











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La transpiración es uno de los procesos que se ha estudiado en algunos cultivos en invernadero; un gran número de ecuaciones empleadas para estimarla se derivan de un balance de energía a nivel de follaje. La transpiración constituye un término importante del balance de energía del follaje y una medida significativa del rendimiento de un cultivo. Los principales estudios sobre transpiración de cultivos en invernadero se han realizado bajo las condiciones templadas de algunos países europeos, por ejemplo los Países Bajos. Sin embargo, estas condiciones no corresponden a los climas en los cuales se encuentran localizados los invernaderos en México.
En la estimación de la transpiración de cultivos en invernadero se ha utilizado el modelo de Penman- Monteith sin modificaciones (Jolliet y Bailey, 1992; Prenger et al., 2002) o con modificaciones (Boulard y Wang, 2000; Bontsema et al., 2007). La aplicación del modelo original o alguna de sus modificaciones ha dado buenas predicciones de la transpiración, aunque la validación no se ha hecho para todas las condiciones climáticas ni para todas las etapas del cultivo (índices de área foliar diferentes).
En México, existen pocas investigaciones sobre transpiración de cultivos en invernadero y se han realizado aún menos en los invernaderos ubicados en zonas áridas o semiáridas. En tales condiciones climáticas la transpiración, siendo el mecanismo más importante de disipación de energía y determinante de las condiciones térmicas del cultivo, puede cobrar una mayor relevancia. Lo anterior, aunado a la poca capacidad de retención de agua de los sustratos empleados en hidroponía, se requiere tener una buena predicción de la transpiración del cultivo en periodos cortos. Además, mantener en el invernadero un dosel con una tasa de transpiración alta constituye una de las formas más eficientes y baratas de enfriar el ambiente durante los días calurosos, con una radiación solar alta (Katsoulas et al., 2002). La advección es el transporte de masa y energía en el plano horizontal y en la dirección del viento (Rosenberg et al., 1983). En la naturaleza, la advección es la norma más que la excepción. Solamente cuando la superficie considerada es idéntica en color, rugosidad, disponibilidad de humedad y con una superficie infinita en contra del viento, prevalecerán condiciones de no advección. La presencia de advección de calor sensible puede inferirse del balance de energía. Si el flujo de calor latente es mayor que la energía disponible (radiación neta y flujo de calor del suelo), es decir, λE > (Rn + G), el calor sensible ha sido extraído del aire y consumido en la evaporación (Rosenberg et al., 1983). Con la modificación que McNaughton y Jarvis hicieron al modelo de Penman-Monteith, es posible calcular por separado los términos balance de energía y transferencia de vapor, añadiendo un factor de ponderación o desacoplamiento, el factor omega (McNaughton y Jarvis, 1983).
En el presente trabajo se comparó el desempeño de tres modelos matemáticos que estiman la transpiración de un cultivo en invernadero (Boulard y Wang, Penman-Monteith y Stanghellini), en dos sitios experimentales con plantas con área foliar diferente en dos épocas del año; además se utilizó el factor omega para determinar el acoplamiento entre el dosel del cultivo y la atmósfera circundante, y como indicador de advección. Considerando que el cultivo no sufrió déficit hídrico y transpiró a su tasa máxima.
Materiales y métodos
Modelos de Transpiración
Modelo de Penman-Monteith. Penman en 1948 fusionó dos teorías concernientes con la evaporación al reconocer que la estimación de la evapotranspiración requiere tanto de un componente termodinámico para el balance de energía en la superficie, como de un componente aerodinámico para la transferencia de vapor, con lo cual la evapotranspiración resulta una función de la radiación solar, temperatura del aire, presión de vapor y velocidad del viento (Monteith, 1998). Monteith (1981) amplió el campo de aplicación de la ecuación original de Penman al introducir resistencias variables para los flujos de momento, calor y vapor de agua, a través del sistema planta-atmósfera y con base en las características de la superficie como la resistencia estomática del cultivo y la resistencia aerodinámica, estableciendo así el modelo Penman-Monteith. Prenger et al. (2002), presentaron la ecuación de Penman-Monteith en los siguientes términos:
donde: λE es la densidad de flujo de calor latente (W m-2), E es la tasa de transpiración (kg s-1 m-2), λ es el calor latente de vaporización del agua (J kg-1) y se calcula conforme a la Ecuación (2) propuesta por American Society of Agricultural Engineers (ASAE Standards, 1998):
donde: Ta es la temperatura del aire (°C). δ es la pendiente de la curva de saturación de vapor de agua (Pa °C-1), se estima con la Ecuación (3) (ASAE Standards, 1998):
Rn es la radiación neta (W m-2) sobre el dosel del cultivo, y se estima así:
donde: τ es la transmisividad de la cubierta (adimensional), Rg es la radiación solar global (W m-2) y Ke es el coeficiente de extinción de la radiación (adimensional).
G es el flujo de calor del suelo (W m-2), se considera despreciable.
γ es la constante psicrométrica (Pa °C-1), se obtiene de la Ecuación (5) (Prenger et al., 2002):
donde: Cp es el calor específico del aire = 1010 J Kg-1 °C-1, MAgua es la relación de peso molecular vapor de agua-aire (0.6216), Patm es la presión atmosférica = 101325 Pa.
donde: ri es la resistencia estomática (s m-1), se calcula con la Ecuación (7) (Boulard et al., 1991):
donde: Di es el déficit de presión de vapor (Pa) dentro del invernadero.
ra es la resistencia aerodinámica (s m-1), se estima con la Ecuación (8) (Campbell, 1977):
donde: d es la longitud característica de la hoja (m), Vi es la velocidad del aire en el interior (m s-1), obtenida con la Ecuación (9) (Wang et al., 1999):
donde: Ac es el área de corte vertical del invernadero (m2), perpendicular a la dirección promedio del viento, o es el flujo de ventilación (m3 s-1) y se estima así (Kittas et al., 1995):
donde S0 es el área de ventilación del invernadero (m2), Cd es un coeficiente de descarga promedio de la ventilación (adimensional), C es un coeficiente de eficiencia relacionado con la velocidad del viento (adimensional) y V es la velocidad del viento en el exterior (m s-1).
ρ es la densidad del aire (kg m-3), se calcula (ASAE Standards, 1998):
ea es la presión de vapor actual (Pa), se estima a partir de la humedad relativa:
Modelo de Stanghellini. Stanghellini hizo una evaluación minuciosa del modelo de Penman-Monteith, revisó la ecuación combinada para representar las condiciones de transpiración en un invernadero. Además, propone un modelo en el que la resistencia estomática (interna) depende de: la radiación solar, el déficit de presión de vapor la hoja, la temperatura foliar y la concentración de bióxido de carbono. La siguiente ecuación es una simplificación del modelo de Stanghellini (Bontsema et al., 2007):
donde: E es la tasa de transpiración (kg m-2s-1), IAF es el índice de área foliar (m2m-2), ε es la relación (adimensional) entre calor latente y calor sensible que contiene el aire saturado ante un cambio de 1 °C, se calcula con la Ecuación (15):
ra es la resistencia aerodinámica (s m-1), Stanghellini a menciona que tomar un valor constante para esta variable afecta muy poco la precisión del modelo, ri es la resistencia estomática o interna (s m-1) y se calcula con la siguiente relación:
donde: Rn es la radiación neta sobre el dosel, la cual se estimó de la misma manera que en el modelo Penman- Monteith (Ecuación 4).
χa es la concentración actual de vapor de agua del aire (g m-3) y se calcula así:
donde: HR es la humedad relativa del aire (%).
λ es el calor latente de vaporización del agua (J g-1), se calcula con la Ecuación 2.
Modelo de Boulard y Wang. Con base en el modelo de Penman-Monteith y a partir de las condiciones meteorológicas al exterior del invernadero, Boulard y Wang (2000) obtuvieron la siguiente Ecuación:
donde: π es la eficiencia calorífica solar del invernadero (adimensional); Qh es el flujo de calor sensible del sistema de calentamiento (W m-2); G (W m-2) se consideró despreciable; el coeficiente total de pérdida de energía, Ks (Wm-2 K-1), se estimó de acuerdo con Bailey y Cotton (1980):
donde: A y B son coeficientes empíricos.
KH y Kv son los coeficientes de transferencia de calor sensible y calor latente por ventilación:
donde: Ag es el área del suelo del invernadero (m2), ξ es el factor de conversión entre el contenido de vapor de agua en el aire a temperatura estándar y la presión de vapor de agua (kilogramo de agua (kgw) por kilogramo de aire (kga-1)).
donde: ri, la resistencia estomática, se calculó con la Ecuación siguiente (Boulard et al., 1991):
ra es la resistencia aerodinámica (s m-1), se estimó con la Ecuación 8.
Do es el déficit de presión de vapor de agua (Pa), afuera del invernadero.
La programación de los modelos se realizó en el ambiente Matlab-Simulink versión 6.5 (MathWorks Inc, 2002).
Factor omega. La modificación que McNaughton y Jarvis (1983) hicieron al modelo de Penman-Monteith permite calcular por separado los términos balance de energía y transferencia de vapor, añadiendo un factor de ponderación o desacoplamiento, el factor omega (Ω), que adopta valores entre 0 y 1. El acoplamiento entre dosel y atmósfera lo describen en términos del déficit de saturación de vapor de agua. El factor Ω se calcula con la siguiente Ecuación:
Este factor se basa en la relación entre resistencia estomática y resistencia aerodinámica, dicha relación determina la distribución de la evapotranspiración entre balance de energía y transferencia de vapor, con base en el grado de desacoplamiento de las condiciones ambientales. Cuando ra es mucho mayor que ri en grado tal que Ω tiende a 1, el flujo de calor latente lo determina principalmente el término de balanc de energía,
Medición de Variables Meteorológicas y Transpiración del Cultivo
Los experimentos se realizaron en dos invernaderos con cubierta de plástico. El primero, tipo capilla, se localiza en el campus Querétaro de la Universidad Autónoma de Querétaro, cuyas coordenadas son 20º 36’ N y 100º 24’ O, a 1820 m de altitud. El invernadero tiene 37 m en la dirección norte-sur y lo constituyen cuatro naves de 6.75 m de ancho, con una altura cenital de 5 m y una altura a canaleta de 3.5 m. Cuenta con dos ventanas laterales (2.5 m × 25 m), dos ventanas frontales (2.5 m × 20 m) y dos ventanas cenitales (0.8 m × 20 m), con un área total de ventilación de 257 m2. El segundo, se localiza en el Campo Experimental de la Universidad Autónoma de Chapingo, Chapingo, Estado de México, localizada a los 19º 29’N y 98º 54’O, a 2244 m de altitud. Este invernadero es tipo sierra, con 76 m en la dirección norte-sur y está constituido por tres naves de 8.5 m de ancho; tiene una altura cenital de 6 m y una altura a canaleta de 4 m. Tiene dos ventanas laterales (3 m × 73 m), dos ventanas frontales (3 m × 23 m), tres ventanas cenitales (1.6 m × 76 m), y un área total de ventilación de 940 m2. Ambos invernaderos tienen malla antiinsectos y pueden abrirse y cerrarse manual o automáticamente. Además cuentan con un sistema automatizado de riego por goteo.
En los dos invernaderos se cultivo jitomate en hidroponía. En el experimento de Querétaro se usó el cultivar híbrido “Reserva” tipo saladette, de crecimiento indeterminado; para el trasplante (realizado el 30 de abril de 2008) se utilizó el sustrato perlita en bolsas individuales a hilera doble, a una densidad de 2.5 plantas m-2. En el experimento de Chapingo se usó el cultivar híbrido Caimán tipo bola, de crecimiento indeterminado. Las plantas se colocaron en bolsas largas (“salchichas”) rellenas del sustrato perlita, a una densidad de 3.1 plantas m-2. En cada invernadero se instaló un lisímetro de pesada (QA150FEG-S, Sartorius, Germany) con capacidad de 150 kg y 2 g de precisión. Los lisímetros se configuraron para registrar datos cada minuto; en la comparación de los modelos se utilizó la transpiración acumulada cada 30 minutos. Los datos de los lisímetros se procesaron de acuerdo con el procedimiento descrito por Meurs y Stanghellini (1992).
El lisímetro en el invernadero de Querétaro soportó cuatro plantas, los registros se hicieron del 6-8, 14-17 y 19-24 de junio 2008. Durante los mismos periodos se midieron: temperatura del aire (ºC), humedad relativa (%), radiación solar global (W m-2) y velocidad del viento (m s-1), cada minuto dentro y fuera del invernadero. El lisímetro en el invernadero de Chapingo soportó cinco plantas y se obtuvieron datos del 24-29 de octubre y del 31 de octubre al 11 de noviembre 2008. En esta localidad se registraron cada minuto dentro del invernadero: temperatura del aire (ºC), humedad relativa (%) y radiación fotosintética activa (μE), y fuera del invernadero sólo temperatura del aire (°C).
La medición de las variables, en ambos sitios, se hizo con una estación meteorológica automática (H21-001, HOBO, Onset Computer Corp., USA). Para las simulaciones se usaron promedios cada 30 minutos de todas las variables. En el sitio Chapingo, las variables de entrada que requiere el modelo de Boulard y Wang: humedad relativa (%), radiación global (W m-2) y velocidad del viento (m s-1), medidas afuera del invernadero cada 30 minutos, se obtuvieron de la Estación Meteorológica de la Universidad Autónoma Chapingo.
El área foliar en ambos sitios se obtuvo con un método destructivo en cada periodo de medición. En el experimento de Querétaro, el área foliar se midió con un integrador de área foliar (LAI-2000, LI-COR Inc., USA) mientras que en el experimento en Chapingo se obtuvo un modelo de estimación del área foliar a partir de mediciones de largo y ancho de las hojas, y con éste se calculó el índice de área foliar.
Resultados y discusión
Para el modelo Penman-Monteith se tomaron los valores 0.7 (Bontema et al., 2007), 0.15 (Boulard et al., 2002), 0.644 y 0.09 (Boulard y Baille, 1995), correspondientes al coeficiente de extinción de la radiación (Ke), longitud característica de la hoja (d), coeficiente de descarga de la ventilación (Cd) y coeficiente de eficiencia del viento (C), respectivamente. La transmisividad de la cubierta (τ) se calculó a partir de la relación entre la radiación global medida dentro y fuera del invernadero (τ = 0.62). En el modelo de Stanghellini se usó un valor fijo para la resistencia aerodinámica (ra = 200 s m-1), ya que la baja velocidad del viento dentro del invernadero hace que ra sea una constante, y afecte muy poco la precisión del modelo (Bontsema et al., 2007). Para el modelo de Boulard y Wang, el valor de ξ, factor de conversión entre el contenido de vapor de agua en el aire a temperatura estándar y la presión de vapor de agua, fue 6.25 × 10-6 kgw kga-1 Pa-1 (Boulard y Wang, 2000). Los valores de los parámetros anteriores se usaron en ambos sitios experimentales. Los índices de área foliar (IAF) calculados para los tres periodos de estudio, 6-8, 14-17 y 19-24 de junio 2008, en el sitio Querétaro fueron 0.7, 1.0 y 1.2, respectivamente. En el sitio Chapingo solo se obtuvo un valor del índice de área foliar (IAF =2.0).
Con base en las salidas obtenidas del ambiente Matlab-Simulink (MathWorks Inc, 2002), para el sitio Querétaro los modelos Penman-Monteith y Stanghellini subestimaron sistemáticamente la transpiración (Figura 1 y 2, Cuadro 1). Una posible causa de esta subestimación es la presencia de advección, debida a la fracción de suelo desnudo (dosel disperso, IAF: 0.7, 1.0 y 1.2) y seco entre el dosel, que generó una carga extra de calor sensible, la cual provocó una transpiración superior a la calculada mediante el balance de energía. Seginer (1994), reporta que un dosel disperso se expone a la advección local del suelo desnudo circundante, ya que la temperatura de plantas sin estrés hídrico y dispersas probablemente es menor que la temperatura del aire, debido a la tasa de transpiración alta (Yang et al., 1989), lo que causa que el flujo de calor latente exceda la radiación neta.
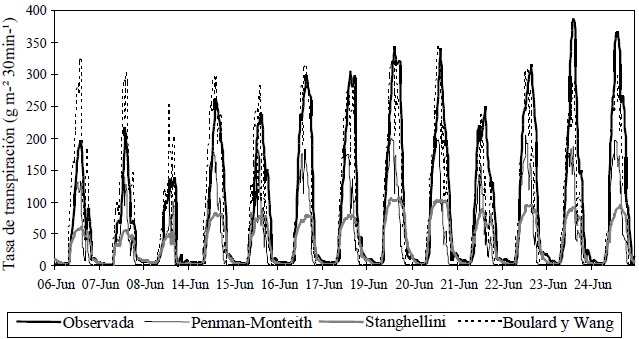
Figura 1 Estimación, utilizando tres modelos, de la tasa de transpiración de un cultivo de jitomate en invernadero, en la Universidad Autónoma de Querétaro, Campus Querétaro.
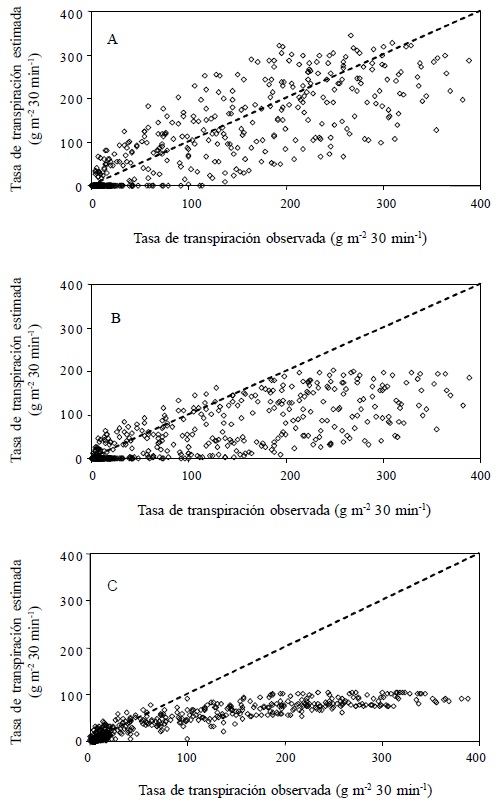
Figura 2 Tasa de transpiración de un cultivo de jitomate, calculada con tres modelos: Boulard y Wang (A), Penman-Monteith (B) y Stanghellini (C); en un invernadero de la Universidad Autónoma de Querétaro, Campus Querétaro. Desde los 37 hasta los 55 días después del trasplante (6 al 24 de junio); línea 1:1 (- - -), n = 624.
Cuadro 1 Medidas estadísticas utilizadas para evaluar tres modelos con los que se estimó la transpiración de un cultivo de jitomate en invernadero en dos sitios experimentales.
| Sitio | Modelo | Medida estadísitica | ||||
|---|---|---|---|---|---|---|
| r2 | IC | EF | RECM | EAM | ||
| Universidad Autónoma de Chapingo | Boulard y Wang | 0.847 | 0.772 | -1.467 | 82.828 | 52.77 |
| Penman-Monteith | 0.494 | 0.763 | -0.579 | 66.277 | 43.806 | |
| Stanghellini | 0.862 | 0.961 | 0.844 | 20.856 | 14.324 | |
| Universidad Autònoma de Querétaro | Boulard y Wang | 0.742 | 0.923 | 0.722 | 54.757 | 35.257 |
| Penman-Monteith | 0.652 | 0.778 | 0.418 | 79.245 | 47.996 | |
| Stanghellini | 0.832 | 0.639 | 0.254 | 89.759 | 53.205 | |
r2 = coeficiente de determinación; IC = índice de concordancia (Willmot, 1981); EF = eficiencia dl modelo (Nash y Suteliffe, 1970); RECM = raíz del error cuadrático medio; EAM = error absoluto medio.
La temperatura del aire dentro del invernadero superó la temperatura del aire en el exterior (Figura 3), esto es, una carga extra de calor sensible Esta carga extra, como se mencionó, puede ser consecuencia de lo disperso del dosel y de que éste no genera el flujo de calor latente necesario para enfriar el aire del invernadero, lo que provoca que el déficit de presión de vapor sea mayor en el interior.
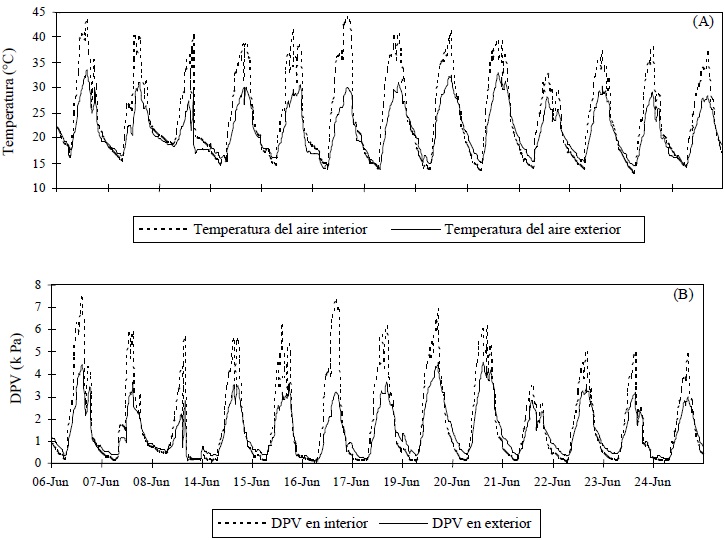
Figura 3 (A) Temperatura del aire registrada dentro y fuera del invernadero. (B) déficit de presión de vapor (DPV) calculado. Sitio Universidad Autónoma de Querétaro, Campus Querétaro.
Para detectar advección se usó el factor omega (Ecuación 26 en la sección de metodología). En la Figura 4 se presentan los valores de este factor, calculado con base en las variables meteorológicas registradas dentro del invernadero e incluido en el programa del modelo Penman-Monteith. En dicha figura se muestra que el factor omega fue prácticamente 1. Esto implica que la resistencia aerodinámica fue mayor a la resistencia estomática, el factor omega tiende a 1, y el déficit de saturación de vapor sobre el dosel alcanzó su valor mediante un equilibrio local (McNaughton y Jarvis, 1983). Esto es, la transpiración debió estar determinada por el balance de energía, sin embargo, las condiciones internas no reflejaron advección porque la ecuación para estimar velocidad del viento en el interior (Ecuación 9) subestima esta variable (Wang et al., 1999), y ésta a su vez es importante en el cálculo de la resistencia aerodinámica. Otro hecho que influye en la presencia de advección, es la localización del invernadero en una región con clima semiárido y la falta de una cortina rompevientos u otros invernaderos alrededor, los cuales disminuirían la velocidad del viento; con tales obstáculos el valor del factor omega se aproximaría a 1 (Jarvis, 1985). Con respecto al modelo de Stanghellini, la subestimación de la transpiración puede deberse al uso de un valor constante para la resistencia aerodinámica, aunado al que el modelo fue desarrollado en otras condiciones ambientales (clima templado en los Países Bajos), con un IAF mayor (2.2 < IAF < 4.2).
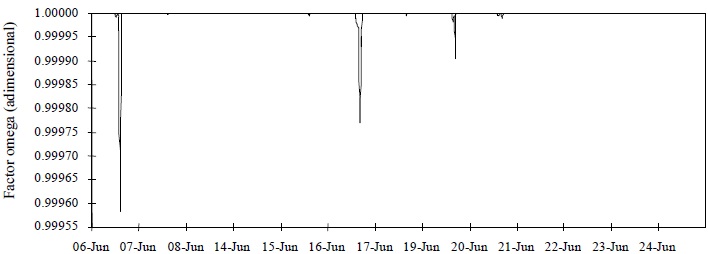
Figura 4 Factor omega (Ω) calculado con base en las variables meteorológicas registradas dentro del invernadero en la Universidad Autónoma de Querétaro, Campus Querétaro.
El modelo de Boulard y Wang tuvo el mejor desempeño con base en las medidas estadísticas, excepto r2 (Cuadro 1). La discrepancia entre r2 y las otras medidas coincide con lo expuesto por Willmott (1982), quien menciona que esta medida no es la más adecuada cuando se comparan datos observados y estimados. La Figura 1 y 2 muestran el mejor ajuste del modelo de Boulard y Wang durante el periodo de estudio para el sitio Querétaro. El mejor desempeño del modelo puede ser resultado de la utilización de variables meteorológicas registradas fuera del invernadero, las cuales reflejaron la presencia de advección. En la Figura 5 se presenta el comportamiento del factor omega fuera del invernadero, se observa una variación mucho mayor de los valores del factor Ω, respecto a los valores obtenidos utilizando las variables meteorológicas medidas dentro del invernadero (Figura 4).
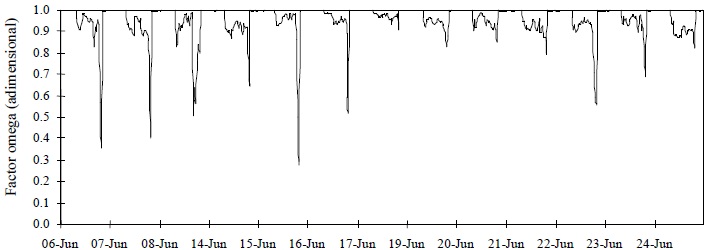
Figura 5 Factor omega (Ω), calculado con base en las variables meteorológicas registradas fuera del invernadero, Universidad Autónoma de Querétaro, Campus Querétaro.
La resistencia aerodinámica fue menor que la resistencia estomática, el factor omega tiende a 0 y el déficit de saturación de vapor sobre el dosel es impuesto por el estado del aire que pasa sobre el dosel. En este caso, la advección de aire caliente y seco incrementó la tasa de transpiración (la advección de aire húmedo puede deprimirla) volviéndola fuertemente dependiente de la resistencia estomática y del déficit de saturación de vapor de agua en el aire (Jarvis, 1985).
En el sitio Chapingo se apreció que el modelo de Boulard y Wang (B-W) fue el que más sobrestimó la transpiración del cultivo (Figura 6), a diferencia de los resultados obtenidos por estos autores, quienes reportaron una subestimación sistemática de la transpiración, también en un cultivo de jitomate.
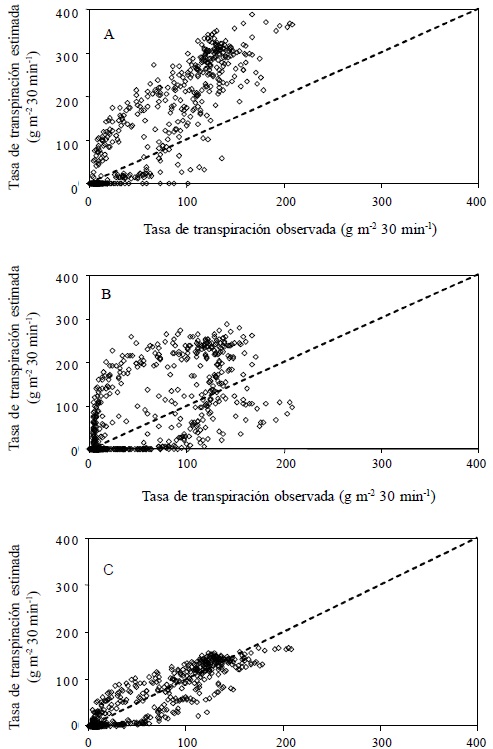
Figura 6 Tasa de transpiración de un cultivo de jitomate, calculada con tres modelos: Boulard y Wang (A), Penman-Monteith (B) y Stanghellini (C); en un invernadero de la Universidad Autónoma Chapingo, Chapingo, Estado de México. Durante los días 24-29 de octubre y del 31 de octubre al 11 de noviembre 2008. Línea 1:1 (- - -), n = 864.
La sobrestimación se atribuye a la utilización de registros de velocidad del viento y humedad relativa de la estación meteorológica de la Universidad Autónoma Chapingo, cuyos valores no reflejaron las condiciones micrometeorológicas que prevalecieron alrededor del invernadero. La sobrestimación de la transpiración con el modelo Penman-Monteith (P-M) fue menor a la del modelo de B-W, aunque el modelo P-M tuvo un r2 y un índice de concordancia (IC) menores; las otras medidas estadísticas indican un mejor ajuste (Cuadro 1), principalmente la RECM y el EAM; Willmott (1982) considera que éstas dos medidas estadísticas están entre las mejores medidas de desempeño de un modelo. El r2 del modelo P-M es similar al obtenido por Prenger et al. (2002), de 0.481, quienes también reportaron la sobrestimación de la transpiración en plantas de arce rojo con este modelo, al igual que Jolliet y Bailey (1992) en un cultivo de jitomate. El modelo de Stanghellini presentó el mejor ajuste en el sitio experimental Chapingo, como se observa en la Figura 6 y en las medidas utilizadas para evaluar el desempeño (Cuadro 1). El mejor desempeño del modelo de Stanghellini puede atribuirse a una mejor estimación de la resistencia estomática, dado que en el cálculo del coeficiente de absorción de la radiación, la otra diferencia principal con el modelo P-M (Jolliet y Bailey, 1992), se usó la misma Ecuación 4 del modelo P-M. Además, el índice de área foliar obtenido (IAF = 2) está muy cercano al rango que utilizó Stanghellini para desarrollar el modelo (2.2 < IAF < 4.2). En la Figura 7 se presenta el comportamiento de los tres modelos durante el periodo de estudio (18 días) para el sitio Chapingo.
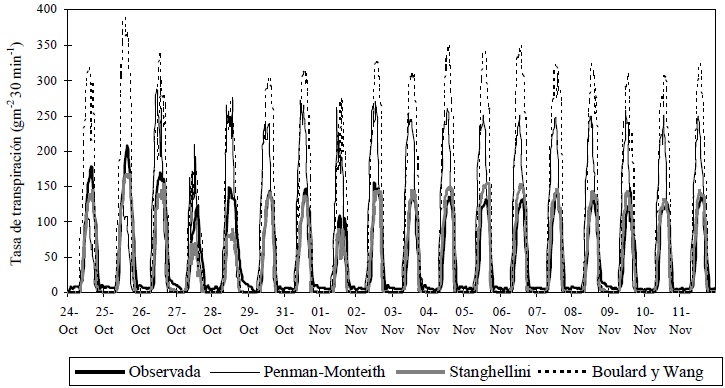
Figura 7 Estimación de la tasa de transpiración de un cultivo de jitomate en invernadero con tres modelos. Universidad Autónoma de Chapingo, Chapingo, Estado de México.
En el sitio Chapingo, con un dosel denso (IAF = 2), el flujo de calor latente fue suficiente para que la temperatura del aire en el interior resultase igual o menor a la temperatura del aire afuera del invernadero (Figura 8), lo cual ocasionó un déficit de presión de vapor en el interior (aproximadamente un 50% y algunos días un porcentaje menor) menor que en el sitio Querétaro. El resultado anterior se corresponde con lo mencionado por Katsoulas et al. (2002): mantener un índice de área foliar alto implica una mayor transpiración, y ésta a su vez modifica las condiciones ambientales del invernadero, principalmente temperatura del aire y déficit de presión de vapor.
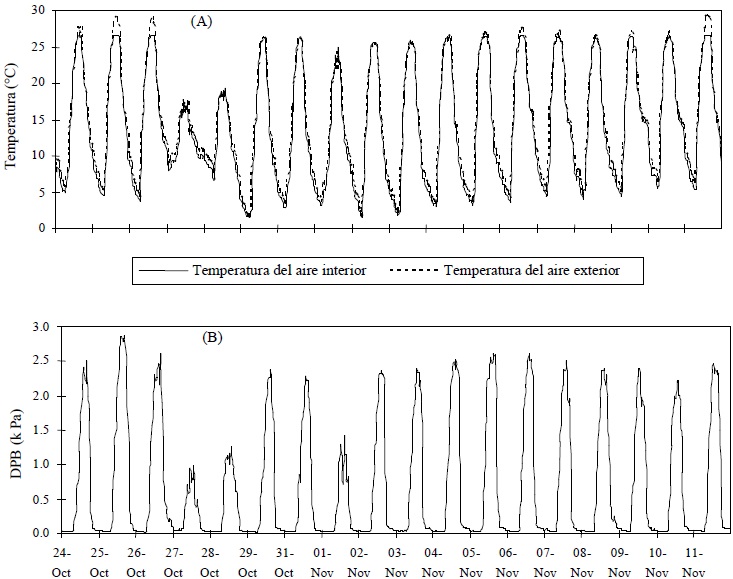
Figura 8 (A) Temperatura del aire registrada dentro y fuera del invernadero, (B) déficit de presión de vapor (DPV) en el interior, calculado. Universidad Autónoma Chapingo, Chapingo, Estado de México.
Con base en los resultados y discusión anteriores, ninguno de los tres modelos tuvo un buen desempeño en ambos sitios y condiciones experimentales, por lo que es necesario hacer más validaciones de los modelos en ambos ambientes.
Conclusiones
El modelo de Stanghellini fue el mejor para estimar la tasa de transpiración de un cultivo de jitomate, en el clima templado de la Universidad Autónoma Chapingo, durante el otoño, con un dosel cerrado (índice de área foliar de 2) y en un invernadero rodeado por otros invernaderos, que actuaron como cortina rompevientos. Por otra parte, los modelos Penman-Monteith y Boulard-Wang tuvieron un menor ajuste, sobrestimaron la transpiración. Para el sitio de la Universidad Autónoma de Querétaro, con el modelo de Boulard y Wang se hizo la mejor estimación de la transpiración; en un clima semiárido, durante el verano, con un dosel disperso (índice de área foliar de 1) y en un invernadero aislado. En este caso, los modelos Penman-Monteith y Stanghellini subestimaron la transpiración sistemáticamente; debido a la presencia de advección, provocada por la fracción de suelo desnudo y las condiciones ambientales circundantes, detectada con el factor omega. Para extrapolar el desempeño de los modelos utilizados a condiciones climáticas similares a las estudiadas, es necesario hacer más validaciones.

