











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Actualmente la salinidad en el suelo es el mayor factor limitante para la expansión de la frontera agrícola en el noroeste de México (Ashraf et al., 2008). Las perspectivas al respecto no son muy alentadoras. Se espera que el incremento de la salinidad en las tierras cultivables en el mundo resulte en una pérdida del 30% de estas tierras en los próximos 25 años y más del 50% para la segunda mitad del siglo XXI (Mahajan y Tuteja, 2005). En las zonas áridas y semiáridas, el problema se acentúa por la escasez y variabilidad de las precipitaciones, junto con las altas tasas de evapotranspiración, lo cual afecta el balance hídrico de los suelos. La baja humedad atmosférica y las altas temperaturas que inciden en las zonas áridas y semiáridas, incrementan el ascenso de la solución del suelo y con ello la deposición y concentración de sales en los horizontes superficiales, causando la acumulación de cloruros, sulfatos, Na, Mg y Ca en los primeros centímetros del suelo (Manchanda y Garg, 2008). Es así como el agua que suministran los sistemas de riego aumenta la concentración de sales año con año en el suelo. Ello origina importantes pérdidas de superficie cultivable que se reflejan directamente en la economía de los productores (Mahajan y Tuteja, 2005).
Por otra parte, el estrés salino afecta la mayoría de los procesos fisiológicos de los vegetales como el crecimiento, la fotosíntesis, la síntesis de proteínas y el metabolismo energético (Apse y Blumwald, 2002; Tester y Davenport, 2003; Parida y Das, 2005). Sin embargo, la duración y severidad del período de estrés, al igual que la especie, determinarán la magnitud de los daños. La presencia de sales en el suelo inhibe el crecimiento de las plantas, debido a su influencia sobre procesos fisiológicos, tales como fotosíntesis, conductividad estomática, ajuste osmótico, absorción de iones, entre otros (Willadino y Cámara, 2005; Munns y Tester, 2008).
Por tanto, muchas investigaciones se están dirigiendo hacia la identificación de los procesos fisiológicos y bioquímicos que se ven afectados por el NaCl, con el propósito de aumentar la tolerancia de las plantas a la salinidad (Apse y Blumwald, 2002; Tester y Davenport, 2003).
En México, aproximadamente el 60% del territorio está conformado por zonas áridas (Rivera et al., 2004), específicamente la Península de Baja California, se ha caracterizado en los últimos años, por las altas concentraciones de sales que oscilan desde los 3 hasta los 10 g L-1 de agua; problema que se agudiza con el fenómeno de intrusión salina. Los efectos de la salinidad en Baja California Sur, han disminuido la productividad de los cultivos agrícolas y por ende la rentabilidad de los mismos. Desde los 80's, el género Ocimum en Baja California Sur, se ha caracterizado por ser un cultivo que ha ofrecido a los productores agrícolas una alternativa en respuesta a la falta de variedad de cultivos agrícolas. Una solución parcial a estos problemas es la obtención de variedades de especies cultivadas tolerantes a la salinidad. Por eso la importancia de conocer la tolerancia a la salinidad de diferentes especies y variedades como la de albahaca, siendo una alternativa para los productores (González y Ramírez, 2002; Mesa, 2003). Con base en lo anterior los estudios comparativos de plantas cultivadas en condiciones de estrés, son importantes para entender y transferir en el futuro los rasgos de tolerancia de las plantas sensibles.
La albahaca es una especie importante en el campo de las plantas aromáticas debido a su uso como condimento, además de poseer aceites esenciales que emanan un agradable aroma y sabor. Tiene uso farmacéutico por sus propiedades diuréticas y estimulantes; empleada en la industria de la elaboración de perfumes (Klimánková et al., 2008). De la albahaca se obtiene un aceite esencial que contiene cineol, metilcahvicol, linalol, estragol, eugenol y timol, los cuales son de amplia aplicación en las industrias alimentaria, farmacéutica, cosmética y de perfumería (Benavides et al., 2010). Su uso como especia está ampliamente difundido en diversos países. En México, además, tiene numerosas aplicaciones en medicina tradicional.
En el presente estudio se eligió albahaca, ya que en ambientes áridos salinos de la Península de Baja California, ha figurado como un cultivo promisorio para el sector agroindustrial. Sin embargo, su producción es mermada por la salinidad, por lo que es importante evaluar variedades con características halotolerantes. El objetivo del estudio fue determinar el efecto del estrés salino en el crecimiento y desarrollo de plantas de variedades de albahaca.
Materiales y métodos
Sitio de Estudio
El experimento se realizó dentro de una estructura de malla sombra, ubicada en el Centro de Investigaciones Biológicas del Noroeste, México, localizado al norte de la ciudad de La Paz, Baja California Sur, México, que se ubica a los 24° 08' 10.03" N y 110° 25' 35.31" O, a una altitud de 7 m.
Material Genético
Se utilizaron semillas de 20 variedades de albahaca (Ocimum basilicum L.) provenientes de la empresa Seed Company (USA) las cuales son: Sweet Dani, Lemon, Sweet Genovese, Siam Queen, Red Rubin, Thai, Dark Opal, Spicy Glove, Licorice, Cinnamon, Mrs Burns, Purple Ruffles, Lettuce Leaf, Italian Large Leaf, Genovese, Dolly, Emily, Genovese Italian, Dolce Vita Blend y Napoletano, cuyo origen fue Estados Unidos de América. No existe información sobre la tolerancia o sensibilidad de las variedades a la salinidad. Con el fin de evaluar la calidad de las semillas de las variedades en estudio, previo al experimento se realizó una prueba de germinación, con la metodología propuesta por ISTA (1999).
Diseño Experimental y Tratamientos
Las semillas se sembraron en charolas de poliestireno de 200 cavidades, las cuales contenían como sustrato sogemix PMMR, el cual contiene turba de Sphagnum Canadiense (65-75% vol-1), vermiculita, cal dolomítica y calcítica, macronutrientes, micronutrientes y un agente humectante. Para mantener la humedad, se aplicaron riegos diarios con el fin de lograr una emergencia homogénea de las plántulas. El trasplante se realizó cuando las plantas presentaron una altura promedio de 15 cm en macetas con capacidad aproximada de 1 kg, mismas que contenían sustrato comercial sogemix PMMR. En cada maceta se colocó una planta. Una vez que se trasplantaron, se inició con la aplicación diaria del riego, con una frecuencia de dos veces al día un volumen de 500 cm3, con agua potable, la cual contenía una solución nutritiva cuyas sustancias utilizadas fueron, nitrato de potasio, nitrato de amonio, fosfato mono amónico, nitrato de calcio, sulfato de magnesio, sulfato ferroso, sulfato de manganeso, sulfato de zinc, sulfato de cobre, ácido bórico) preparada de acuerdo a Samperio (1997). Después de una semana del trasplante, se inició con la aplicación de los tratamientos salinos. El experimento se estableció en un diseño completamente al azar con arreglo factorial y cuatro repeticiones, considerando las veinte variedades de albahaca como factor A y a los tratamientos salinos como factor B, con tres niveles (0, 50, 100 mM de NaCl), para un total de 60 tratamientos y 240 plantas. Cada repetición estuvo representada por una maceta con una planta. Durante la segunda semana se inició con la aplicación gradual de los tratamientos salinos, con el fin de evitar un shock osmótico en las plántulas, acorde con la metodología propuesta por Murillo-Amador et al. (2007). La cantidad de solución salina aplicada en cada riego fue de 500 mL, consiguiendo que la solución aplicada drenara a través de los orificios de las macetas, con el fin de evitar la acumulación de sales en el sustrato. El pH de la solución con los tratamientos salinos y los nutrientes se ajustó a un valor de 6.5, adicionando KOH o H2 SO4.
Variables Morfométricas
A los 45 días de aplicación de los tratamientos salinos, las plantas se trasladaron al laboratorio donde se procedió a separar raíz, tallo y hojas, se midió longitud de tallo (cm), longitud de raíz (cm), biomasa (g) fresca y seca de raíz, tallo, hoja y área foliar (cm2) esta última se determinó mediante un integrador de área foliar (Li-Cor, modelo-LI-3000A, serie PAM 1701). Para determinar peso fresco y seco de biomasa, se utilizó una balanza analítica (Mettler Toledo, modelo AG204). La obtención de biomasa seca en todas las etapas fenológicas iniciales, como son los tejidos correspondientes a hojas, tallos o raíces, se obtuvó colocando el material en bolsas de papel y secado en una estufa (Shel-Lab, modelo FX-5, serie-1000203) a una temperatura de 80 °C, hasta obtener su deshidratación completa (72 h). Posteriormente se pesaron en una balanza analítica (Mettler Toledo, AG204), el resultado se expresó en gramos de materia vegetal seca.
Análisis Estadístico
Se realizaron análisis de varianza y comparaciones múltiples de medias (Tukey, P ≤ 0.05). Los análisis se realizaron con el programa estadístico Statistica v. 10.0 para Windows (StatSoft, Inc., 2011).
Resultados y discusión
Se presentaron diferencias significativas en la interacción variedades × concentración de NaCl para longitud de tallo (F =38 180 = 6.10, P 0.001); longitud de raíz (F =38 180 = 17.98, P ≤ 0.001); biomasa fresca de raíz (F =38 180= 16.64, P ≤ 0.001); biomasa seca de raíz (F =38 180= 33.36, P ≤ 0.001); biomasa fresca de tallo (F =38 180= 19.53, P ≤ 0.001); biomasa seca de tallo (F =38 180= 30.68, P ≤ 0.001); biomasa fresca de hoja (F =38 180 = 14.62, P ≤ 0.001); biomasa seca de hoja (F =38 180= 8.06, P ≤ 0.001) y área foliar (F =38 180= 3.46, P ≤ 0.001).
Todas las variables que se evaluaron en las diferentes variedades mostraron valores mayores en el control (0 mM) y disminuyeron conforme los niveles de NaCl se incrementaron (Cuadro 1, 2 y 3). Lo anterior confirma que las sales tienen un efecto negativo en el desarrollo de la plantas de albahaca. Para longitud de raíz, las variedades Sweet Dani y Dolly, mostraron los valores mayores en 0 mM, exhibiendo un grado de afectación de 23.45% y 43.53% para 50 y 100 mM, respectivamente. Mientras tanto las variedades Emily, Red Rubin y Genovese Italian e Italian Large Leaf, tuvieron los valores menores para esta variable en 0, 50 y 100 mM, con una afectación de 29.42% en 50 mM y 54.25% en 100 mM con respecto a 0 mM (Cuadro 1). Para biomasa fresca de raíz, la variedad Napoletano mostró los valores mayores en 0, 50 y 100 mM, teniendo un grado de afectación de 27.84% con 50 mM y 45.61% con 100 mM, mientras que los valores menores en 0 mM, lo presentaron las variedades Siam Queen, Red Rubin, Cinnamon, Italian Large Leaf y Emily, presentando un grado de afectación de 39.01% en 50 mM y 74.47% en 100 mM. La variedad Red Rubin mostró el valor inferior en 50 mM y Siam Queen en 100 mM (Cuadro 1). Acorde con De Araujo et al. (2006) la salinidad puede inhibir el crecimiento de las plantas debido a varios factores, incluyendo la toxicidad iónica, deficiencia en la nutrición mineral y cambios en las relaciones hídricas. Esto último es un efecto similar al producido por la sequía. El grado en el que cada uno de estos factores puede afectar el crecimiento depende de la variedad y de las condiciones ambientales, por lo antes expuesto, en el presente estudio las variedades Sweet Dani, Dolly exhibieron mayor tolerancia a la salinidad al mostrar valores mayores de longitud de raíz, longitud del tallo y área foliar, además la variedad Napoletano presentó un mayor desarrollo en la biomasa fresca y seca de raíz, tallo y hoja. La salinidad del suelo provoca una sintomatología relacionada con una inhibición irreversible del crecimiento en cuanto a menor área foliar y talla de la planta (el crecimiento es lento y no llega a ser completo), menor producción de materia seca, quemaduras en los bordes de las hojas, entre otros factores (Campos et al., 2011). Los principales iones salinos Na y Cl, pueden suprimir la absorción de nutrimentos debido a las interacciones competitivas iónicas o afectar la integridad de la membrana. Niveles altos de Na por lo general indican deficiencias de K. El estrés iónico se asocia con relaciones altas de Na/K y Na/Ca y acumulación de Na y Cl en los tejidos, lo cual es dañino para el metabolismo general de las células. El mantenimiento de K citosólico y la homeostasis de la concentración intracelular de iones es aún más crucial en estrés salino. Otros efectos negativos de la exposición a la salinidad en el sustrato, se manifiesta en las reducciones en el crecimiento y muerte de órganos, asociadas al ingreso de Na, disminución en la concentración interna de K y Ca, restricciones al ingreso de agua y la expansión celular (Hasegawa et al., 2002). Los resultados obtenidos en el estudio coinciden con los reportados por Llanes et al. (2005) al estudiar la respuesta del crecimiento de Prosopis strombulifera a soluciones monosalinas osmóticas de KCl, NaCl, Na2SO4 y K2So4, encontraron que en los tratamientos salinos y un pH ácido, el crecimiento de los hipocótilos siguieron un patrón de inhibición del crecimiento similar al de las radículas. De la misma manera, los resultados encontrados en biomasa fresca de raíz coinciden con lo determinado por Mora-Aguilar (2004) quien estudió la salinidad en plantas de Solanum tuberosum y encontró que en todos los tratamientos salinos, excepto en los de NaHCO3 y salinidad sulfático-sódica, las plantas acumularon menos biomasa, lo que se atribuye a la disminución natural del crecimiento de la raíz al final del ciclo biológico de la planta, pero también al efecto salino. De manera general se puede considerar que el efecto en el desarrollo radicular de las plantas de albahaca bajo estrés salino se debe a una restricción en el crecimiento celular, debido al potencial bajo de agua del medio externo, a la interferencia de los iones salinos con la nutrición de las plantas o a la toxicidad de iones acumulados que conducen a la muerte celular. Asimismo, los resultados del estudio coinciden con los obtenidos por Al-Karaki (2000) quien evaluó el efecto del NaCl sobre las raíces de genotipos de L. esculentum, cultivares Sera, 898 y Rohana, determinando que el incremento de la concentración de la salinidad afecta adversamente el crecimiento de las raíces, con lo que trae como consecuencia menor biomasa fresca.
Cuadro 1 Respuesta de la interacción de los factores variedad × salinidad en la longitud de tallo, altura de planta y biomasa fresca de raíz de variedades de albahaca sometidas a estrés salino.
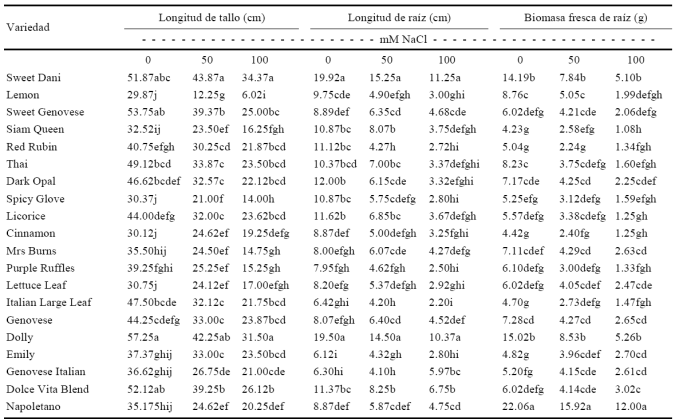
*Medias con letras distintas en una misma columna difieren estadísticamente (Tukey P ≤ 0.05).
Cuadro 2 Respuesta de la interacción de los factores variedad × salinidad en la biomasa seca de raíz y biomasa fresca y seca de tallo de variedades de albahaca sometidas a estrés salino.
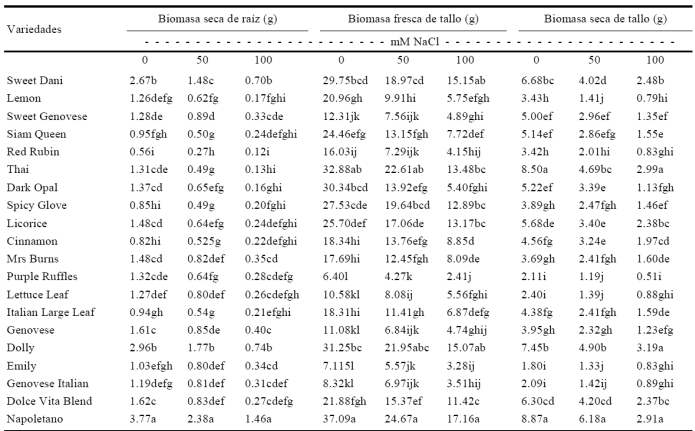
*Medias con letras distintas en una misma columna difieren estadísticamente (Tukey P ≤ 0.05).
Cuadro 3 Respuesta de la interacción de los factores variedad × salinidad en la biomasa fresca y seca de hoja de variedades de albahaca sometidas a estrés salino.
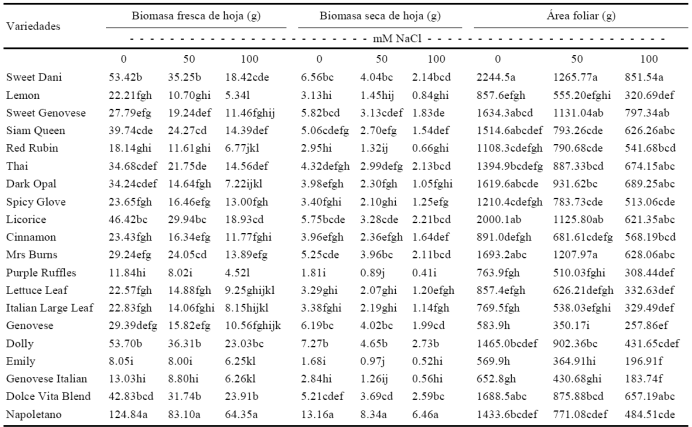
*Medias con letras distintas en una misma columna difieren estadísticamente (Tukey P ≤ 0.05).
La variedad Sweet Dany presentó mayor longitud de tallo con un menor grado de afectación de un 15.42% para 50 mM y un 33.73% para 100 mM de NaCl. La variedad Lemon mostro un mayor grado de afectación con un 58.98% en 50 mM y un 79.84% en 100 mM respectivamente (Cuadro 1). Estos resultados concuerdan con lo establecido por Wahome (2003) quien menciona que las plantas son afectadas ante el estrés salino, al disminuir su tasa de crecimiento, formar hojas más pequeñas y menor cantidad de ellas y disminuir su altura, con estas mediciones se podría seleccionar materiales tolerantes y sensibles a la salinidad en estas etapas, puede efectuarse exitosamente y proporcionar una técnica efectiva para realizar tamizados, selecciones y clasificaciones rápidas de material genético para tolerancia a este estrés. La tolerancia de las plantas a la salinidad puede ser determinada por su altura, lo cual se le atribuye a que la salinidad reduce el acceso de agua para la planta y disminuye el crecimiento de la misma. El exceso de las sales causa daños celulares a través del proceso de transpiración en las hojas y por toda la planta, por lo que el crecimiento se inhibe (Munns et al., 2006). Por otra parte, Chilo et al. (2009) estudiando el efecto de la temperatura y la salinidad en la germinación y el crecimiento de partes aéreas de dos variedades de Chenopodium quinoa indicaron que el crecimiento disminuyó con una baja temperatura y alta salinidad.
La variable biomasa seca de raíz fue mayor en la variedad Napoletano mostrando una afectación de un 36.88% en 50 mM y 61.28% para 100 mM y menor en Red Rubin, exhibiendo una afectación de 51.79% en 50 mM y 78.58% para 100 mM ambas en 0, 50 y 100 mM, respectivamente (Cuadro 2). Estos resultados coinciden con lo señalado por Abrisqueta et al. (1991) quienes encontraron que conforme se incrementaron los niveles de salinidad a partir de 50 hasta 100 mM de NaCl, se redujo la biomasa seca de raíz. Esto puede atribuirse a la restricción en el crecimiento celular, debido al potencial bajo de agua del medio externo y a la interferencia de los iones salinos con la nutrición de las plantas o a la toxicidad de iones acumulados que conducen a la muerte celular (Cuartero y Fernández-Muñoz, 1999). La acumulación de biomasa seca se usa ampliamente como una medida de crecimiento de la planta, porque refleja un balance entre la producción total de foto-asimilados y la respiración. Según Shani y Ben-Gal (2005) su reducción como consecuencia de la salinidad está asociada con disminuciones equivalentes en la absorción de agua debido a un efecto osmótico, ya que la biomasa fresca y seca de la parte aérea disminuyó en todas las variedades conforme se incrementaron los niveles a NaCl de 50 y 100 mM. La acumulación menor de biomasa fresca y seca observada en los tratamientos salinos se atribuye al efecto osmótico que resulta de la concentración elevada de sales disueltas que disminuyen el potencial osmótico de la solución y consecuentemente la disponibilidad de agua para la planta (Willadino y Camara, 2004; Tadeo y Gómez-Cadena, 2008). El déficit hídrico inducido por la salinidad restringe el crecimiento y ocasiona una menor tasa de división y elongación celular, así como disminución de la asimilación de CO2 (Pio et al., 2001). Los resultados del estudio son comparables con los señalados por Pio et al. (2001), quienes encontraron que la materia seca de cinco porta-injertos de vid disminuyó proporcionalmente con los niveles de salinidad. Por su parte, Cavagnaro et al. (2006) mencionan que las respuestas diferenciales al someter a salinidad cultivares de vid europeos como Malbec, Cabernet Sauvignon y Chardonnay junto a cultivares criollos argentinos, presentaron mayores contenidos de biomasa fresca y seca.
Otro estudio atribuye al efecto osmótico y al impacto de la salinidad en el crecimiento, al efecto del estrés osmótico en la zona radical y esto trae consigo la reducción del peso, que coincide con la reducción de área foliar y biomasa de la planta (Urrestarazu, 2004). La respuesta en características morfológicas de raíz de albahaca, muestra que el estrés salino conduce a cambios en crecimiento y en consecuencia en morfología, así como en la fisiología de este órgano, lo que turnará en cambios en la absorción de agua e iones en la producción de señales (hormonas) que comunican información a la parte aérea. En términos fisiológicos y de metabolismo, la raíz como órgano de adsorción, tiene importancia en la respuesta a corto y largo plazo al estrés salino. Aquí se sintetiza ácido abscísico (ABA) una de las señales de estrés, capaz de producir cambios fisiológicos (conductividad hidráulica) y a distancia (cierre estomático) (Hartung et al., 2002) por lo que las características anatómicas y morfológicas de la raíz tienen efecto en la capacidad de adaptación a la salinidad (Maggio et al., 2001). La relación salinidad y crecimiento de raíz resulta compleja, ya que factores como la composición iónica de las sales, la relación Na:Ca, afectan la respuesta del crecimiento de la raíz e incrementan su biomasa.
La biomasa fresca de tallo fue mayor en la variedad Napoletano respecto al resto de las variedades, siendo más tolerante a la salinidad para esta variable al mostrar menor afectación en 50 mM (33.49%) y 100 mM (53.74%), mientras que la variedad más sensible a la salinidad fue Purple Ruffles exhibiendo un mayor grado de afectación de 35.29% en 50 mM y 62.35% en 100 mM (Cuadro 2). La biomasa seca de tallo fue mayor en Thai y Napoletano en 0 mM y menor en Purple Ruffles, Lettuce Leaf, Emily y Genovese Italian. La variedad Napoletano mostró mayor biomasa seca de tallo en 50 mM de NaCl exhibiendo una afectación de un 44.83%, mientras que las variedades Purple Ruffles, Lemon, Lettuce Leaf y Emily mostraron valores menores, presentando la variedad Purple Ruffles una afectación de 43.61%. Por su parte, Thai, Napoletano y Dolly mostraron mayor biomasa seca de tallo en 100 mM, mostrando la variedad Thai una afectación de 64.83% y Purple Ruffles el valor menor, exhibiendo una afectación de 75.83% (Cuadro 2). Los resultados son acordes con lo planteado por Villafañe (1997) quien menciona que las sales acumuladas en el suelo afectan el crecimiento y desarrollo de la planta, al restringir la absorción de agua con la disminución del potencial osmótico; además, las plantas del tratamiento salino mostraron modificaciones morfológicas en las hojas y mayor acumulación de los iones de sodio y cloruro, lo que sugiere ausencia de mecanismos de exclusión de estos iones, sin embargo, una de las perspectivas de introducción de la albahaca está dentro de los indicadores más significativos para la producción agrícola qué es la producción de biomasa fresca y seca de parte aérea ya que representa la parte de la planta que se comercializa. En este sentido, se observó que para todas las variables morfométricas medidas en esta etapa, se encontraron diferencias significativas entre las variedades, la salinidad y la interacción de variedades × salinidad, lo cual podría representar un cultivo alternativo para condiciones de salinidad. Por su parte, Farhatulah y Raziuddin (2002) encontraron que el peso fresco y seco de tallo disminuyó con el incremento de los niveles de NaCl. La respuesta al NaCl de variables morfométricas de albahaca, muestran similitud con otros estudios, donde se asevera que la salinidad disminuye el crecimiento y desarrollo de las plantas, especialmente glicófitas, mediante la inducción de mal funcionamiento de procesos fisiológicos (Shannon et al., 1994). Además, la exposición de las plantas al NaCl, afecta el transporte de iones y agua (Läuchli y Epstein, 1990) así como el crecimiento y desarrollo vegetativo (Ebert et al., 1999).
La variedad Napoletano mostró mayor biomasa fresca de hoja en 0, 50 y 100 mM, presentando una afectación de 33.44% en 50 mM y 48.46% en 100 mM, mientras que Purple Ruffles y Emily exhibieron los valores menores en 0, 50 y 100 mM de NaCl, con afectación de 93.58% en 50 mM para la variedad Purple Ruffles. Las variedades Lemon y Purple Ruffles mostraron los valores inferiores en 100 mM, siendo mayormente afectada en 100 mM la variedad Purple Ruffles con 96.38% respecto a 0 mM (Cuadro 3). La variedad Napoletano presentó el valor mayor de biomasa seca de hoja en 0, 50 y 100 mM, exhibiendo una afectación de 36.63% para 50 mM y 50.92% para 100 mM, mientras que el valor menor lo presentaron las variedades Purple Ruffles y Emily en 50 mM de NaCl, mostrando la variedad Purple Ruffles una afectación de 50.83% en 50 mM y 77.35% en 100 mM (Cuadro 3). La acumulación de osmorreguladores de iones en las hojas es una respuesta adaptativa de las plantas a altas concentraciones de sal en el medio. A través de este mecanismo, las plantas pueden hacer la regulación osmótica (Casierra-Posada y Rodríguez, 2006). El fenómeno de la osmorregulación se ha observado en hojas (Westgate y Boyer, 1985) y en muchas de las plantas evaluadas en condiciones de salinidad se ha encontrado una relación directamente proporcional entre el peso específico de las hojas y la concentración de sales en el sustrato (Casierra-Posada y García, 2005; Casierra-Posada y Hernández, 2006; Casierra-Posada y Rodríguez, 2006); por el contrario, en algunos casos se ha encontrado que el peso específico de las hojas se reduce a niveles crecientes de NaCl en el sustrato (Syeed y Khan, 2004; Ewe y Sternberg, 2005) lo que puede indicar que la salinidad induce la formación de hojas delgadas, además, es posible que en condiciones de salinidad se acumule menor cantidad de asimilados en las hojas, como consecuencia de una tasa reducida de fotosíntesis en respuesta a la acumulación de Na y Cl en las hojas, pues se ha encontrado que el aumento en niveles de estos iones se debe a la salida pasiva de iones en las membranas para lograr el ajuste osmótico (Syeed y Khan, 2004). Al igual que en el ensayo, la respuesta de las plantas a la salinidad en lo relacionado con el peso específico de las hojas tiene un fuerte componente varietal (Syeed y Khan, 2004; Ewe y Sternberg, 2005; Casierra-Posada y García, 2005). Los resultados del estudio también coinciden con lo reportado por Mohammad et al. (1998) quienes determinaron que la respuesta inmediata al estrés salino en tomate, es la reducción de la tasa de expansión de la superficie foliar, lo que reduce la biomasa fresca y seca de la hoja. En otras especies como algodón, se indica que el incremento en los niveles de NaCl resulta en una disminución significativa en la parte aérea y el crecimiento de la biomasa foliar (Meloni et al., 2001). Por otra parte, Farhatulah y Raziuddin (2002) encontraron que el peso fresco disminuyó con los niveles de NaCl. Se ha demostrado que la disminución en el peso fresco y seco de parte aérea parece no depender de la reducción en el número de hojas, lo cual ocurre solamente en conductividades eléctricas por arriba de 6 dS m-1 (Cruz y Cuartero, 1990).
La variedad Sweet Dani mostró los valores mayores de área foliar en 0 y 50 mM, exhibiendo una afectación de 43.61% en 50 mM, mientras que los valores menores lo presentaron las variedad Emily con una afectación de 35.97% en 50 mM y Genovese en 0 mM. Las variedades Sweet Dani y Sweet Genovese exhibieron la mayor área foliar en 100 mM, presentando Sweet Dani una afectación de 62.07% en 100 mM y la variedad Genovese Italian y Emily mostraron valores inferiores, destacándose la variedad Emily con una afectación de 65.45% en 100 mM. (Cuadro 3). La salinidad reduce el crecimiento de la parte aérea suprimiendo la iniciación y la expansión de las hojas así como el crecimiento de los entrenudos y acelerando la abscisión de las hojas. Se ha planteado que el área foliar es una característica suficiente para conocer la tolerancia a la salinidad y que la morfología es uno de los principales factores que influyen en el crecimiento de las plantas (Munns, 2002). Los cambios en el área foliar influyen en la capacidad para aceptar los nutrientes, agua y luz, es importante atender la respuesta morfológica cuando crecen en estrés salino, para conocer el grado de tolerancia a ese estrés. Estos resultados coinciden con los planteados por Pardossi et al. (1998) en cuanto a la reducción del crecimiento de las plantas al ser cultivadas en condiciones de salinidad, respuesta que pudiera explicarse, según Taiz (1984), por una disminución en la utilización del carbono para la síntesis de la pared celular. En términos generales, la reducción en la síntesis de la pared celular durante el estrés, es debido a las afectaciones que se presentan en la longitud de radícula y consecuentemente esto trae consigo una reducción de la biomasa fresca y seca de radícula, tallo y área foliar.
La disminución del crecimiento se ha asociado con el contenido de Cl en las hojas (Kozlowski, 1997). En Psidium guajava se ha reportado reducción del área foliar con estrés salino por NaCl (Távora et al., 2001), mayor sensibilidad en el desarrollo de los primordios foliares que en la expansión foliar y puede afectar la expansión de las hojas mediante la reducción de la presión de turgencia y de la extensibilidad de la pared celular. Resultados similares en cuanto a la reducción del área foliar en condiciones de salinidad fueron reportados también en plantas de Fragaria sp. (Casierra-Posada y García, 2005), Lycopersicon esculentum (Romero-Aranda et al., 2001) y Psidium guajava (Távora et al., 2001). De igual manera, se ha encontrado que el área foliar total en Sorghum bicolor se redujo notablemente con la salinidad (Netondo et al., 2004). La disminución del área foliar se ha atribuido a la senectud y a la muerte temprana de las hojas, a la tasa de crecimiento reducida y a la aparición retrasada de hojas (Bernstein et al., 1993). Además, las partes superiores dobladas de la hoja, la necrosis rápida de los extremos de la lámina, o ambos son índices de desarrollo anormal de la hoja que contribuye a la disminución del área foliar. En términos generales, los resultados del presente estudio muestran similitud con otros estudios publicados, donde se confirma que la salinidad causa numerosos problemas para el crecimiento y desarrollo de las plantas. Esto se debe a la inducción de un mal funcionamiento de procesos fisiológicos (Shannon et al., 1994), además de que la exposición de las plantas al NaCl, afecta también el transporte de iones y agua en las plantas (Läuchli y Epstein, 1990). La salinidad afecta a las plantas cultivadas en la fase de germinación, emergencia y desarrollo vegetativo, por tanto, causa reducción en las variables del crecimiento (Ebert et al., 1999).
Conclusiones
En la etapa de crecimiento vegetativo inicial se observó una respuesta diferencial de las variables morfométricas entre variedades en condiciones de estrés salino, destacando las variedades Sweet Dani y Dolly con una mayor longitud de tallo y raíz, mientras que Napoletano mostró mayor biomasa fresca y seca de raíz, tallo y hoja; y la variedad Sweet Dani mostró mayor área foliar, dichas variedades podrían representar un potencial para ser introducidas en condiciones de salinidad.
La variedad Napoletano fue la menos afectada en la mayoría de las variables medidas, ya que presentó valores mayores para biomasa fresca y seca de raíz, tallo y hoja en los diferentes niveles de salinidad, considerándose esta variedad como una de las más promisorias para su cultivo en condiciones de suelo y agua con problemas de salinidad, ya que las variables más significativas para la producción agrícola son la producción de biomasa fresca, que es la parte de la planta que se comercializa. La variedad Sweet Dani también se considera promisoria para su cultivo en condiciones de suelo y agua con problemas de salinidad, dado que mostró valores mayores respecto al resto de variedades para las variables longitud de tallo y de raíz y área foliar.
Se determinó que existe una respuesta diferencial entre las variedades de albahaca al someterlas a diferentes niveles de salinidad en la etapa de crecimiento vegetativo inicial, por lo que no se presentaron patrones claros que permitan la agrupación de las diferentes variedades dependiendo de su respuesta.

