











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En México se tuvo una modesta generación de patentes entre 1994 y 2008 con un promedio anual de menos de 500 solicitudes; mientras que en el mismo período, otros países alcanzaron cientos de miles (WIPO, 2009). La principal razón de lo anterior, fue un modesto presupuesto asignado a la ciencia y tecnología ~ 0.4% del PIB (WIPO, 2009). Sin embargo, en México, se podrían desarrollar tecnologías de bajo costo, utilidad práctica y fácil adopción. Particularmente, para el control o manejo de organismos perjudiciales para el hombre, que aún hoy limitan la producción y conservación de productos agropecuarios e impactan la salud humana y animal. Al mismo tiempo, es necesario reutilizar residuos animales y vegetales, descontaminar suelos y aguas y generar ambientes estériles. Para todo ello, los ácidos grasos volátiles de hasta seis carbonos (AGVs) son compuestos potencialmente útiles.
Los AGVs se encuentran en una amplia gama de ambientes, lo que puede permitir su uso. Algunos de ellos son: el suelo (Fujii et al., 2010); agua contaminada con efluentes de fábricas de explosivos (Singh et al., 2009); las nubes, donde se genera ácido fórmico en concentración de nanomoles (Stavrakou et al., 2012); y exudados de raíces de plantas (Kai et al., 2011) entre otros. El ácido acético (AA) presente en el vinagre se ha utilizado por siglos para conservar alimentos (Escabeche, 2013); asi mismo, el AA presente en la vagina de la mujer se asoció con un incremento en la absorción de fármacos (Lee et al., 2011). Algunas tecnologías a desarrollarse con los AGVs podrían llegar a ser más económicas, de uso práctico y sustentable, pero por razones de diferente índole, no se han promovido ni abordado.
Los objetivos de este trabajo fueron: i) mostrar los principios químicos que ayuden a entender el comportamiento de los AGVs, y ii) discutir usos, tecnologías e innovaciones potenciales a partir de AGVs.
Relación entre el pH Y pKa de los AGVs
Las dos formas en que se encuentran los AGVs en solución son disociada y no disociada. El pKa propio de cada uno de estos ácidos, indica su constante de disociación en donde ambas formas disociada -no disociada se encuentran en 50%. Por ejemplo, el pKa del ácido acético es de 4.75 que se puede expresar como -log10 (4.75) o bien como1/10(4.75) cuyo resultado es 1.75 × 10-5 mol L-1. Conforme el pH en una solución disminuye por debajo del pKa de cada ácido, la forma no disociada de los ácidos aumenta. Para estimar la concentración molar o mili molar de los AGV en algunas de sus dos formas, se necesitan los valores de su pKa, el pH de la solución en donde están contenidos y la concentración total de los ácidos añadidos o generados en esa solución.
Los pKa de los AGV acético (AA), isobutírico (AiB o AB), n-butírico (AnB), capróico (AC), fórmico (AF), propiónico (AP), isovalérico (AiV) y n-valérico (AnV) son 4.75, 4.84, 4.81, 4.83, 3.75, 4.87, 4.77 y 4.82, respectivamente (Tenuta et al., 2002). Entonces, el cálculo de los ácidos en su forma no disociada se describe por la siguiente fórmula:
En donde mmol AGV d y AGV nd indican milimoles de ácidos en forma disociada y no disociada, respectivamente (total de ácidos en solución); y pKa y pH son la constante de disociación del ácido y pH de la solución que contiene el ácido respectivamente. El cálculo automático de AGV nd se puede hacer utilizando la hoja de cálculo de Excel, con la siguiente fórmula: = ((celda con el valor total del ácido graso volátil) / (10^-pKa del ácido graso volátil/10^-pH de la solución donde está contenido el ácido+1)). La fórmula se puede utilizar para calcular los mmol L-1 nd de cada AGV contenidos en una mezcla, solo se requiere el pH de la solución y la concentración de cada ácido.
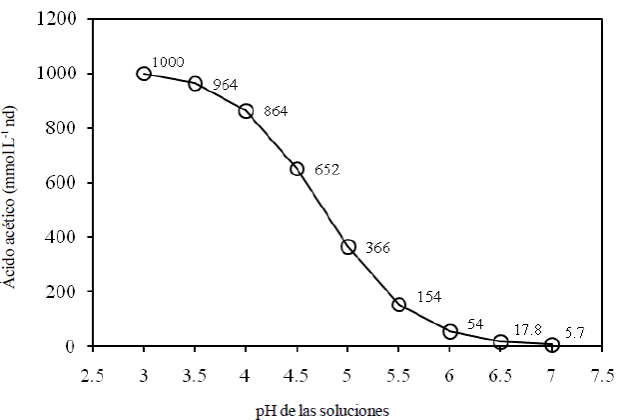
Si bien, los AGVs son potencialmente tóxicos en sus dos formas, la toxicidad es mucho mayor conforme el pH toma valores de 3 a 4, donde los AGVs se encontrarán de 85 a 99% en forma no disociada. De tal manera que, para tener una misma cantidad de un AGV en forma no disociada se requiere menos de 175 veces a pH 3 que a pH 7, como se muestra en la Figura 1. Por tanto, la toxicidad de los AGV está dada principalmente por la forma no disociada (Tenuta et al., 2002).
Efectos usuales de los AGV
Los AGVs pueden tener efecto fungicida- fungistático, bactericida-bacteriostático, nematicida, herbicida, fumigante o ser tóxicos para plantas e inducir muerte por apoptosis en células de diferentes microorganismos e incluso en células cancerosas (Hu et al., 2011). También pueden ser metabolizados por múltiples organismos o pueden ser precursores para formar metano u otros biocombustibles (Jin e Heiji, 2009). Los efectos de los AGV sobre los organismos dependen y varían de acuerdo al ácido, a su concentración, al pH del medio en el que se encuentren, sí están en forma líquida o como gas, a la temperatura, la susceptibilidad del organismo objetivo, la adaptación o resistencia adquirida por los microorganismos y a la interferencia con otras sustancias.
Efectos sobre las plantas. Las plantas y plántulas de los cultivos de arroz, trigo, cebada y otros cereales son afectadas en su crecimiento o establecimiento en presencia de AA, AB y AP a concentraciones de menos de 2 a más de 15 mmol L-1 (Marini-Kopp et al., 2007; Shan et al., 2008; Pang et al., 2007). Las plantas acuáticas que constituyen malezas Hydrilla verticillata, Potamogeton nodosus y Spartina alterniflora, son restringidas en su proliferación por AA de 38 a 250 mmol L-1 (Anderson, 2007). Los AA y AF permiten la entrada y acumulación de cadmio (Cd) en las raíces de las plantas, sin afectar su crecimiento en algunas de ellas (Sun et al., 2011). Los bosques boreales y tropicales inducen la formación del 90% de AF en la atmósfera (Stavrakou et al., 2012).
Como intermediarios en la producción de energía. Algunos AGVs son los principales insumos para las bacterias metanógenas aunque el AF es tóxico para estas bacterias, los AP y AB son comúnmente convertidos a AA, el cual es utilizado por las bacterias para formar metano. Los estiércoles fermentados son una fuente abundante de AA, que al utilizarse por las bacterias forman metano (Wang y Zhang, 2009). Tanto el pH del medio como las formas disociada -no disociada determinan la formación de metano. De tal forma que, conforme se incrementa la concentración de AGVs, el pH disminuye, y la forma no disociada de los ácidos aumenta e inhibe la actividad de las bacterias metanogénicas en el rumen (Zhou et al., 2011).
Acción contra los organismos fitopatógenos en el suelo. Bacterias, hongos y nemátodos pueden morir por aplicación de AGVs. En el caso del nemátodo Meloidogyne incognita murió con 5.7 y 1.5 mmol L-1 nd de AA y AB, respectivamente (Katase et al., 2009). Entretanto, AGVs contenidos en excretas líquidas de cerdo fueron efectivas para el control de los nemátodos Pratylenchus spp. (Mahran et al., 2008b). El hongo Verticillium dahliae fue muerto por AA, AB, AF, AP a concentraciones de 26.2, 29.0, 3.6 y 27.0 mmol L-1, respectivamente (Tenuta et al., 2002). También murió el hongo Rhizoctonia solani al estar en contacto con AA ~ 21 mmol L-1 nd obtenido del líquido del césped procesado con calor (Ushiwata et al., 2009). Una emulsión de pescado cuyo contenido de AGV fue de ~ 200 mmol L-1 fue letal para V. dahliae y Pythium ultimum cuando se agregó entre 1 y 4% en arena (Abbasi et al. 2009). Entre 17 y 33 mmol L-1 nd de AA y n-butírico (AnB) mataron clamidosporas de especies de Fusarium oxysporum y la bacteria Ralstonia solanacearum (Momma et al., 2006). El AA también se ha utilizado en forma volátil para controlar en el suelo especies de los géneros de hongos Fusarium, Rhizoctonia y Sclerotium (Abd-El-Kareema, 2009). Phymatotrichopsis omnivora es un hongo que ataca a más de 2000 especies de plantas, tanto su micelio como sus esclerocios no fueron viables con AA, AB, AF y AP a 38 μg ml-1 (ácido - aire) o 50 mmol L-1 en solución, respectivamente (Samaniego-Gaxiola y Balagurusamy, 2013).
Muerte de bacterias y hongos patógenos de humanos. De acuerdo a Yossa et al., (2011) el AA a 333 mmol L-1 disminuyó 6 órdenes de magnitud (1 × 106) la población de Salmonella typhimurium en el suelo. Según Ryssel et al. (2009 y 2011), el AA es un antimicrobiano usado al 3% elimina a Pseudomonas aeruginosa y una gama amplia de bacterias patógenas de humanos, con la posibilidad de aplicarse en tratamientos a personas quemadas para evitar infecciones. Candida albicans levadura patógena de humanos, muere en minutos por concentración de AA de 120 a 300 mmol L-1 nd (Ramsdale et al., 2008). En la bacteria Listeria innocua, disminuyó su tasa de crecimiento en más del 80 % con 5.1 mmol L-1 nd de AA (George et al., 2008).
Conservadores y desinfectantes de alimentos, desperdicios y productos agrícolas. En fase volátil los AGVs se han utilizado o están asociados con la conservación del contenido de los silos, (Nkosi et al., 2011). Especies de Aspergillus que producen micotoxinas y deterioran pistaches almacenados, fueron restringidas cuando se fumigó el fruto una hora con AA al 60% (Nawar, 2008). Como desinfectante de carne se ha utilizado el AA a 166.5 mmol L-1 (Loretz et al., 2011). Frutos en poscosecha como cítricos, tomate, cerezas fueron conservados con AA y AP, al impedir el desarrollo de varios de los hongos que los deterioran (Hassenberg et al., 2010; Sholberg, 2009; Sholberg y Randall, 2007). También se reportó que los AA y AP fueron los únicos en impedir el crecimiento de 65 especies de levaduras obtenidas de alimentos que contienen azúcar (Senses- Ergul et al., 2007). El AA a 83.3 mmol L-1 fue un antiviral efectivo para el virus de Newcastle (Reddy et al., 2010); este mismo ácido combinado con ácido peracético y peróxido de hidrógeno logró eliminar por completo bacterias y hongos patógenos de humanos y animales en instalaciones avícolas (Gehan, 2009). El ácido perfórmico, eliminó casi por completo bacterias patógenas de humanos contenidas en aguas residuales (Gehr et al., 2009). También se ha consignado la disminución de Escherichia coli en puré de pepino al adicionarle hasta 500 mmol kg-1 de AA (Lee et al., 2010). Recientemente, se logró eliminar por completo bacterias coliformes del calostro de vacas al bajar el pH a 4 adicionando AF (Collings et al., 2011). Leche de cabra adicionada con 50 mmol L-1 nd de AA redujo la cantidad de bacterias coliformes (Samaniego-Gaxiola et al., 2011).
Mecanismos de acción de los AGVs
El AA propicia que Candida albicans y Saccharomyces cerevisiae generan enzimas que destruyen su ADN, se fragmente su célula y finalmente mueran (muerte por apoptosis) (Ramsdale et al., 2008). La enzima Homoserina O-Succinil transferasa es inhibida por el AA, lo que conlleva a una carencia de síntesis de metionina y la subsecuente muerte de Escherichia coli (Giannattasio et al., 2012). Otra forma en la que el AA afecta a Clostridium thermoaceticum y S. cerevisiae es el cambio de su pH interno y su potencial de membrana lo que induce su muerte (Mills et al., 2009).
Formación e importancia de los AGVs en suelo
El proceso que explica lo que ocurre durante la formación de AGVs en el suelo, fue descrito en dos trabajos distintos. En uno, se explican los cambios del suelo inundado cultivado con arroz (Sahrawat, 2008), y en el otro, los cambios del suelo después de adicionar paja de trigo, saturar con agua y cubrirlo con plástico, esto último conocido como desinfección biológica (DBS) o anaerobia del suelo (DAS) (Momma, 2008). Durante la DAS, el oxígeno es consumido al empezar a degradarse la materia orgánica, el pH disminuye a la par que los AGVs aparecen y el potencial óxido- reducción Eh (mv) alcanza valores negativos (-100 a - 400 mV), lo que implica una condición de la solución del suelo altamente reducida y anaerobia. Quienes primero obtuvieron evidencias del efecto de los AGVs sobre un hongo fitopatógeno Fusarium oxysporum f. sp. raphani fueron Okazaki y Nose (1986).
Por tanto, en el suelo, los AGVs parecen estar involucrados directa o indirectamente en la DAS. Directamente los AGVs pueden ser tóxicos hacia organismos fitopatógenos en el suelo; indirectamente estos ácidos pueden descomponerse y originar ácido nitroso o amoniaco a pH ácido o alcalino del suelo, respectivamente. Tanto el ácido nitroso como el amoniaco son igualmente tóxicos para los organismos fitopatógenos (Conn et al., 2005; Katase et al., 2009). O bien, los AGVs en el suelo pueden provenir de compuestos como el alcohol que en contacto con el aire formará AA (Momma et al., 2010).
Producción de AGV
En los campos agrícolas, silos y otros sitios podría llegar a requerirse grandes cantidades de AGVs, los cuales pueden generarse descomponiendo residuos de plantas, basura o desperdicios de alimentos, frutas y verduras.
En contraste, el uso de AGVs en cantidades moderadas podría ser suministrado en grado industrial, el costo de AA es barato, alrededor de $20 por litro (Quiminet, 2013).
Algunos trabajos sugieren que AGVs podrían generarse en el suelo al descomponerse 3.8 y 4 Mg ha-1 de residuos de ballico (Lolium) y brócoli, respectivamente (Blok et al., 2000); o bien, degradando 30 Mg ha-1 de residuo de pasto (Messiha et al., 2007). En la solución del suelo, medios líquidos o dentro de los ensilados pueden producirse y obtenerse grandes cantidades de AGVs. En el Cuadro 1 se exponen algunos ejemplos al respecto.
Cuadro 1 Ambiente, condiciones y microorganismos involucrados en la producción de AGVs.
| Medio | Condiciones | Microorganismos | Ácidos-cantidad | Referencia |
|---|---|---|---|---|
| Suelo | Paja de trigo 39 Mg ha-1 descompuesta en suelo húmedo cubierto con plástico a pH de 5 a 6 | Bacterias nativas del suelo | AA ~ 32 y AB ~ 25 mmol kg-1 (ácidos-suelo) | (Momma et al., 2006) |
| Suelo | Paja de trigo o arroz con 20 Mg ha-1, suelo húmedo cubierto con plástico | Bacterias nativas del suelo | AA 32 y AB 25 mmol L-1 extracto acuoso del suelo | Kim et al., 2007 |
| Líquido | Fermentación de desperdicios de comida, pH final < 4. | Lactobacillusplantarum | AA 116 y AP 135 mmol L-1 | (Ye, et al., 2008) |
| Líquido | Hidrolizado ácido de ramas < 5 cm y hoja de árbol de olivo | Pichia stipitis | AA 566 y AF 273 mmol L-1 | (Díaz et al., 2009) |
| Líquido | Residuos de cereales y frutas, sujetos a temperatura y presión elevadas | Ningún organismo | AA hasta 20% y AF hasta 80% | (Jin e Heiji, 2009) |
| Líquido (bacterias) aisladas de Sedum alfredii | Medio líquido con 80 mg L-1 de Cd (CdCO3 +(PbCO3)2·Pb(OH)2) o Pb (CdCO3 +(PbCO3)2·Pb(OH)2) | Burkholderia cepacia | AF 110 mmol L-1 | (Li et al., 2010) |
| Medio líquido con 80 mg L-1 de (PbCO3)2·Pb(OH)2 | Microbacterium seperdae + Enterobacter cancerogenes | AF 140 mmol L-1 | ||
| Líquido | Vinaza del tequila | No intervienen organismos | AGV 2.5-3.4 g L-1 | (Méndez- Acosta et al., 2010) |
| Líquido | Excretas líquidas de cerdo | No intervienen organismos | AGV >250 mmol L-1 | (Mahran et al., 2008a) |
| Líquido | Encontrado en efluente de fábrica de explosivos (RDX y HMX) | No intervienen organismos | AGV >250 mmol L-1 | (Singh et al., 2009) |
| En la materia seca | Ensilado de maíz | Lactobacillus buchneri | AA 659 mmol Kg-1 | (Nkosi et al., 2011) |
Usos, Limitantes y Perspectivas de los AGVs
El uso de los AGVs como conservadores de alimentos y productos agrícolas podría extenderse en México, por ejemplo, para conservar tomate, uva, fresa entre otros (Sholberg, 2009). La ventaja de usar AA en los empaques permitiría ahorro de agua e hipoclorito de sodio, ya que al combinar los AGVs y el hipoclorito, este último sería más eficiente para desinfectar frutos. El AA puede emplearse en dosis 3-5% donde se considera inocuo (Shafiur, 2007).
El AA podría eliminar algunas plagas al aplicarse en fase gaseosa, el ácido logra matar mosquita blanca y la palomilla del manzano Bemisiatabasi y Cydia pomonella, respectivamente (Randall et al., 2011; Samaniego-Gaxiola et al., 2012). Aunque en el caso de B. tabasi, habría que buscar la aplicación de AA en una forma innovadora, como su posible uso en solución.
El uso de AGVs podría combinarse con otros compuestos para incrementar su efecto letal; aún existe poca información al respecto. Sin embargo, los esclerocios del hongo P. omnivora murieron en 20 min al combinar AA o soluciones amortiguadoras que contienen este ácido y 1000 ppm del fungicida Propiconazole (Tilt®), en contraste, con los tres días que se necesitó para matar los esclerocios cuando solo se usó el fungicida; asimismo, Trichoderma sp. únicamente pudo atacar y matar a P. omnivora en presencia de AA (Samaniego-Gaxiola, 2008).
En alimentos en donde el AA se añadió, se observó una adaptación de las bacterias al ácido (Oh et al., 2009), ello significa un factor potencialmente limitante para el uso de los AGVs. Otros factores que interfieren con la toxicidad de los AGV son: i) el mecanismo de eflujo activo del AA que poseen algunas bacterias tolerantes al ácido (Mills et al., 2009); ii) la asimilación y posterior traslocación o remoción activa del AA (Mollapour y Piper, 2007); iii) la hipertonicidad en el medio, debida a una elevada concentración de NaCl (Chapman y Ross, 2009) y iv) la presencia de metionina en el medio, la cual contrarresta los efectos tóxicos del AA (Mills et al., 2009). Para contrarrestar los factores que limitan la toxicidad de los AGVs hacia los organismos, podrían cambiarse algunas condiciones fisicoquímicas del medio como el potencial Eh y sales, lo cual puede ser posible (Momma et al., 2011).
Recientemente, se analizaron trabajos, en donde se evaluó la efectividad que tiene la materia orgánica incorporada en el suelo para matar hongos que atacan raíces de plantas, ahí se destaca la necesidad de conocer los mecanismos por los que los hongos mueren, pues hay resultados inconsistentes al usar distintos tipos de materia orgánica y su efectividad para matar hongos (Bonanomi et al., 2007). En contraste, en la DAS se consideran a los AGVs como una constante, por ello, la investigación en este sentido podría redundar en un método más eficiente de uso generalizado, para el control de organismos fitopatógenos en el suelo. Adicionalmente, la DAS se enfoca a disminuir el uso de pesticidas en el suelo (Muramoto et al., 2008; Shennan et al., 2010).
La mayor parte de los suelos de México (principalmente en el norte del país) son alcalinos y limitarían el uso de los AGVs, si bien, la descomposición de carbohidratos y residuos vegetales en el suelo inundado induce la aparición de AGVs y una disminución del pH ~ 6, además una disminución adicional permanente del pH no es práctica, pues se necesitaría añadir ácido sulfúrico en grandes cantidades en el suelo. No obstante, una vez generados los AGVs en el suelo, al cambiar temporalmente el pH de ~ 6 a ~ 4 hizo que los esclerocios de P. omnivora murieran (datos no publicados). La idea de cambiar temporalmente el pH del medio para favorecer que los AGVs incrementen su toxicidad, puede estar acorde con el tiempo en el que los ácidos pueden ser letales hacia los microorganismos. Algunos ejemplos así lo indican: en 15 min 300 mmol L-1 nd de AA mataron más del 95% C. albicans (Ramsdale et al., 2008); los esclerocios de P. omnivora no sobrevivieron más del 10% al permanecer 15 min con 50 mmol L-1 nd de los ácidos fórmico y propiónico (Samaniego-Gaxiola y Nagamani, 2013).
La eficiencia de los AGVs para matar organismos parece ser elevada. El micelio de P. omnivora no sobrevivió a 38μg/ml de aire con AA, AB, AF o AP (Samaniego-Gaxiola y Nagamani, 2013) cantidad que es menor (~ 63μg ml-1 de aire) a la reportada para varios hongos fitopatógenos expuestos a una mezcla de 14 compuestos volátiles que se han detectado en el hongo Muscodor crispans (Mitchell et al., 2010). No obstante, se reportó que el aceite de orégano fue tan eficiente en matar esclerocios de Sclerotinia sclerotiorum que se necesitaron concentraciones menores a μg ml-1 (Soylu et al., 2007).
La inundación es una condición que favorece la formación de AGVs, pero niveles cercanos a la inundación también podrían serlo (Shennan et al., 2010); sin embargo, es necesario investigar más profundamente ambos casos para determinar en qué condición de humedad se favorece la mayor generación y eficiencia de AGVs para matar a organismos. Particularmente, las raíces de ciertas variedades de trigo poseen la capacidad de favorecer microbiotas desfavorables para organismos fitopatógenos y favorables para los cultivos (Mazzola, 2010), ello podría estar relacionado con los exudados de AGVs por las raíces, puesto que se sabe que plantas pueden exudar o acumular AA (Sun et al., 2011).
El pH no solo es determinante para las formas de los AGVs, también regula la biodiversidad de bacterias, explica el predomino de los hongos en suelo ácido y bacterias en suelos alcalinos (Rousk et al., 2010a y b). Así, el pH es un factor que al poderse manipular ayudaría a combatir microorganismos dañinos de humanos, animales y plantas (Peñalva et al., 2008). Aunque, en el suelo, el pH es dinámico y está influenciado por el suministro de agua, vegetación, temperatura, presencia de sales, entre otros (Zoltán, 2008).
Formas novedosas de utilizar los AGVs podrían involucrar la reutilización de desechos orgánicos producto de las actividades humanas y excretas de animales propios de la actividad pecuaria. Destaca también el uso reciente de los AGVs para el control de plagas de almacén (Randall et al., 2011; Sahrawat, 2008; Zoltán, 2008). Asimismo, cantidades abundantes de AGVs, particularmente AA y AF, podrían producirse en un sistema que acople la degradación de residuos orgánicos y la producción de AGVs (Díaz et al., 2009; Jin e Heiji, 2009) con su utilización como insumo de una cepa S. cerevisiae altamente eficiente en la producción de alcohol (Medina et al., 2010), o bien, que el ácido sea utilizado por bacterias metanógenas.
Conclusiones
Los efectos tóxicos de los ácidos grasos volátiles (AGVs) en suelo, agua y aire, los coloca como compuestos útiles para disminuir organismos perjudiciales para las actividades humanas. De tal manera que se podrían restringir microorganismos que contaminan alimentos, fumigar productos vegetales, preservar ensilados, granos y semillas, así como reducir poblaciones de malezas, nemátodos, hongos y bacterias que habitan el suelo y atacan a plantas. Por el costo, el ácido acético es económico y, éste y los otros AGV podrían generarse en grandes cantidades degradando residuos vegetales y animales u obtenerse de excretas de animales. El pH entre 4 a 5 favorecería la toxicidad de los AGVs hacia los organismos objetivo. Investigación concerniente a la fisicoquímica del suelo (pH, Eh, sales) podría aumentar el entendimiento y eficiencia de la aplicación de los AGVs.

