NaCl no mostraron diferencia estadística entre sí, respecto al área foliar, presentando los valores más bajos. Los resultados obtenidos en cuanto al efecto de las concentraciones salinas mayores (150 y 200 mM) coinciden con lo expresado por Hasegawa et al. (2000) quienes mencionan que uno de los efectos más evidentes del estrés salino es la reducción en la capacidad de absorción de agua, que se puede manifestar en una reducción de expansión foliar y pérdida de turgencia en tallo y hojas de las plantas. Asimismo, las causas de la salinización ocasionan una caída repentina del potencial de agua de la hoja, el cual no es inmediatamente contabilizado por la disminución lenta del potencial osmótico de la hoja. En salinidades relativamente bajas, esto puede resultar en una reducción transitoria (quizás por unos minutos) de la turgencia y la tasa de crecimiento foliar (Sacher y Staples, 1985; Yeo et al., 1991, Munns et al., 2005). De la misma manera Romero et al. (2001) mencionan que a nivel de hojas la salinidad genera una reducción en su número y en el área foliar.
Conclusiones
La salinidad produce efectos negativos en la germinación y el crecimiento inicial de las plántulas de tomate, sugiere que la salinidad afecta de forma diferenciada según el cultivar, jugando un papel importante la variabilidad genética y la etapa del desarrollo de las plantas, siendo las etapas juveniles más sensibles que las etapas adultas.
Los cultivares "Missouri y Tropic" mostraron mayor tolerancia a salinidad, siendo en las concentraciones de 0, 50 y 100 mM donde se presentó mejor desarrollo de la altura de planta, área foliar, peso fresco y seco del área foliar y la germinación.
Las concentraciones elevadas de NaCl inhiben la germinación, el crecimiento y desarrollo de las plantas de tomate, principalmente de los genotipos "Feroz y Vita" en crecimiento inicial.
El trabajo experimental debe evaluarse en campo para corroborar la tolerancia a la salinidad en las diferentes etapas fenológicas de los variados genotipos del cultivo de tomate.

