











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los elementos de las tierras raras (ETR) se consideran benéficos para el metabolismo vegetal, (Andrès et al., 2003). El suministro de ETR en pequeñas cantidades, particularmente lantano (La) y cerio (Ce) incrementan el crecimiento, la calidad y el rendimiento de diversos cultivos (Zhang et al., 2003; Hu et al., 2006). También se ha demostrado que el La afecta distintos procesos y reacciones fisiológicas, así como absorción, transporte y asimilación nutrimental (Pang et al., 2002; Tyler, 2004). La influencia del La sobre la absorción de nutrimentos puede deberse a que al ser el más electropositivo de los ETR, resulta muy similar en sus propiedades químicas a los elementos alcalino-térreos, entre los que se encuentran el Ca y Mg (Feyerabend et al., 2008). Ejemplo de lo anterior son los radios iónicos semejantes, es decir, el del ión Ca es de 9.9 nm, en tanto que el del La de 11.6 nm (Hu et al., 2004).
El efecto que en particular tiene el La sobre algunos nutrimentos esenciales como K, Ca y Mg ha sido estudiado en diversas especies. En cultivos como algodón (Gossypium hirsutum L.), trigo (Triticum aestivum L.) y arroz (Oryza sativa L.), el suministro de La favorece la absorción de K (Wahid et al., 2000). Por otra parte, Dong et al. (1992), demostraron que la aplicación de cloruro de lantano incrementó la actividad de las K+ y Mg+2-ATPasas de la membrana celular bajo condiciones de deficiencia de calcio. Además, los efectos del La en la absorción iónica, son comparables a aquellos que induce el Ca, lo cual sugiere que el lantano afecta la regulación de los niveles celulares de Ca (Hu et al., 2004). De ahí que el La puede desplazar al Ca en los sitios de unión extra-celulares, inhibiendo el influjo y eflujo celular del Ca (Diatloff et al., 2008).
Si se consideran las propiedades químicas del La y los efectos fisiológicos que éste ocasiona en las plantas, es posible notar funciones parecidas a las que presentan tanto Ca como K (D’Aquino et al., 2009). El La, al ser un catión y presentar propiedades químicas similares a los elementos Ca y K, puede tener efectos sobre la absorción y acumulación de éstos, los cuales son nutrimentos de suma importancia para el cultivo de tulipán (Tulipa gesneriana L.); el primero incrementa la altura de la planta y peso de materia seca, favorece la firmeza del tallo, mejora la síntesis y migración de glúcidos hacia el bulbo y la coloración de los pétalos, en tanto que su deficiencia genera aborto de yemas florales y se produce la fisiopatía denominada “vuelco de tulipán”, la cual se caracteriza por el doblamiento del pedúnculo a la altura del cuello de la planta, al formarse la flor (Nelson et al., 2003). Por su parte, el K favorece la rigidez del tallo y está implicado en el crecimiento celular y la estabilización de la membrana y de la pared celular (Armstrong, 2002). Así también, se ha reportado que el lantano puede ser considerado benéfico para diversos cultivos (Hu et al., 2004). En este contexto, esta investigación planteó como objetivo evaluar el efecto de fuentes y concentraciones de lantano en la solución nutritiva sobre la acumulación de K, Ca y La en hojas por planta de tulipán y determinar las relaciones entre el suministro de La y la acumulación de K, Ca y La, expresadas como coeficientes.
Materiales y métodos
Ubicación del Experimento y Condiciones Experimentales
La investigación se realizó en el Colegio de Postgraduados, Campus Montecillo, México (19° 29’ N, 98° 53’ O y una altitud de 2250 m) en invernadero. La plantación de bulbos de tulipán cultivar Ille de France, calibre 12+, se realizó empleando un sustrato correspondiente a una mezcla 70/30 (v/v) de tezontle de 3 mm (roca volcánica porosa, pH de 7.3 y CE de 0.15 dS m-1) con Promix® (combinación de helecho Sphagnum, perlita, vermiculita, pH de 4.7 y CE de 1.4 dS m-1) (Ramírez et al., 2010). Antes de la siembra, los bulbos se limpiaron y desinfectaron, sumergiéndolos por 15 min en una solución fungicida compuesta por Ridomil® (Metalaxyl-m) 2 g L-1 más Captan® (N triclorometiltio-4-ciclohexeno-1,2-dicarboximida) a razón de 1 g L-1.
Una vez que las plantas emergieron, las macetas se regaron cada tercer día, con la solución nutritiva universal de Steiner (Steiner, 1984) al 50%, complementada con micronutrimentos en las siguientes concentraciones (mg L-1): 1.6 de Mn; 0.11 de Cu; 0.86 de B; 0.023 de Zn y 5.0 de Fe. Los micronutrimentos Mn, Cu y Zn fueron suministrados en forma de sulfatos; B como H3BO3, y Fe como quelato (Fe-EDTA), de acuerdo a lo descrito por Steiner y van Winden (1970). El pH de la solución fue ajustado a 5.5 con HCl 1M. Las soluciones nutritivas fueron preparadas con reactivos grado analítico.
Diseño Experimental y Tratamientos
Se utilizó un diseño experimental factorial 6 × 2 (concentración y fuente de La en la solución nutritiva, respectivamente), con tres repeticiones, distribuidas completamente al azar. Las unidades experimentales fueron macetas de 15.2 cm de diámetro conteniendo una planta. Las fuentes de La fueron LaCl3 y La(NO3)36H2O, en tanto que los niveles probados fueron 0, 5, 10, 20, 30 y 40 μM de La, en cada caso, respectivamente.
Variables Evaluadas
Acumulación nutrimental. La acumulación nutrimental en hojas fue estimada a partir del peso de biomasa seca de éstas y de las concentraciones nutrimentales de Ca, K y La en hoja. Para la obtención de la biomasa seca en hojas, éstas fueron separadas del resto de la planta inmediatamente después del corte de los tallos florales (65 días después de la plantación) y se secaron hasta peso constante a 70 °C en una estufa de aire forzado. Para la determinación de las concentraciones nutrimentales, las muestras de hojas secas fueron molidas utilizando un molino equipado con hojas de acero inoxidable con malla de 2 mm. En este material molido se determinaron las concentraciones de Ca, K y La, mediante digestión húmeda del material seco con una mezcla de ácidos perclórico y nítrico (Alcántar y Sandoval, 1999). La lectura de los extractos obtenidos después de la digestión y filtrados se determinaron en el equipo de espectroscopía de emisión atómica de inducción por plasma acoplado ICP-AES VARIAN® Liberty II. El límite de detección para La fue de 0.002 μg L-1 con una longitud de onda 407.7 nm.
Coeficientes de determinación. Los coeficientes de determinación entre el suministro de La con dos fuentes y las acumulaciones de K, Ca y La, se calcularon derivando la función racional y polinomial de tercer grado, misma que permite aproximar la precisión en el ajuste de los datos originales (Hunt, 2003).
Análisis Estadístico
Con los resultados de las acumulaciones de K, Ca y La se realizó un análisis de varianza y una prueba de comparación de medias utilizando la prueba estadística LSD (Least significant difference, P ≤ 0.05), mediante el programa estadístico SAS versión 9.0 (SAS Institute, 1990).
Resultados y discusión
Acumulación Nutrimental
Efecto de la concentración de La en la solución nutritiva. No se presentaron síntomas visuales de deficiencias nutrimentales en las plantas de tulipán tratadas con La, como es el caso de deficiencias de N y P reportadas en arroz tratado con este elemento (Zeng et al., 2006), o de K, Ca y Mg reportadas en presencia de concentraciones relativamente bajas de La en frijol (von Toucher et al., 2001).
Se observaron diferencias significativas (LSD, P ≤ 0.05) en la acumulación nutrimental de K, Ca y La en hojas por planta de tulipán por efecto del factor concentración de La en la solución nutritiva (Figura 1). Las mayores acumulaciones de K y Ca (32.36 mg K hojas-1 por planta y 8.38 mg Ca hojas-1por planta, respectivamente), se registraron en plantas crecidas en soluciones con 10 y 20 µM de La, respectivamente. Asimismo, con la mayor concentración de La evaluada (40 µM) se observó un incremento en las acumulaciones de K y Ca, con respecto al testigo, aunque este incremento es menor al registrado en el tratamiento con menor concentración de La (5 µM). Contrario a lo aquí obtenido, Xie et al. (2002) reportaron que el suministro de La a partir de 9 mg L-1 disminuye la absorción de Ca en grano y paja de arroz. En este mismo sentido, Szczerba et al. (2009), demostraron que los efectos inhibitorios del La sobre la absorción de K son atribuidos a la asociación del La a sitios de unión a K en las membranas celulares.
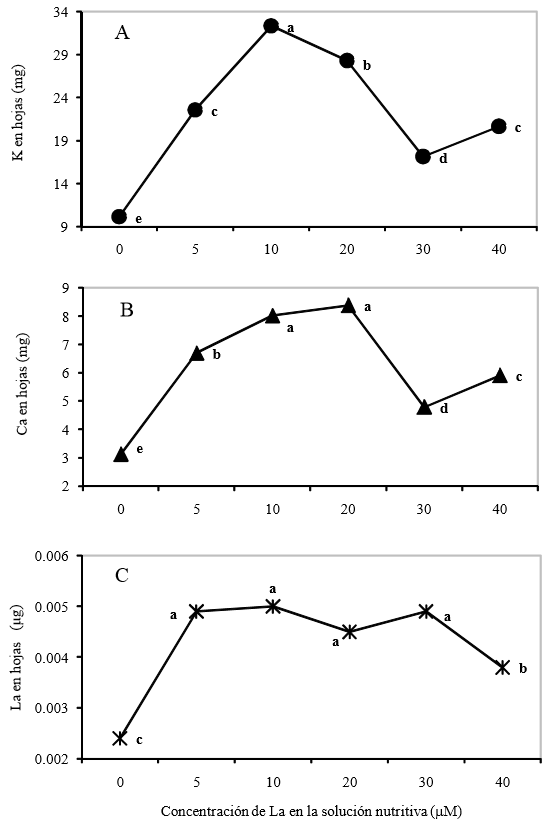
Valores con letras distintas en cada elemento son estadísticamente diferentes (LSD, P ≤ 0.05).
Figura 1. Acumulación de potasio (K) (A), calcio (Ca) (B) y lantano (La) (C) en hojas por planta de tulipán cultivar Ille de France con diferentes concentraciones de lantano.
La capacidad de los iones La para estimular el sistema de transporte de Ca ha sido reportado en células animales, dado que el La3+ estimula el influjo de Ca2+ a través del plasmalema por la inducción de un cambio conformacional de un transportador de Ca2+que promueve la unión de Ca con esta proteína transportadora o con los fosfolípidos del plasmalema (Li et al., 1999). Los resultados obtenidos en esta investigación, sugieren que en células vegetales, el La3+ pudiera también ocasionar cambios conformacionales en proteínas transportadoras de cationes que incrementan la capacidad de absorción de éstos.
Es importante notar efectos diferenciales en la acumulación de Ca y K en función de la concentración de La adicionada (Figura 1A y 1B), observándose que el efecto positivo del La sobre la absorción de estos cationes esenciales disminuye a medida que su concentración en la solución nutritiva aumenta. Tendencias similares han sido reportadas por Wang et al. (2012) en plántulas de haba (Vicia faba L.) con curvas dosis-respuesta en forma de U, observándose contenidos ascendentes de Ca, K y Fe en raíces tratadas con La en un intervalo de 7 a 57 µM de La (1 a 8 mg de La L-1 de solución nutritiva). Asimismo, Xie et al. (2002) mencionan que el suministro de La incrementa el área foliar y masa radical, generando una mayor absorción de Ca, K y otros nutrimentos.
Zhimang et al. (2001) demostraron que la bioacumulación de elementos de las tierras raras en plantas es mucho mayor en raíces que en la parte aérea. No obstante, la similitud en los patrones de bioacumulación observado en las raíces y vástagos, sugirieron que estos elementos pueden ser transportados desde el suelo hasta la parte aérea, previa absorción radical (May et al., 1997), lo cual coincide con los resultados obtenidos en esta investigación, pues se identificó La en parte aérea (Figura 1C).
Los resultados de acumulación de La en hojas por planta pueden dividirse en tres grupos estadísticamente diferentes: el primero, ausencia de La; el segundo, intervalo de concentración de 5 a 30 μM; y por último el tratamiento con 40 µM. En experimentos realizados en suelo, se reporta acumulación de La tanto en vástagos como en raíces, cuando los suministros de lantano son bajos (0 a 10 mg kg-1 de suelo) y éstos permanecen constantes aun cuando se incremente la dosis de lantano (Zhang y Shan, 2001).
Efecto de la fuente. Las fuentes de La evaluadas ocasionaron respuestas estadísticas diferentes (DMS, P ≤ 0.05). Las mayores acumulaciones de K, Ca y La se observaron con LaCl3 como fuente. Las cantidades de K, Ca y La encontradas en hojas tratadas con La(NO3)36H2O representaron sólo el 35.2, 39.8 y 37%, de las obtenidas con el suministro de LaCl3, respectivamente (Cuadro 1). Los resultados obtenidos muestran que la fuente de La3+ suministrada a la solución nutritiva tiene la capacidad modificar la acumulación de cationes esenciales como K y Ca.
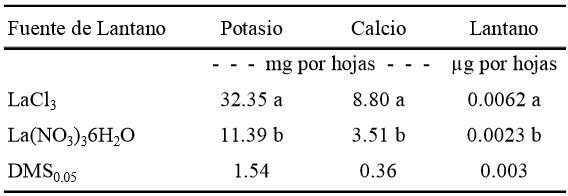
Valores con letras distintas en cada columna son estadísticamente diferentes (DMS, P ≤ 0.05). LSD0.05 = diferencia mínima significativa al 5 %.
Cuadro 1. Acumulación de K, Ca y La en hojas por planta de tulipán cultivar Ille de France, tratado con dos fuentes de La.
En los resultados que se presentan en el Cuadro 1 es posible observar un efecto sinérgico del Cl- como anión acompañante, sobre la absorción de K, Ca y La.
La sinergia del cloro sobre la absorción catiónica puede ser explicada por la existencia de cotransportadores catión-cloro (CCC) que catalizan el simporte coordinado de K+ , Na+ y Cl- (Colmenero-Flores et al., 2007). Recientemente, Kong et al. (2011) reportaron que la proteína CCC está involucrada en el transporte y la homeostasis tanto de K+ como de Cl-. White y Broadley (2001) explican que el Cl- también actúa como un contranión, y los flujos de Cl- están implicados en la estabilidad del potencial de membrana, la regulación de los gradientes del pH intracelular y la excitabilidad eléctrica.
Además, se ha reportado que el NO3- es inestable y puede inmovilizarse más fácilmente en materiales orgánicos que el Cl-, lo que en consecuencia ocasiona una mayor actividad de los cationes en solución (Jakobsen, 1992); en esta investigación el sustrato empleado poseía un componente orgánico. Ejemplo de lo anterior fue reportado por Weggler-Beaton et al. (2000), quienes demostraron que la concentración creciente de Cl- en la solución del suelo favoreció la absorción de Cd2+ en trigo (Triticum vulgare L.) y en acelga (Beta vulgaris L.) en suelos enmendados con biosólidos. Sin embargo, en tal investigación no se separaron los efectos de Cl sobre el Cd originado por los biosólidos, de los efectos sobre el Cd contenido en el suelo agrícola utilizado. Se concluyó que la formación de complejos entre el Cl y el Cd incrementó la biodisponibilidad de Cd en suelos enmendados con biosólidos.
Interacción concentración por fuente. En el Cuadro 2 se presentan los efectos de la interacción de los factores evaluados. En general, las acumulaciones de K, Ca y La fueron mayores cuando las plantas de tulipán fueron tratadas con La, independientemente de la fuente utilizada. No obstante, es evidente que concentraciones de La iguales o superiores a 30 µM inhiben la acumulación de los elementos en estudio. Como fue mencionado previamente, es altamente probable que en células vegetales, el La3+ pudiera también ocasionar cambios conformacionales en proteínas transportadoras de cationes que incrementan la capacidad de absorción de éstos; no obstante, Wang et al. (2012) demostraron que el efecto del La sobre la absorción de cationes es dependiente de la concentración de éste; concentraciones elevadas de La contribuyen a un desbalance en el contenido de Ca, Fe y K en raíces. Así, las dosis variables de La son responsables de la dosis-respuesta en forma de U de los contenidos nutrimentales. De manera semejante, Li et al. (2008) reportan también que la absorción de La3+ dependiente de su concentración ocurre de manera bifásica debido a la operación de dos sistemas de transporte separados: uno de alta afinidad, el cual es saturado a 10 µM de La y uno de baja afinidad que muestra una dependencia lineal sobre la concentración hasta al menos 100 µM de La.

Valores con letras distintas en cada columna son estadísticamente diferentes (LSD, P ≤ 0.05). DMS0.05 = diferencia mínima significativa al 5 %.
Cuadro 2. Efecto de la interacción entre tratamientos (concentración de La por fuente de La en la solución nutritiva) en la acumulación de K, Ca y La en hojas de tulipán cultivar Ille de France.
Así también, es evidente que la eficiencia en la aplicación de lantano, desde el punto de vista de absorción de nutrimentos, es dependiente del ión acompañante, es decir, el ión acompañante afecta la movilidad dentro de la planta, registrándose mayor acumulación de K, Ca y La cuando este último fue suministrado como LaCl3; como se discutió previamente.
Coeficiente de Correlación entre Suministro de La y Acumulación Nutrimental
El coeficiente de correlación entre el suministro de lantano y la acumulación de K, Ca y La en hojas se ajustó a modelos de función racional cuando el lantano fue adicionado como LaCl3 (Figura 2A), mientras que cuando se adicionó como La(NO3)36H2O se ajustó a modelos polinomiales de tercer grado (Figura 2B).

Figura 2. Curvas de acumulación de potasio (K), calcio (Ca) y lantano (La) en hojas de tulipán cv. Ille de France con LaCl3 (A) y La(NO3)36H2O (B). AC = acumulación; C = concentración en la solución nutritiva; r = coeficiente de correlación.
De estos hallazgos deriva una aplicación práctica, puesto que las ecuaciones que se generaron permiten estimar en campo el coeficiente de acumulación (mg o μg) de K, Ca y La presentes en hojas por planta de tulipán haciendo uso de datos reales.
Algunos cationes del grupo de los ETR se han utilizado ampliamente en estudios de canales de transporte de Ca en el endoplasma y como inhibidores de canales de Ca (Zeng et al., 2000). Los elementos del grupo de los lantánidos pueden ser transportados con un acarreador artificial a las células de las plantas a través de canales de Ca2+ (Zheng et al., 2000). A pesar de que la mayoría de estos cationes son trivalentes, se ha propuesto que existen mecanismos idénticos o muy similares implicados en la absorción de Ca y de elementos de las tierras raras (Tyler y Olsson, 2005). Estos estudios permiten establecer una relación antagónica entre el La y cationes esenciales en plantas. Sin embargo, las acumulaciones de K y Ca en hojas de tulipán halladas en esta investigación se correlacionaron de manera altamente significativa con la acumulación de lantano, cuando éste fue suministrado como LaCl3 o como La(NO3)36H2O. Dicha correlación fue más evidente a concentraciones de La menores a 20 µM para LaCl3 y menores a 30 µM para La(NO3)36H2O (Figura 2). Si bien los coeficientes de correlación entre suministro de La y acumulación de K, Ca y La son positivos con ambas fuentes de La estudiadas, los grados de correlación son distintos; por ejemplo, en el caso de acumulación de Ca en respuesta a concentraciones de La como La(NO3)36H2O se observó una correlación positiva casi perfecta (r = 0.99) que indica dependencia total entre ambas variables; por el contrario, el coeficiente de correlación entre la acumulación de La y la solución nutritiva La(NO3)36H2O, fue considerablemente menor (r = 0.70) en comparación con el resto.
En lo que respecta a coeficientes de acumulación, que permiten estimar la variabilidad en este caso de acumulación de La en hojas con respecto a la concentración suministrada en la solución nutritiva, existen divergencias en la literatura. Zhang y Shan (2001) reportaron que no existió una relación entre la acumulación de elementos de las tierras raras, entre ellos La, en vástagos de trigo y las fracciones extraíbles de éstos en el suelo. Por el contrario, en frijol no se observó relación positiva entre el contenido de La en el vástago y la concentración de éste en la solución nutritiva, y se encontró una alta acumulación de lantano en las raíces (von Toucher y Schmidhalter, 2005). Por otra parte, Xu et al. (2002), reportaron acumulaciones mayores de La en raíces que en vástago de maíz. Es importante considerar que los sistemas radicales de las especies antes descritas no son comparables con el del tulipán, especie que posee como órgano de reserva, multiplicación y anclaje un bulbo, y no existen reportes al respecto en especies bulbosas que puedan ser comparados con los resultados aquí reportados.
Los coeficientes de correlación suministro/bioacumulación de La fueron de 0.959 y 0.623 cuando se usó La(NO3)36H2O y LaCl3 respectivamente, como fuentes de este elemento. En suelo, los valores reportados para este coeficiente oscilan entre 0.6159 y 0.7410, cuando el lantano es aplicado en concentraciones de 20 mg kg-1, equivalentes aproximadamente a 144 μM de La (Hu et al., 2006), intervalo en el que se encuentra el coeficiente obtenido cuando se empleó La(NO3)36H2O, y el cual presenta valores inferiores de bioacumulación de La a los registrados con la otra fuente (Cuadro 1).
Los resultados positivos de ambas fuentes evaluadas a concentraciones menores a 20 μM de La sobre la acumulación de cationes esenciales como Ca y K, se relacionan de manera positiva con algunas variables de calidad comercial en esta especie. Ramírez et al. (2009), evaluaron el efecto de 0, 5, 10, 20, 30 y 40 μM de La suministrado como La(NO3)36H2O en la solución nutritiva y observaron que la adición de 10 μM de La incrementó significativamente el diámetro y longitud de la flor, en comparación con el tratamiento sin La. Adicionalmente no observó disminución de la longitud del tallo floral en las plantas tratadas con La, respecto al tratamiento testigo. Asimismo, reportan que no fueron observados efectos tóxicos del La en esta especie en hojas o flores.
Conclusiones
- Los resultados obtenidos indican que la especie Tulipa gesneriana L. responde diferencialmente al suministro de lantano (La) con distintas fuentes y en distintas concentraciones, y que este elemento de las tierras raras es traslocado a la parte aérea en esta especie.
- Concentraciones de La en la solución nutritiva entre 5 y 20 μM promueven la bioacumulación tanto de este elemento como de K y Ca, en particular cuando es abastecido como LaCl3. Estos hallazgos, aunado al hecho que no se observaron síntomas de deficiencias o de toxicidad en la planta, ni alteraciones en el crecimiento, permiten concluir que el La tiene efectos benéficos en el cultivo de tulipán.
- Desde el punto de vista práctico, el suministro de La para esta especie se puede considerar como una alternativa en la producción de tulipán, por su efecto sinérgico sobre la absorción de K y Ca, así como efectos positivos, reportados en otros estudios realizados por este grupo de trabajo, sobre parámetros de calidad de la flor de corte en tulipán como son diámetro y longitud de la flor.

