











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La tecnología de los sensores remotos, en plataformas espaciales o a nivel terrestre, muestra un alto potencial para determinar el estado nutrimental de la vegetación. Esto implica un escalamiento de las reflectancias, obtenidas en los sensores remotos, desde el nivel de parcela, rodal o píxel al de constituyentes bioquímicos en células de las hojas. La tarea no es fácil y no puede ser determinada en forma directa.
Al considerar que los procesos ecológicos que intervienen en la transferencia de energía y masa entre el sistema suelo-vegetación-atmósfera (fotosíntesis, evapotranspiración, respiración, producción primaria y descomposición), están relacionados con el estado nutrimental y factores de crecimiento de la vegetación (Peterson y Hubbard, 1992), las concentraciones de clorofilas, agua, proteína, celulosa y lignina; principalmente, definen estos acoplamientos. La estimación de las concentraciones bioquímicas de las hojas es fundamental para obtener parámetros que permitan caracterizar y modelar los procesos ecológicos. Por ejemplo, la descomposición del mantillo orgánico sobre el suelo está gobernada por los contenidos foliares de lignina y nitrógeno (Melillo et al., 1982). La disponibilidad de nitrógeno en la vegetación condiciona fuertemente la producción primaria neta (Vitousek, 1982) y la evapotranspiración por el acoplamiento fotosíntesis y transpiración (Dickinson et al., 2002). La relación carbono-nitrógeno (C/N) es un parámetro básico en la dinámica de los procesos ecológicos. El nitrógeno tiene una relación lineal estable con la proteína en las hojas; lo mismo sucede con el carbono para la lignina y la celulosa (Jacquemoud et al., 1996), por lo que se pueden usar en términos intercambiables.
La estimación de las concentraciones bioquímicas usando reflectancias R o absortancias A [aproximadas por log(R-1)] ha sido evaluada en términos estadísticos (Curran, 1989; Curran et al., 1992; Jacquemoud et al., 1995b; Fourty y Baret, 1998), usando generalmente técnicas de regresión lineal por pasos para seleccionar bandas espectrales optimas que se correlacionen fuertemente con las concentraciones bioquímicas. Aunque los resultados obtenidos por las técnicas estadísticas han sido buenos (clorofilas, agua, proteína, celulosa y lignina) (Curran, 1989), las bandas espectrales definidas no coinciden con los picos de absorción de los constituyentes bioquímicos y no son estables al usarse en un conjunto amplio de espectros de hojas (Grossman et al., 1996). Ante esto, se ha planteado el uso de modelos radiativos para estimar las concentraciones bioquímicas (Jacquemoud et al., 1995b; Grossman et al., 1996).
La modelación de la transferencia radiativa se ha desarrollado para hojas anchas (Jacquemoud et al., 1990, 1995c y 1996) y aciculares (Dawson et al., 1998). La discusión en lo siguiente está focalizada en hojas anchas, aunque los desarrollos son válidos para hojas aciculares (que incluyen un factor de escala de agrupamiento de las acículas en los racimos; Smolander y Stenberg, 2005).
La estimación de las concentraciones bioquímicas foliares ha sido aproximada al utilizar índices espectrales (le Maire et al., 2004), con resultados poco satisfactorios para ser usados en un conjunto más amplio de datos bioquímicos-espectrales que los de calibración. Los modelos radiativos foliares se han invertido con más o menos éxito (Fourty et al., 1996), como consecuencia del esquema de calibración usado en estos modelos. En el caso del follaje de la vegetación, se han utilizado con poco éxito modelos radiativos de medios turbios (homogéneos) acoplados a modelos de las hojas para invertirlos (Jacquemoud et al., 1995a, 2000); ya que las estimaciones son dependientes del acoplamiento bioquímico-estructural del follaje. También, se han realizado algunos esfuerzos usando índices espectrales de la vegetación (Broge y Leblanc, 2000) con resultados similares. En el caso de medios radiativos heterogéneos (tri-dimensionales) (Gastellu-Etchegorry y Bruniquel-Pinel, 2001), los problemas del acoplamiento bioquímico-estructural han estado presentes, limitando las estimaciones.
En la actualidad, usando el marco teórico de los invariantes espectrales (Huang et al., 2007), Lewis y Disney (2007) y Knyazikhin et al. (2013) demostraron en términos más o menos generales que, dado el acoplamiento de las propiedades estructurales del follaje (índice de área foliar o IAF y cobertura o fv) con la bioquímica foliar, no es posible realizar estimaciones robustas de ambas propiedades, sin importar lo angosto de las bandas espectrales usadas o el número de ellas. Cualquier estimación que se realice de ambas propiedades contendrá un error muy grande, además de ser espuria (Knyazikhin et al. 2013), y representa un problema matemáticamente indeterminado (muchas soluciones al mismo problema). Así, la perspectiva de estimar las propiedades bioquímicas de las hojas usando datos espectrales del follaje de la vegetación parece tener una limitación fundamental e intrínseca.
En este trabajo se presenta un esquema de modelación que permite evitar la restricción establecida por Lewis y Disney (2007) y Knyazikhin et al. (2013), de tal forma que para estimar las concentraciones bioquímicas se pueden invertir de forma directa los modelos radiativos foliares. Considerando la accesibilidad de la información espectral usando sensores remotos, se establece un enfoque de modelación radiativa foliar acoplado a los sensores remotos, que permite evitar problemas de calibraciones independientes de las componentes radiativas del albedo foliar. Al final se muestra la relación de los desarrollos con la teoría de invariantes espectrales.
Materiales y métodos
Conversión de Medios Radiativos Heterogéneos a Homogéneos
El caso de la evolución temporal del cubrimiento aéreo de un cultivo sembrado en surcos se muestra en la Figura 1, para ejemplificar la situación de como diferentes aproximaciones teóricas de modelación (desde un esquema unidimensional a uno tridimensional de transferencia radiativa) son requeridas. Al principio de la etapa de crecimiento se observa un suelo desnudo (caso 1-D) y después un patrón de plantas individuales, donde las plantas no se tocan entre sí (caso 3-D). Al avanzar el crecimiento, las plantas en un mismo surco se tocan entre sí y forman un cultivo en hileras o surcos (caso 2-D). Cuando las plantas de diferentes surcos se tocan, tenemos la situación de cobertura total del suelo (caso 1-D). Esta dinámica de crecimiento requiere de un esquema de modelación que haga equivalentes, en términos radiativos, a las diferentes geometrías observadas en la Figura 1, donde muestran medios homogéneos que pueden ser aproximados por medios radiativos turbios (Figura 1a y 1d).
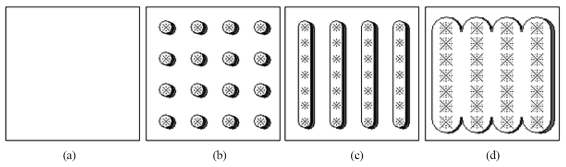
Figura 1 Geometría asociada a un cultivo sembrado en surcos. (a) suelo desnudo; (b) plantas individuales sin traslape; (c) plantas traslapadas en surcos; y, (d) vegetación densa traslapada.
Para focalizar las discusiones, en lo siguiente se usara R para denotar la reflectancia en la banda del rojo e IRC para la banda del infrarrojo cercano.
El espacio R-IRC asociado con la dinámica del crecimiento de la vegetación (medio turbio) representada por el índice de área foliar se muestra en la Figura 2a, donde se usaron cuatro suelos con propiedades ópticas (reflectancias) diferentes (Paz et al., 2005). En esta figura las líneas iso-IAF (IRC = a0 + b0R) representan la misma cantidad de vegetación, pero con suelos ópticamente diferentes; donde el caso de IAF = 0 representa la línea del suelo desnudo (IRC = aS + bSR). Si el suelo (reflectancia) permanece constante, entonces el crecimiento de la vegetación sigue una curva iso-suelo. El atrayente de la dinámica del crecimiento de la vegetación para la banda del R es el punto de convergencia de todas las curvas iso-suelo. En este punto la banda del R se satura (no cambia de valor al incrementarse el IAF) y representa a un medio ópticamente denso o infinito (Ross, 1981), por lo que se le denomina como R∞. El valor del IRC asociado al R de saturación es IRCR∞. banda del IRC sigue creciendo hasta alcanzar su propio punto de saturación definido por IRC∞. Las reflectancias en el infinito están en función de las propiedades ópticas de las hojas y su distribución angular, y geometría sol-sensor, principalmente (Ross, 1981; Zarco et al., 2008; Paz et al., 2009a y 2013). Las concentraciones de los constituyentes bioquímicos foliares están en función de las propiedades ópticas de las hojas (reflectancia y transmitancia) (Jacquemoud et al., 1996), por lo que determinadas éstas es posible realizar estimaciones desacopladas de los parámetros estructurales del follaje, puesto que la reflectancia en el infinito es invariante por efecto del IAF o fv (los valores superiores de estos parámetros no cambian las reflectancias en el infinito).
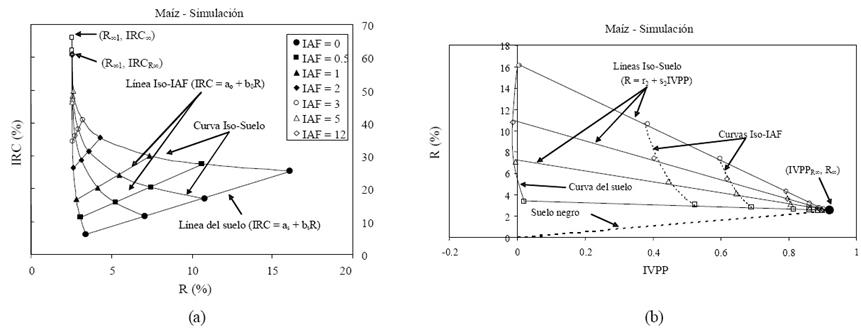
Figura 2 Dinámica espectral del crecimiento de la vegetación. (a) Espacio R-IRC y (b) espacio IVPP-R.
Una forma de evitar los patrones no lineales de las curvas iso-suelos de la Figura 2a es usar el espacio espectral IVPP-R (Paz et al., 2009a y 2013), donde IVPP = dIRC/IRC (IVPP = índice de vegetación de proporciones pareadas; Paz et al., 2009a y b), Figura 2b, con dIRC = IRC - (aS + bSR) (Paz et al., 2005 y 2006).
En el espacio IVPP-R, las líneas iso-IAF son curvas no lineales (Figura 2b) contrastando con la linealidad de las curvas iso-suelo (R = r2 + s2IVPP). Por definición de utilizar a R sin transformación, el uso de cualquier valor de los parámetros aS y bS de la línea del suelo no cambia la estimación de R∞ y solo los valores de I IVPPR∞ (Figura 3), ya que la intersección del conjunto de líneas iso-suelo en la figura permanece invariante en términos de la banda del R.

Usando el espacio IVPP- R podemos estimar R∞ para poder usarlo en la estimación del albedo de las hojas. El caso de la estimación de IRC∞ requiere de un esquema de estimación diferente y se discute en otros trabajos (Casiano et al., 2012 y Paz et al., 2013). El método de estimación del R( es válido para las bandas del verde o V y azul o A del espectro visible, así como para los infrarrojos medios (Paz et al., 2009b y 2013). El procedimiento es general, ya sea que la curva del suelo sea lineal o no lineal e independiente de sus parámetros (Paz et al., 2009b).
Estimación del Albedo Foliar en Medios Homogéneos
Los atrayentes (puntos de convergencia de las líneas o curvas iso-suelo) de la dinámica del crecimiento de los cultivos asociada a los espacios espectrales analizados en la sección anterior, (R∞, IRCR∞) y (R∞, IRC∞) permiten estimar las propiedades ópticas y geométricas del follaje (medio equivalente con parámetros efectivos) al convertir un medio heterogéneo (reflectancias no en el infinito) a uno homogéneo (caracterizado por reflectancias en el infinito) (Paz et al., 2009a y 2013)
La reflectancia en el infinito para hojas no horizontales, puede aproximarse razonablemente (Goudriaan y van Laar, 1994) usando la relación:
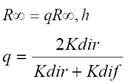 (1)
(1)
donde: K es un coeficiente de extinción, dir se refiere a radianza directa, dif a radianza difusa y h a hojas con distribución angular horizontal. R se refiere a cualquier banda espectral.
Para evaluar el coeficiente de extinción Kdir es necesario conocer la distribución de probabilidad angular de las hojas (Ross, 1981):
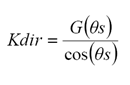 (2)
(2)
donde: θs es el ángulo cenital de iluminación y G(θs) es la proyección del área foliar de las hojas en la dirección θs (Ross, 1981).
Usando el modelo de radiación difusa de "cielo nublado estándar" (Moon y Spencer, 1942), Sinoquet et al. (2000) usaron datos obtenidos por Bonhomme y Varlet-Grancher (1977) para generar la relación:
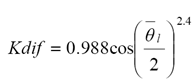 (3)
(3)
donde: es el ángulo promedio de las hojas.
Goudriaan (1977) y Ross (1981) propusieron una serie de simplificaciones para estimar la reflectancia en el infinito para hojas horizontales. Así, para hojas horizontales, con ρ = τ, donde ρ es la reflectancia foliar y τ la transmitancia, que es una buena aproximación (Gausman et al., 1973 y Ross, 1981) y bi-Lambertianas (isotrópicas), la reflectancia en el infinito está dada por:
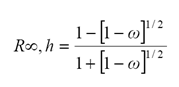 (4)
(4)
donde: ω (= ρ+τ) es el albedo foliar simple.
De esta forma, conocidos los coeficientes de extinción, podemos invertir las relaciones 1 y 4 para obtener el albedo foliar, conocida la reflectancia en el infinito, como:
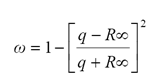 (5)
(5)
La relación 5, requiere de conocer la distribución angular de las hojas, y podemos estimarla en forma indirecta usando propiedades de invarianza asociadas a la geometría sol-sensor (Paz et al., 2009a). Todas las distribuciones de probabilidad angulares de las hojas tienen un valor aproximado de G(θs) ? 0.5 cuando θs ? 57 (ángulo de visión a nadir) (Ross, 1981). Así, bajo esta condición, aproximada por una distribución angular esférica, se obtiene el valor q = 1.046. Paz et al. (2009a) introducen un modelo de geometría sol-sensor que permite hacer las transformaciones de las reflectancias a la geometría especificada:
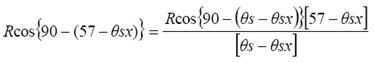 (6)
(6)
donde: R puede ser la banda del R o del IRC y es el ángulo cenital mínimo en el día de medición.
En el caso de mediciones multi-temporales de las reflectancias, las proporciones de radiación directa y difusa en la iluminación solar durante el crecimiento de la vegetación pueden variar substancialmente. En este caso, los atrayentes (reflectancias en el infinito) de la dinámica espectral permanecen invariantes (son iguales para el caso de solo radiación directa, solo radiación difusa o para radiación total - cualquier proporción de directa a difusa) bajo variaciones en las componentes de la radiación solar (Ross, 1981).
Modelo PROSPECT de Transferencia Radiativa
El modelo radiativo PROSPECT (Jacquemoud et al., 1990, 1995c y 1996), ha sido utilizado en numerosas publicaciones. Dicho modelo utiliza la relación entre la hoja y sus constituyentes foliares i:
 (7)
(7)
donde: k es el coeficiente de absorción (o absortancia α) de la hoja, ci y kj son las concentraciones (masa por unidad de área foliar) y los coeficientes de absorción específicos de los constituyentes bioquímicos i = 1, 2,....,m. N es un parámetro que depende de la estructura del mesófilo foliar (número de capas). En la relación 7 solo se ha considerado el espectro electromagnético de 500 a 2500 nm (región del verde al infrarrojo medio lejano. Para la región del azul (400-500 nm) debe agregarse un coeficiente de extinción correspondiente a una hoja albina (Jacquemoud et al., 1990).
En la versión inicial de PROSPECT (Jacquemoud et al., 1990) sólo se consideraron los coeficientes específicos del agua (kw ) y de la clorofila a+b (kab ). En versiones posteriores (Jacquemoud et al., 1996), al considerar los problemas asociados a la estimación de la proteína y celulosa+lignina (los picos de absorción están enmascarados por los del agua; Fourty et al., 1996), se incluyó a km (materia seca). La materia seca incluye a todos los componentes bioquímicos, con excepción de los lípidos y las cenizas. La Figura 4 muestra los coeficiente de absorción específicos de esta última versión (hay varias además de ésta) mencionada en el modelo PROSPECT.
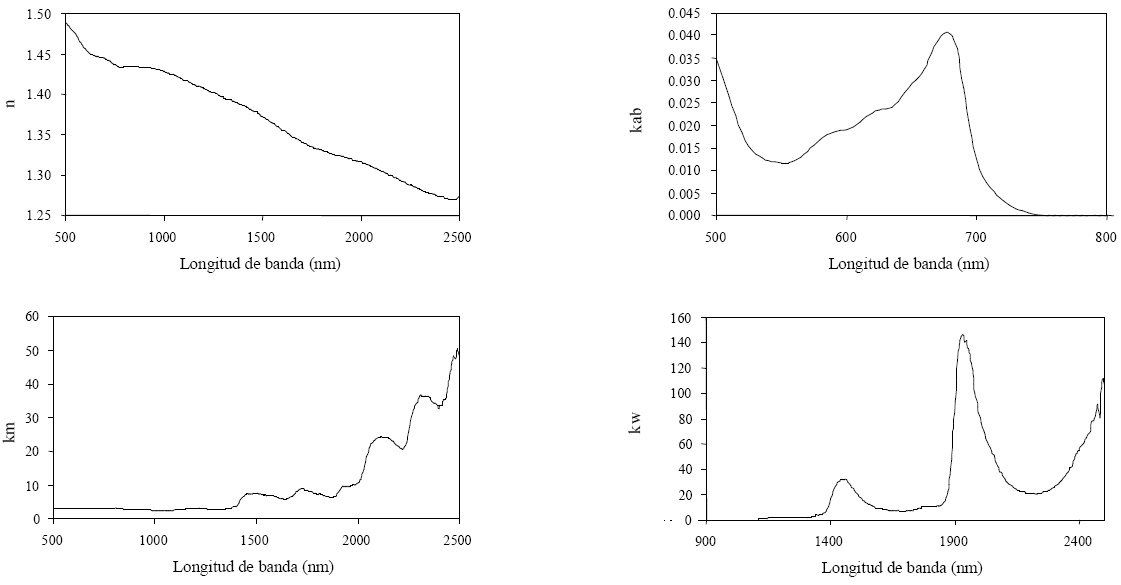
Figura 4 Funciones espectrales de las componentes del modelo PROSPECT Versión 3.01 (Fuente: http://teledetection.ipgp.jussieu.fr/opticleaf/models.htm, verificada el 20 de octubre de 2008).
Se observa, en la Figura 4, que kab tiene valores nulos más allá de 750 nm y que desde 800 nm en adelante solo hay contribuciones de km (valor constante) y kw . Adicionalmente, en la misma Figura se observa la dependencia espectral del índice de refracción n (interfase aire-células), el cual es requerido por el modelo PROSPECT al ser un modelo de placas de transferencia radiativa.
Se muestran en la Figura 5 los coeficientes de extinción específicos de la proteína (kp) y de la celulosa+lignina (kcl) de la versión del modelo PROSPECT discutida en Jacquemoud et al. (1995 c).

Figura 5 Funciones espectrales de kcl y kp del modelo PROSPECT Versión 2.01 (Fuente: http://teledetection.ipgp.jussieu.fr/opticleaf/models.htm, verificada el 20 de octubre de 2008).
La relación entre kcl y kp con km se muestra en la Figura 6, donde se observa una alta correlación en diferentes regiones espectrales.

Como se mencionó anteriormente, es importante estimar la relación C/N para caracterizar los procesos ecológicos de intercambio de masa y energía, además de los factores asociados al crecimiento de las plantas. La concentración cm (peso foliar especifico o PFE) nos permite convertir las estimación del IAF a valores de biomasa foliar aérea seca, el cual es un parámetro requerido en modelos de crecimiento de la vegetación (Goudriaan y van Laar, 1994). cm y ccarbono están altamente correlacionados (ccarbono = 0.5065 × PFE, R2 = 0.995; Fourty y Baret, 1998), lo mismo para cp y cnitrógeno (cnitrógeno = 0.1584cp , R2 = 0.963; Fourty y Baret, 1998). Una alta correlación también se presenta para ccl y ccarbono (Jacquemoud et al., 1995c y 1996). La existencia de altas correlaciones discutidas implica que los modelos radiativos no pueden discriminar adecuadamente componentes bioquímicos particulares, pero las relaciones establecidas empíricamente pueden ser usadas para estimar estos componentes. En el caso de la estimación de las concentraciones de proteínas, dado el enmascaramiento de los picos de absorción por el agua, es posible establecer relaciones entre el contenido de las clorofilas y el nitrógeno foliar y, por lo tanto, con la fotosíntesis (Field y Mooney, 1986; Evans, 1989; Ercolli et al., 1993). Las proteínas fotosintéticas representan una gran proporción del N foliar total (Field y Mooney, 1986; Evans, 1989). Lawlor (2002) revisa las bases fisiológicas de esta relación. Cerca del 75 % del nitrógeno foliar está involucrado en los procesos fotosintéticos (Evans, 1989), donde la partición del nitrógeno dentro del follaje se ha descrito en forma extensiva (Grindlay, 1977). El acoplamiento IAF-N foliar puede modelarse en forma razonable (Yin et al., 2003), permitiendo reproducir patrones temporales del N foliar congruentes con las bases fisiológicas, como los descritos por Gitelson et al. (2005).
Le Maire et al. (2004) han criticado la calibración indirecta del modelo PROSPECT, donde primero se calibra el parámetro N usando dos o tres bandas espectrales en el IRC y después se determinan los coeficientes de absorción específica (no se miden en forma directa; con excepción de kw, sino que se estiman por ajustes estadísticos). El índice de refracción también es estimado en forma independiente. Feret et al. (2008) recalibraron los coeficientes de absorción específicos usando mejorías en la determinación del índice de refracción y en el comportamiento de la superficie de las hojas, además de la utilización de métodos estadísticos conjuntos de ajustes (estimaciones simultaneas de parámetros). Un nuevo coeficiente de absorción específico propuesto es el kc de los carotenoides totales; aunque éstos presentan una alta correlación lineal con las clorofilas a+b (Gitelson et al., 2003; Feret et al., 2008). Esta mejoría del modelo PROSPECT no será considerada en este trabajo.
Parametrización Directa del Modelo PROSPECT y Esquema Generalizado de Estimaciones Bioquímicas
Recientemente, Lewis y Disney (2007) desarrollaron una aproximación directa al modelo PROSPECT (R2 > 0.9997 con relación al original de Jacquemoud et al., 1996) que simplifica fuertemente la inversión del modelo. En esta aproximación el parámetro N se dejó fijo (N = 1), ya que el PROSPECT fue parametrizado en función del albedo foliar y no de sus componentes de reflectancia y transmitancia foliares, tal como es el caso del modelo original. El parámetro N afecta solo la razón ρ/τ, que en la región del IRC implica un coeficiente de dispersión s más o menos constante.
La aproximación del modelo PROSPECT en función del albedo foliar ω puede ser puesta como (Lewis y Disney, 2007):
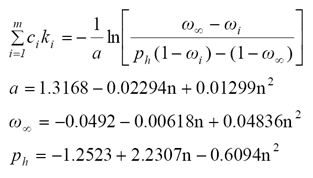 (8)
(8)
donde: ω∞ es el albedo foliar en el infinito asociado con la dispersión de la superficie de la hoja (no interactúa con los constituyentes bioquímicos foliares internos) y ph es la probabilidad de recolisión dentro de la hoja.
La relación 8, muestra que el modelo PROSPECT puede ser puesto en función sólo del índice de refracción n, el cual es función de las bandas espectrales (Figura 4). Aun usando un valor de n fijo, n = 1.39, la aproximación resulta excelente (R2 > 0.995 con relación al original).
La formulación aproximada del modelo PROSPECT tiene implícitos los esquemas empíricos de calibración usados y refleja una estrategia desagregada (ρ y τ) de modelación, la cual fue replanteada por Lewis y Disney (2007) como agregada (ω). Desde el punto de vista de la utilización de las reflectancias en el infinito, relación 5, las relaciones 8 establecen el escalamiento del follaje con las hojas y de allí a sus componentes bioquímicos. Esto puede formularse de manera directa, para el desarrollo de un modelo de transferencia radiativa en hojas acoplado a la información espectral accesible por sensores remotos (R∞ y ω) sin necesidad de recurrir a hipótesis de medios turbios o de condiciones de suelo negro (el suelo absorbe todos los fotones que chocan con él), tal como se discute en la sección siguiente.
Una forma directa de eliminar el parámetro N de la relación 7 es usar la función de remisión de medios turbios, f(R∞) = k/s y así desarrollar un esquema de modelación directo:
 (9)
(9)
donde: si es un coeficiente de dispersión específico (dos direcciones).
La relación entre el albedo foliar ω, y la absortancia foliar α = 1- ω con k/s está dada por:
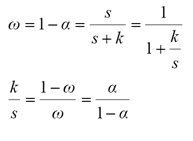 (10)
(10)
Así, la relación 9 puede ser puesta como:
 (11)
(11)
 (12)
(12)
que pueden calibrarse de forma directa con la información espectral asociada a las hojas, reduciendo las incertidumbres de los esquemas actuales de calibración usados en el PROSPECT. De las reflectancias en el infinito, relación 5 y 10, podemos establecer:
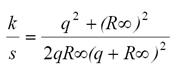 (13)
(13)
que se puede usar en la formulación propuesta en las relaciones 11 y 12.
Un planteamiento similar al uso de una relación 13 la hicieron Schmuck et al. (1993), en la 9, pero utilizaron la hipótesis limitada de (k/s) ? α, además de que solo plantearon el modelo para el caso de una distribución angular uniforme (medio turbio). En forma similar, Gitelson et al. (2005), usaron un enfoque tipo empírico (índices espectrales) bajo la misma hipótesis que Schmuck et al. (1993). De las relaciones 10, resulta claro que la aproximación entre k/s y α utilizada por estos autores implican una constante aditiva y multiplicativa adicionales que permanece aún después de los intentos de filtrarla (véase a Gitelson et al., 2005). Los autores mencionados utilizan un coeficiente de retro-dispersión s' = s/2.
Invariantes espectrales y escalamiento de las estimaciones
La teoría-p o de invariantes espectrales (Knyazikhin et al., 1998; Panferov et al., 2001; Smolander y Stenberg, 2005; Stenberg, 2007; Huang et al., 2007; Lewis y Disney, 2007; Knyazikhin et al., 2013) plantea en forma físicamente coherente (respecto de la ley de conservación de energía) que la ecuación tri-dimensional de la transferencia radiativa tiene simetrías (escalamientos) que permiten simplificar en forma fuerte su solución. Esta idea de escalamiento y simetría ha sido planteada previamente por van de Hulst (1980) y desarrollada por Mitrescu y Stephens (2004) para la función de fase anisotrópica y así generar relaciones iguales a las del grupo mencionado, pero en el contexto de modelaciones atmosféricas. La similaridad implicada en la ecuación de transferencia radiativa establece simplemente que diferentes problemas radiativos, asociados a diferentes funciones de fase, pueden ser hechos equivalentes unos con los otros usando diferentes pares del espesor óptico (parámetro estructural, fundamentalmente) y el albedo foliar simple (dependiente de la banda espectral).
La teoría-p, caso de suelo negro, plantea el escalamiento entre el follaje de la vegetación y las hojas que lo componen:
 (14)
(14)
donde: p es la probabilidad de recolisión de los fotones chocando con la vegetación; es decir, es la probabilidad media de que un fotón dispersado por una hoja del follaje de la vegetación interactúe de nuevo dentro del follaje. W es el albedo de dispersión simple, K es el coeficiente de absorción (absortancia) y S es el coeficiente de dispersión, todos a nivel del follaje de la vegetación y no de las hojas. En las relaciones 14 se ha supuesto p como constante para todos los órdenes de interacción, por lo que los valores p corresponden a valores efectivos pef (Huang et al., 2007). En el caso de la absorbancia, p alcanza su valor teórico después de solo una interacción, por lo que se puede establecer una relación de escalamiento entre el follaje y la vegetación (Knyazikhin et al., 1998):
 (15)
(15)
que puede también ponerse en función del albedo de dispersión simple W, ya que la absortancia del follaje es definida como A = 1- W. La relación 15 establece en forma clara que la información espectral asociada a cualquier banda espectral (λ), ancha o angosta, está relacionada con cualquier otra (λ0) a través de los albedos foliares (que son función de λ) y el parámetro p que es función de la estructura de la vegetación (independiente de λ). Así, la discusión de bandas "óptimas" de los sensores remotos para alguna estimación específica debe estar relacionada con los albedos foliares y sus constituyentes bioquímicos.
Los desarrollos analíticos de la teoría-p son solo válidos para el caso de suelo negro, por lo que es necesario desarrollar un esquema de conversión de las reflectancias de un suelo cualquiera a uno negro (Figura 2b.) En el caso de las reflectancias en el infinito, la condición de suelo negro se satisface, por lo que pueden usarse las relaciones 14 en forma directa para relacionar mediciones del follaje con las de las hojas. En el caso de medios ópticamente densos, el valor de p es de alrededor de 0.9 para el caso asociado a θs ? 57° (Lewis y Disney, 2007), discutido al inicio de este trabajo.
En el caso de los desarrollos de similaridad y escalamiento en el contexto de las aplicaciones atmosféricas, una función de fase altamente anisotrópica puede descomponerse en dos partes: una en la dirección contraria a la iluminación y otra en la dirección de retro-iluminación. Así, definiendo a f como la fracción (probabilidad) de la energía incidente dispersada en la dirección contraria a la iluminación, Mitrescu y Stephens (2004) establecieron:
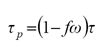 (16)
(16)
 (17)
(17)
donde: τ es el espesor óptico.
Para un medio con dispersores isotrópicos iluminados por radiación isotrópica, este se vuelve ópticamente denso (reflectancias en el infinito) cuando (van de Hulst, 1980):
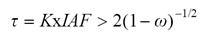 (18)
(18)
donde: K es un coeficiente de extinción.
Las relaciones 16 y 17 representan las relaciones de similaridad introducidas por van de Hulst (1980), que establecen que la solución de la ecuación de transferencia para funciones de fase compleja puede ser aproximada en términos de soluciones con funciones de fase más simple; esto es, se puede resolver la ecuación de transferencia radiativa usando valores escalados del albedo de dispersión simple y el espesor óptico al utilizar funciones de fase más simétricas. Esta representación es una propiedad de la ecuación de transferencia radiativa y no de la función de fase. Este esquema de similaridad y escalamiento ha sido usado en forma semi-empírica por Bolaños et al. (2007) para aproximar la función de distribución bi-direccional de reflectancias en la vegetación.
Conclusiones
Los desarrollos mostrados en este trabajo plantean un esquema integrado de escalamiento de las propiedades ópticas y geométricas del follaje de la vegetación, medios homogéneos y heterogéneos, con las propiedades ópticas y bioquímicas de las hojas. El esquema de conversión de medios heterogéneos a homogéneos permite acceder el albedo foliar en términos de la información espectral disponible en los sensores remotos, además de generar una condición de frontera de suelo negro y medio ópticamente denso (con reflectancias en el infinito asociadas), que pueda usarse para establecer relaciones escalantes basadas en la teoría-p o de invariantes espectrales. De esta forma, el parámetro foliar fundamental albedo puede ser estimado en términos desacoplados de las propiedades estructurales (índice de área foliar y cobertura de la vegetación). El marco teórico introducido abre un nuevo paradigma para asociar mediciones de sensores remotos, en campo o en plataformas espaciales, con estimaciones de las componentes bioquímicas foliares en forma directa, evitando los problemas asociados a los múltiples enfoques empíricos que han intentado aproximar estas estimaciones.
De acuerdo a los desarrollos mostrados, es posible relacionar directamente el albedo foliar estimado con modelos de transferencia radiativa en hojas, tales como el PROSPECT, con las estimaciones de las componentes bioquímicas foliares. Así, la estimación, sin el enmascaramiento estructural (IAF), de las concentraciones de los componentes bioquímicos de las hojas puede hacerse en forma estable y congruente con el marco teórico de la transferencia radiativa. Alternativamente, es posible replantear la construcción de un modelo de transferencia radiativa en hojas parametrizándolo en forma directa en función de los albedos o absortancias foliares, reduciendo así las incertidumbres asociadas a las calibraciones del modelo cuando se usan calibraciones basadas en reflectancias y transmitancias foliares por separado.

