











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El nitrógeno es, después del agua, el nutriente más limitante para la productividad de las plantas de zonas áridas y semiáridas. En forma natural, el nitrógeno se encuentra en la atmósfera como gas, en un reservorio no disponible para las plantas, por lo que para poder utilizarlo, las plantas requieren establecer simbiosis con algunas especies de bacterias, lo que sucede por ejemplo en diversas especies de la familia de las leguminosas entre otras, lo que les permite aportar materia orgánica rica en nitrógeno al suelo de zonas áridas. La poca materia orgánica del suelo no puede ser utilizada por las plantas directamente, y para ello debe descomponerse y mineralizarse, procesos que son llevados a cabo principalmente por microorganismos del suelo, los cuales a su vez en ambientes áridos, no tienen siempre condiciones ideales para su actividad en cuanto a temperatura y humedad, por lo que los procesos de mineralización de nitrógeno en zonas áridas se dan por periodos breves de tiempo generalmente posteriores a pulsos de precipitación. En periodos secos es importante la fragmentación abiótica de la materia orgánica por el intemperismo, en especial por la radiación UV sobre compuestos como lignina, de difícil degradación. En la mineralización de nitrógeno en zonas áridas son de gran importancia las altas temperatura, la humedad variable e impredecible y las especies de plantas presentes, como determinantes de su gran heterogeneidad temporal y espacial. La utilización de los ecosistemas áridos por el hombre puede ocasionar perturbaciones que rompen el equilibrio en los procesos del suelo, su capacidad auto reguladora y el reciclaje de materia orgánica y nutrientes, pudiendo favorecer pérdidas de nitrógeno del sistema y de fertilidad del suelo, como es el caso de los desmontes y siembras de especies exóticas, el sobrepastoreo del ganado y el fuego.
Ciclo del Nitrógeno y Fijación Biológica
El nitrógeno es el elemento más limitante en los ecosistemas terrestres, de todos los nutrientes del suelo necesarios para el crecimiento de las plantas (Vitousek et al., 1997), en especial en ecosistemas áridos y semiáridos (Schlesinger et al., 1996; Chapin et al., 2002; Whitford, 2002). El nitrógeno inorgánico está constituido por las formas solubles como nitrito (NO2 -) y nitrato (NO3 -), amonio (NH4 +) intercambiable y no intercambiable fijado en las arcillas (Rutherford et al.,2007). El contenido de nitrógeno orgánico en el suelo incluye una gran variedad de formas (Binkley y Vitousek, 1989), puede no estar disponible para las plantas en moléculas complejas o perderse por denitrificación, erosión del suelo, lixiviado, volatilización (Philippot y Germon, 2005), y desempeña un papel importante en la composición y diversidad de las especies, y en la dinámica y funcionamiento de la mayoría de los ecosistemas. La mayoría de las plantas nativas de cada región están adaptadas y su funcionamiento óptimo va de acuerdo a las concentraciones más bajas de nitrógeno disponible en sus suelos (Vitousek et al., 1997).
La atmósfera es el reservorio más grande de nitrógeno con un 79% en su forma elemental (N2), sin embargo metabólicamente no está disponible para su asimilación por las plantas superiores que no poseen mecanismos para romper el triple enlace covalente (Philippot y Germon, 2005). El nitrógeno en forma gaseosa es la principal fuente primaria de entrada para los ecosistemas, existen organismos simbiontes y de vida libre con la capacidad de fijarlo de la atmósfera (Vitousek y Howarth, 1991). El proceso de fijación de nitrógeno es complejo y envuelve a una variedad de microorganismos, microfauna del suelo, plantas y animales, representa una entrada al ciclo terrestre del nitrógeno de gran importancia para ecosistemas áridos. Una de las familias de plantas más reconocidas como fijadoras de nitrógeno son las Leguminosas, tanto árboles como arbustos abundan en la mayoría de las regiones áridas y semiáridas, presentando una simbiosis con bacterias del género Rhizobium o Bradyrhizobium, formando nódulos donde la bacteria fija nitrógeno (Whitford, 2002).
Varios tipos de bacterias y algas azul-verde poseen la enzima nitrogenasa, que convierte el N2 atmosférico a amonio (NH4 +). Algunas de ellas como Rhizobium y actinomicetos del género Frankia existen en asociaciones simbióticas con plantas, pero otras como Azobacter ocurren en forma de vida libre en los suelos. En los ecosistemas desérticos son de gran importancia ambos procesos de fijación (Whitford, 2002), por simbiosis se beneficia la planta hospedera, posteriormente el nitrógeno entra al ecosistema cuando las hojas con alto contenido de nitrógeno se descomponen estimándose un aporte entre de 25 a 30 kg por hectárea al año en los mezquitales de California dentro del Desierto Sonorense (Rundel et al., 1982), mientras que en costras microbióticas del suelo formadas por líquenes, cianobacterias, algas verdes, musgos y hongos (Evans y Belnap, 1999) se ha estimado que aportan de 7 a 18 kg por hectárea por año en el desierto Sonorense en Arizona (Veluci et al., 2006). En costras de algas azul-verde también se reportan entre 1 y 5 kg de nitrógeno por hectárea por año (Schlesinger, 1997).
La fijación biológica de nitrógeno con microorganismos de vida libre y simbiótica con algunas plantas, representa una importante entrada de nitrógeno al suelo, lo cual cobra especial importancia en ecosistemas limitados por nitrógeno, como son las zonas áridas y semiáridas. La planta se beneficia con la asociación simbiótica y al morir, sus hojas o raíces aportan nitrógeno disponible tanto a microorganismos como a otras plantas.
Descomposición
La mayor parte del nitrógeno del suelo está contenido en la materia orgánica de los animales, plantas, hongos y bacterias muertos (Schlesinger, 1997), pero en el corto plazo no es disponible para las plantas y solo mediante los procesos de descomposición y posterior mineralización vuelve a estar disponible. Las principales formas biológicas de nitrógeno activas para las plantas son inorgánicas: amonio y nitrato (Vitousek y Matson, 1985).
Los procesos de descomposición y mineralización son llevados a cabo por una comunidad muy dinámica de fauna y microorganismos descomponedores (Huxman et al., 2004; Osler y Sommerkorn, 2007). La descomposición depende del clima, de la composición química de los restos vegetales y de los microorganismos del suelo. En zonas áridas y semiáridas los desdoblamientos microbianos de la materia orgánica de la superficie es limitada a breves periodos cuando el mantillo y el suelo están húmedos y las temperaturas del suelo son moderadas (Whitford, 2002). El inicio de la descomposición es la fragmentación que puede ser abiótica o biótica. La fragmentación y mineralización en ambientes mésicos ocurre de manera simultánea, mientras que en ecosistemas desérticos puede estar separada temporal y espacialmente. Viento, agua, calor y luz pueden interactuar con la descomposición de algunos compuestos de las plantas (Chapin et al., 2002).
El clima mediante la temperatura y la humedad, es el principal predictor global de la descomposición, la evapotranspiración actual es un índice del efecto del clima en la descomposición, dentro de una región climática el contenido químico como la relación lignina - nitrógeno de la hojarasca, pueden predecir su descomposición, principalmente en regiones tropicales y mediterráneas (Aerts, 1997).
La radiación UV es importante en zonas áridas ya que degrada ciertos compuestos por oxidación fotoquímica, la cual parece ser más importante durante los períodos secos (Gallo et al., 2009), y otros procesos descomponen la materia orgánica en periodos lluviosos. En Arizona dentro del desierto Sonorense se evaluó el efecto de la radiación UV en la pérdida de peso de la hojarasca de Larrea tridentata encontrando que la lignina fue el elemento más afectado por dicha radiación, seguida de los lípidos, estimándose que entre el 14 al 22% de la pérdida de peso podría ser atribuible a la radiación UV en un periodo de cinco meses de exposición (Day et al., 2007).
La descomposición continúa con la despolimerización de compuestos orgánicos nitrogenados, inicialmente no son biodisponibles por ser demasiado grandes (Chapin et al., 2002), sin embargo son separados por enzimas extracelulares a monómeros como aminoácidos, amino-azúcares, ácidos nucleicos, que se denominan nitrógeno orgánico disuelto y está disponible para los microorganismos, hongos micorrícicos (Govindarajulu et al., 2005) y plantas en ecosistemas limitados en nitrógeno, aunque aún no está bien clara la proporción adquirida por las plantas (Hodge et al., 2000).
La descomposición de la materia orgánica permite reciclar nutrientes no disponibles para las plantas y volverlos a hacer disponibles, en mayor proporción los microorganismos del suelo son los responsables de esta tarea.
Microorganismos del Suelo
En el suelo existen una gran variedad de microorganismos como bacterias y protozoarios, así como nemátodos, ácaros y hongos, que intervienen en los procesos de transformación de la materia orgánica del suelo. Adicionalmente con la depredación entre ellos, por ejemplo los nemátodos que se alimentan de bacterias y protistas, y microártropodos que se alimentan de hongos y lombrices juveniles (Osler y Sommerkorn, 2007), liberan nutrientes que son reutilizados en descomposición y mineralización. Estos microorganismos son de gran importancia actuando como descomponedores y como reservorio de nitrógeno, que eventualmente se libera con su muerte o depredación.
La temperatura y la humedad óptimas para el crecimiento bacteriano no siempre se presentan al mismo tiempo en los ecosistemas áridos y semiáridos por lo que las tasas de descomposición y mineralización de la materia orgánica se llevan a cabo principalmente en periodos cortos de tiempo e irregulares en el año, durante los pulsos de humedad. Los microorganismos responden rápidamente a la disponibilidad de humedad en el suelo, aún cuando solo sea un evento pequeño que solo humedezca la parte superficial, esto lleva a que el nitrógeno disponible se acumule en la época seca cuando las plantas no pueden utilizarlo (Austin et al., 2004). Los microbios presentes en suelos semiáridos están adaptados a utilizar incrementos pequeños de humedad (Huxman et al., 2004). Durante períodos de sequía, los microorganismos del suelo incorporan solutos para ajustarse osmóticamente a la ausencia de agua en el suelo, inmovilizando nutrientes disponibles en el medio. La capacidad de aclimatación a los pulsos de agua y pulsos de humedad (sequedad puede variar entre microorganismos), siendo mayor para bacterias gram positivas (Schimel et al., 2007).
Cuando los periodos de secado son muy prolongados, al humedecerse se presenta un estrés fisiológico para las comunidades microbianas superficiales, y pueden inducir la ruptura de la pared celular y muerte de biomasa microbiana. Al rehumedecer, se produce un choque osmótico microbiano que induce la lisis de las células con liberación de solutos intracelulares, estos sustratos lábiles de carbono y nitrógeno pueden ser mineralizados rápidamente por los microorganismos sobrevivientes (Fierer et al., 2003). Por otra parte, los pulsos de secado y humedecimiento del suelo pueden causar la ruptura de agregados, permitiendo a los descomponedores de la materia orgánica acceder a ella (Lundquist et al., 1999). En el Desierto Chihuahuense se encontró que con los periodos de sequía se aumentaba la disponibilidad de nitrógeno, sugiriendo que diversos procesos son afectados resultando en dicho aumento de nitrógeno (Fisher et al., 1987).
Los microorganismos del suelo son de gran importancia al transformar compuestos orgánicos a inorgánicos disponibles para las plantas, con su actividad de descomposición y mineralización de nitrógeno. También cobra importancia en zonas áridas y semiáridas la heterogeneidad de humedad y temperatura presente bajo el dosel de árboles y arbustos en comparación con los espacios abiertos, que permite a los microorganismos bajo el dosel tener más actividad a lo largo del año (Whitford, 2002).
Mineralización de Nitrógeno y su Medición
Se denomina mineralización al proceso mediante el cual el nitrógeno orgánico del suelo es transformado por los microorganismos del suelo a formas inorgánicas (amonio y nitrato; Binkley y Hart, 1989). El primer producto de la mineralización es el amoniaco (NH3) el cual puede adquirir un hidrógeno y formar amonio (NH4 +), que puede ser fijado por las arcillas del suelo o por la materia orgánica, volatilizado como amoniaco, asimilado por plantas o microorganismos (Philippot y Germon, 2005), lixiviado u oxidado por bacterias autotróficas mediante el proceso de nitrificación, en donde pierde dos átomos de hidrógeno para formar nitrito inorgánicas son tomadas por los microorganismos incorporándolas a su biomasa y es llamada inmovilización. El nitrato puede pasar al proceso de denitrificación donde se convierte a óxido nitroso (N2O) y nitrógeno elemental (N2; Binkley y Vitousek, 1989).
La cantidad total de nitrógeno liberada de la materia orgánica es llamada mineralización gruesa o bruta, mientras que la cantidad resultante después de eliminar la inmovilización microbiana se denomina mineralización neta (Binkley y Vitousek, 1989) y se expresa como la tasa de cambio en un periodo de tiempo. La mineralización neta se ha utilizado para estimar el nitrógeno disponible para la planta (Hart et al., 1994; Neill et al., 1999), por lo que se han desarrollado formas in situ y de laboratorio para determinar dicha tasa. Algunas dificultades se presentan en su medición debido a los procesos de denitrificación, volatilización, inmovilización, adquisición por la planta y lixiviado (Campbell et al., 1995).
En varios ecosistemas la tasa neta de mineralización de nitrógeno y la cantidad total de nitrógeno son indicadores de fertilidad del suelo (Nadelhoffer et al., 1983; Vitousek y Matson, 1985). El nitrógeno total puede usarse para clasificar sitios de acuerdo a la disponibilidad de nitrógeno en grandes extensiones, sin embargo para sitios y hábitats específicos no lo es, así como donde la perturbación del suelo es importante (Binkley y Vitousek, 1989).
Los métodos para evaluar la mineralización son un indicador de la cantidad de nitrógeno orgánico que pasa a inorgánico en un periodo de tiempo, para lo cual se realizan incubaciones de muestras de suelo que permiten calcular la tasa de amonio y nitrato que se mineraliza después de la incubación (Robertson y Vitousek, 1981). Existen técnicas de laboratorio con incubaciones anaeróbicas y aeróbicas, en las primeras a pesar que no representan condiciones normales en el suelo se utilizan para estimar la toma de nutrientes en cultivos agrícolas. Las incubaciones aeróbicas consisten en incubaciones de 10 a 30 días, utilizando de 5 a 20 g de muestra a temperatura de 20 o 25 °C (Binkley y Vitousek, 1989) donde se obtiene la mineralización neta potencial.
También se incuba directamente en el sitio obteniendo la tasa neta de mineralización in situ, y se usan tres métodos principalmente: las bolsas enterradas, cilindros y trampas de resinas (Westermann y Crothers, 1980; Raison et al., 1987; Eghball, 2000). El primer método utiliza bolsas de polietileno, se llenan con suelo y sellan, durante varios días permanecen en el sitio de muestreo bajo las mismas condiciones de temperatura del suelo. Es importante considerar que las bolsas pueden presentar daño por insectos o raíces de las plantas, pudiendo contribuir a pérdidas de mineralización de nitrógeno (Eno, 1960), mientras que concentraciones elevadas de nitratos y dióxido de carbono en las bolsas pueden promover la denitrificación (Hanselman et al., 2004). Destaca la característica impermeable de la bolsa por lo que la humedad es constante, sin tener las fluctuaciones en humedad del suelo, y si la bolsa conserva su integridad durante el periodo de incubación no existirá lixiviado de nutrientes. La mineralización neta de nitrógeno por el método de bolsas enterradas es la mineralización que excede a la inmovilización microbiana, y al estar la muestra de suelo en la bolsa se evita la toma por la planta (Schlesinger, 1997).
El método de cilindros fue desarrollado como alternativa más durable a las bolsas de plástico, se utilizan cilindros de PVC o metal para excluir la lluvia, existen metodologías con la parte superior cerrada o abierta. Los cilindros presentan abertura en la parte inferior, lo que le confiere mayor aireación que la bolsa de plástico, inclusive se le adicionan perforaciones laterales para promover intercambio de aire con el suelo a sus lados, sin embargo parte del nitrógeno mineralizado puede perderse a través de los orificios de aireación, o existe también posibilidad de entrada de raíces en el cilindro, absorber nitrógeno y afectar el resultado de mineralización (Hanselman et al., 2004). En este método también se perturba el área, matando las raíces y evitando la absorción de ellas que normalmente ocurriría, por lo cual los microorganismos contaran con mayor humedad que la que tiene en el suelo.
El tercer método más citado son las bolsas con resinas de intercambio iónico, consiste en el uso de bolsas de nylon adicionadas con resinas. La cantidad de resina adicionada debe tener suficiente capacidad de intercambio para retener todos los iones de amonio y nitrato que alcancen la bolsa, pero los resultados son difíciles de interpretar en términos de procesos porque la captura de iones por la bolsa de resinas es afectada por la tasa de mineralización, la tasa de respuesta y la movilidad de los iones en el suelo (Binkley, 1984). Al igual que en los métodos anteriores se excluye a la planta de la toma de agua y nutrientes. Existen combinaciones de los métodos por ejemplo el uso de cilindros con resinas en los extremos.
En el Cuadro 1 se pueden apreciar algunas características de métodos para medir la mineralización neta in situ donde diferentes autores han utilizado bolsas de plástico, cilindros (cerrados, abiertos y con resinas), latas y bolsas con resinas de intercambio iónico; enterrando estos recipientes a diferentes profundidades en el sitio de muestreo, generalmente en la parte superficial del suelo, de 2 a 15 cm de profundidad. Es importante considerar la profundidad de muestreo porque en la parte superficial es donde se encuentra mayor cantidad de materia orgánica para descomponer y mineralizar.
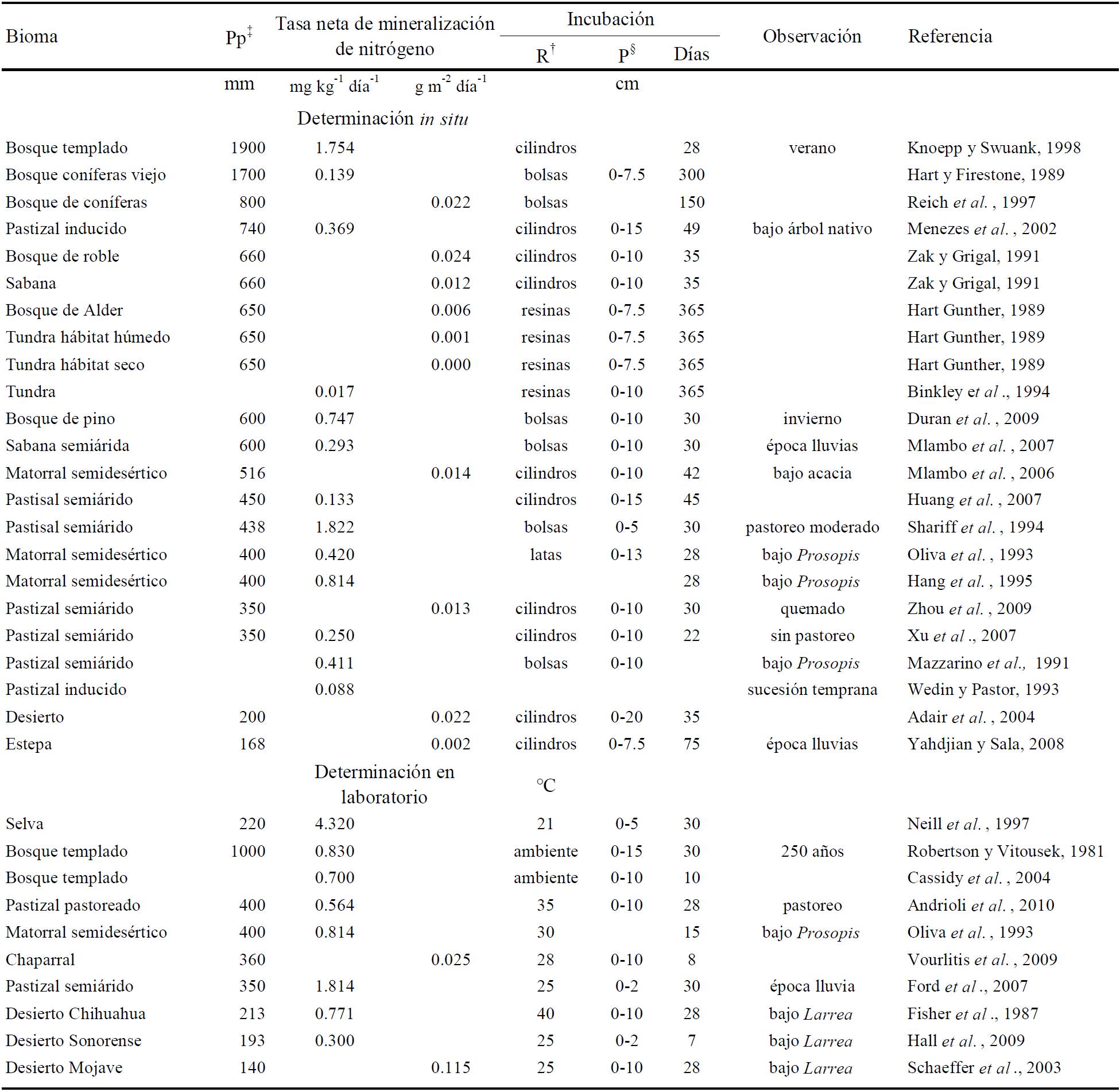
‡ Pp = precipitación pluvial; † R = recipiente utilizado para medir la mineralización neta in situ; § P = profundidad.
Cuadro 1 Valores máximos de tasa de mineralización neta en diferentes biomas utilizando metodologías in situ y de laboratorio.
El periodo de incubación más usado en la recopilación del Cuadro 1, es cercano a 30 días, sin embargo hay estudios con incubaciones de hasta 365 días, generalmente en lugares fríos. Se reporta la cantidad de amonio y nitrato incubado restando el contenido inicial, por peso (gramos de suelo) o superficie de suelo seco (metros cuadrados y por hectárea) en un determinado periodo.
Las determinaciones de laboratorio a diferencia de las in situ, se llevan a cabo a una temperatura ambiente o constante de 21 a 40 °C y con incubaciones de 7 a 30 días (Cuadro 1). Las incubaciones en laboratorio nos muestran el potencial máximo de mineralización al contar con humedad adicional y temperatura constante. Las determinaciones in situ buscan que las condiciones de incubación sean más cercanas a las que tiene el suelo en fluctuación de temperatura, humedad y aireación.
Los valores máximos de tasa de mineralización mostrada en el Cuadro 1, tanto in situ como potencial se presentan en los lugares con mayor precipitación, mientras que en regiones áridas y semiáridas los resultados máximos fueron obtenidos en el suelo bajo el dosel de arbustos o árboles (principalmente de la familia de las leguminosas); otro elemento importante es el periodo de análisis de la mineralización, en algunos casos se restringe al periodo con mayor actividad microbiana; por lo que la comparación entre biomas debe de considerar la heterogeneidad espacial y temporal de las zonas áridas y semiáridas.
Ningún método provee una estimación exacta de la disponibilidad de nitrógeno, las técnicas de laboratorio no pueden considerar las variaciones en las condiciones de campo entre los sitios de muestreo. Cualquier método debe ser considerado como un indicador de disponibilidad en vez de la disponibilidad actual real (Binkley y Vitousek, 1989).
Condiciones que Afectan la Mineralización en Zonas Áridas
La mineralización de nitrógeno es afectada por factores ambientales físicos como temperatura, humedad y aireación, por el contenido de nitrógeno del sustrato, tipo de compuestos orgánicos y por los microorganismos del suelo (Binkley y Hart, 1989).
En zonas áridas la actividad microbiana es frecuentemente limitada por la humedad (Austin et al., 2004), por lo que la tasas de mineralización e inmovilización de nitrógeno están ligadas a los pulsos de precipitación (Fisher et al., 1987) y entrada de materia orgánica, las sequías periódicas resultan en biomasa muerta de microorganismos que sirve de reservorio (Whitford, 2002) y dejan disponibles nutrientes para las plantas u otros microorganismos, por lo que la inmovilización microbiana puede reducir pérdidas de nitrógeno del ecosistema.
Como resultado de las condiciones ambientales en la tundra casi solo se encuentra amonio mientras que en los desiertos el nitrato es más importante como elemento presente disponible para las plantas (Schlesinger, 1997). En el Cuadro 1 el valor más bajo de mineralización corresponde a este bioma. Se menciona que en las zonas áridas y semiáridas donde se presentan los niveles más bajos de materia orgánica del suelo (Stevenson, 1994), debido a la densidad baja de plantas que condiciona su aporte de materia orgánica al suelo.
También la mineralización de nitrógeno en zonas áridas es afectada por la disponibilidad de carbono y nitrógeno (Schaeffer et al., 2003), la calidad de la materia orgánica es tan importante como la relación carbono/nitrógeno, y la concentración de lignina regula la cantidad y velocidad de la mineralización de nitrógeno (Lajtha y Schlesinger, 1986). En el desierto de Mojave Rundel y Gibson (1996) estimaron la tasa de mineralización en 6.5 kg de nitrógeno ha-1 año-1.
La tasa de mineralización no siempre cambia en gradientes de humedad, debido a que la productividad primaria neta, la materia orgánica del suelo, la biomasa microbiana y la inmovilización de nitrógeno del suelo también cambian con la precipitación (Epstein et al., 2006) por lo cual es importante considerar el nitrógeno total y disponible del suelo para comparar entre gradientes.
Tipos Funcionales y Nitrógeno
Los tipos funcionales de plantas se pueden definir como conjuntos de especies que muestran respuestas similares al ambiente que las rodea y efectos similares sobre el funcionamiento del ecosistema (Díaz y Cabido, 1997). Los tipos funcionales pueden dar una idea de los procesos que ocurren en los ecosistemas, y su cuantificación nos define la diversidad funcional. La diversidad funcional afecta procesos como la productividad primaria neta aérea, ciclos del agua y nutrientes, formación y retención de suelo, etc. (Díaz et al., 2006). Adicionalmente las características de los tipos funcionales afectan el suministro de nutrientes a través de sus efectos en exudados de las raíces, microambiente y calidad de la hojarasca. Los exudados de las raíces son compuestos que pueden propiciar o reducir la mineralización. Con la sucesión a través del tiempo la vegetación modifica la temperatura, la humedad del suelo y la calidad y cantidad de hojarasca que entra al sistema (Lambers et al., 1998), es por eso que la presencia de un árbol o arbusto en los desiertos favorece la acumulación de nutrientes bajo su dosel.
Respecto al ciclo del nitrógeno las plantas con características que permiten la fijación microbiana de nitrógeno atmosférico impactan el suelo (Canadell et al., 2007), favoreciendo mayor contenido de nitrógeno en hojarasca y posteriormente su mineralizado en el suelo, por lo que es de gran importancia para el nitrógeno del suelo de zonas áridas la presencia de tipos funcionales que fijan nitrógeno.
Estructura de la Vegetación
Islas de fertilidad. Las islas de fertilidad en zonas áridas y semiáridas se forman bajo el dosel de árboles y arbustos, con suelos que contienen mayor humedad, nutrientes, incluyendo el nitrógeno disponible (García-Moya y McKell, 1970; Stubbs y Pyke, 2005) y microclima, en comparación con los espacios abiertos o interespacios. Varios mecanismos se han propuesto para explicar la formación de islas de fertilidad en las comunidades del desierto, como la acumulación de residuos en las plantas, favoreciendo las comunidades microbianas (Garner y Steinberger, 1989), la fijación de nitrógeno por simbiosis en la raíz por el aumento del nitrógeno en las islas de fertilidad que se desarrollan debajo de leguminosas y el acceso a agua subterránea por plantas freatofitas (Whitford, 2002). En varios desiertos del mundo se han documentado islas de fertilidad bajo el dosel de árboles o arbustos (Cuadro 2) con mayor contenido de nitrógeno: en el Sonorense (Schlesinger et al,. 1996), de Mojave (Ewing et al., 2007), Chihuahuense (Kieft et al., 1998), así como en Argentina en la región del Chaco (Oliva et al., 1993) y en África en el Kalahari (Wang et al., 2009). La distribución de plantas en zonas áridas no es uniforme (Bolton et al., 1990), existe vegetación perenne que propicia un microclima y anuales que solo prosperan en periodos favorables para su crecimiento, como es el caso en el desierto de Mojave donde se ha estudiado cómo el arbusto Larrea tridentata favorece islas de fertilidad y acumula bajo su dosel mayor concentración de nutrientes, capacidad para retener agua y actividad microbiana. Las comunidades microbianas son más grandes, diversas y reciclan nitrógeno más eficientemente que los microorganismos de los espacios abiertos (Ewing et al., 2007).
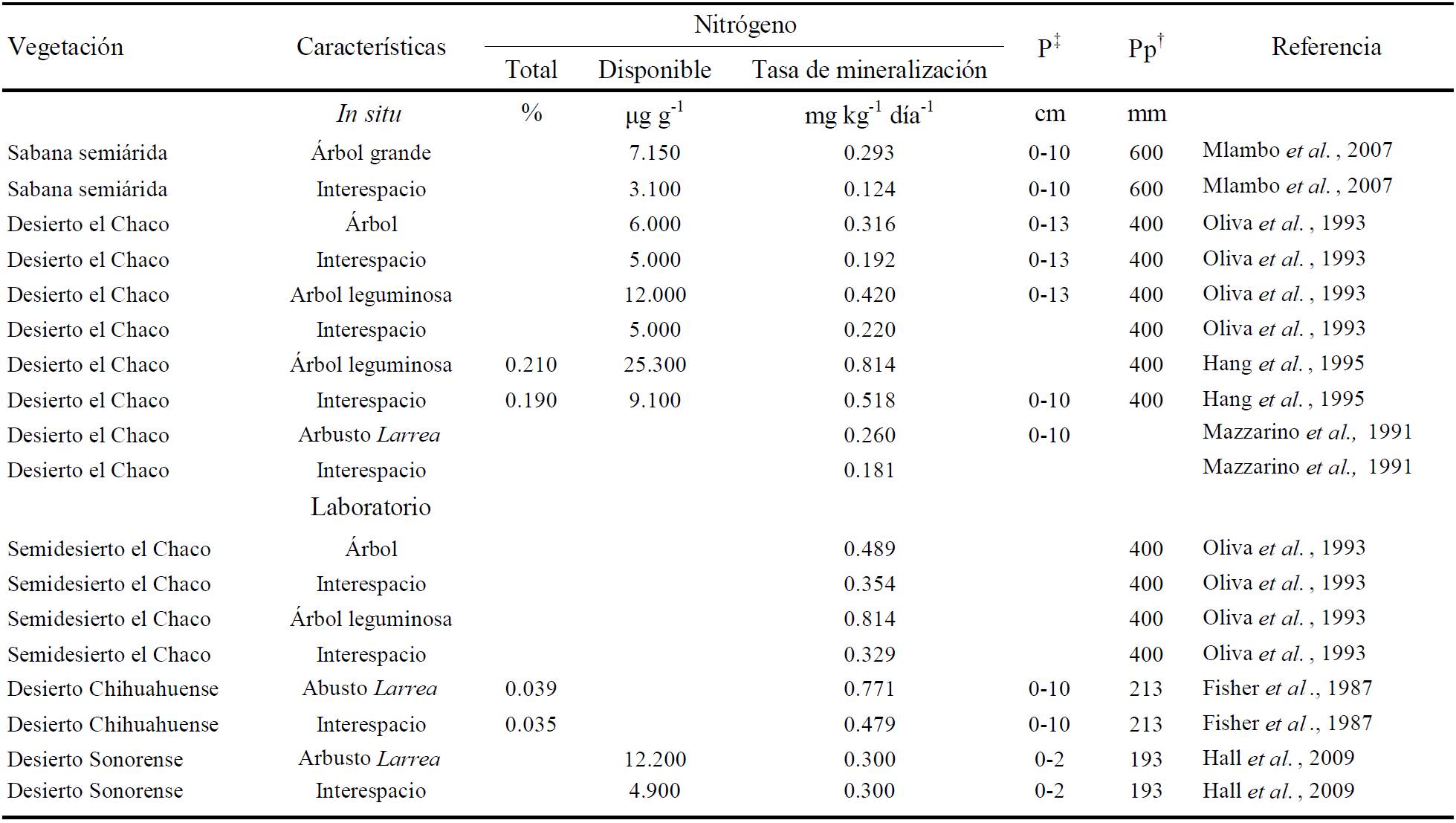
‡ P = profundidad; † Pp = precipitación pluvial.
Cuadro 2 Nitrógeno total, disponible y mineralización neta bajo el dosel e interespacio.
Al norte del desierto Chihuahuense donde han ocurrido cambios de pastizal a matorral desértico de Larrea, los nutrientes se han redistribuido en islas de fertilidad. Comparando el contenido de nutrientes bajo pasto y bajo Larrea, en estos dos tipos de vegetación, Larrea concentra mayor cantidad de nutrientes que bajo los pastos, sin embargo como el pasto tiene una mayor cobertura, al convertir a área los recursos disponibles son mayores en el pastizal (Kieft et al., 1998).
La presencia de leguminosas leñosas en el desierto Sonorense, favorece la formación de suelos más fértiles bajo su dosel al incrementar la cantidad de nitrógeno. En la parte de California de este desierto se encontraron cantidades muy elevadas de nitrato bajo el dosel de mezquite por la disminución de procesos de lixiviación y denitrificación debido a la aridez (Virginia y Jarrell, 1983). En la parte central de Arizona se encontró también que bajo el dosel de Prosopis velutina existe mayor contenido de materia orgánica y mineralización neta de nitrógeno (Schade y Hobbie, 2005).
Las islas de fertilidad en ecosistemas desérticos también son favorecidas por el movimiento de agua a través de procesos físicos y biológicos (Schlesinger y Pilmanis, 1998). El acceso a aguas subterráneas de plantas freatofitas como Prosopis velutina lleva a cambios en los procesos biogeoquímicos del suelo (Schade y Hobbie, 2005). La productividad de plantas que tiene acceso a aguas subterráneas es mucho mayor que la que se podría obtener con los niveles de precipitación de algunos desiertos, en los mezquitales de California, dentro del desierto Sonorense la precipitación promedio anual es de 70 mm, sin embargo la biomasa aérea de los mezquitales de esa región es de 13 000 kg por hectárea con una productividad anual de 3 700 kg por hectárea. Esa gran productividad es posible gracias al acceso a aguas subterráneas, a la simbiosis con microorganismos fijadores de nitrógeno y la dinámica de los procesos de mineralización de nitrógeno (Rundel et al., 1982).
Fuera del dosel de las islas de fertilidad, los procesos de mineralización son contrastantes y propician la heterogeneidad biológica y funcional en los ecosistemas áridos y semiáridos (Cuadro 2). Existe un patrón recurrente en esos ecosistemas, dado que en un número significativo de estudios se han determinado mayores tasas de mineralización neta de nitrógeno bajo árboles y arbustos, en particular de leguminosas, que las encontradas en el interespacio cercano desprovisto de vegetación leñosa (Cuadro 2). La heterogeneidad del paisaje ocasionado por las islas de fertilidad, resulta en diferencias de nutrientes del suelo, microclima y microorganismos en las zonas áridas, y ocasiona que a lo largo del paisaje la mineralización de nitrógeno sea marcadamente diferente. La heterogeneidad espacial es una característica importante y clave para entender el funcionamiento de los ecosistemas áridos y semiáridos, en particular los procesos de dinámica y mineralización neta del nitrógeno.
En la mayoría de los trabajos reportados en el Cuadro 2, además de mostrar mayor tasa de mineralización bajo árboles ó arbustos, también presentan valores mayores de nitrógeno disponible (la suma de amonio más nitrato) y nitrógeno total. Algunos estudios incluso, han encontrado mayor nitrógeno disponible bajo el dosel de Larrea (Hall et al., 2009) aunque la tasa de mineralización potencial para Larrea e interespacio fue similar. Tanto los datos de evaluaciones in situ como potenciales mostraron las mismas tendencias. Lo anterior confirma la importancia determinante de la especie o tipo funcional presente, en el aporte de nitrógeno al suelo y mineralización de nitrógeno en los ecosistemas de zonas áridas y semiáridas. Aún los interespacios cercanos a una leguminosa mostraron más nitrógeno en el suelo que los interespacios cercanos a árboles no leguminosas (Oliva et al., 1993). Una misma tendencia se ha encontrado previo a la lluvia y posterior a ella (Hang et al., 1995).
El Cuadro 2 muestra evidencias de que las especies de plantas presentes influencian el contenido de nitrógeno del suelo, y que la presencia de ciertos tipos de cubierta vegetal en ecosistemas de zonas áridas, favorece el contenido de nitrógeno total, disponible y tasa de mineralización de nitrógeno, de acuerdo a las características funcionales de las especies existentes.
Perturbación de la cubierta vegetal. En el Cuadro 3 se muestra la tasa neta de mineralización en vegetación natural y perturbada de zonas áridas y semiáridas de varios autores. En forma general la vegetación natural presenta mayores tasas de mineralización neta de nitrógeno, con las excepciones de trabajos donde la perturbación era reciente y quizás el efecto sobre los procesos de mineralización aún no se mostraba (Oliva et al., 1993), y donde el área desmontada presentaba mayor tasa de mineralización en un año lluvioso (Hang et al., 1995), sin embargo en un año con lluvias por debajo del promedio la vegetación natural presentó mayor tasa de mineralización.
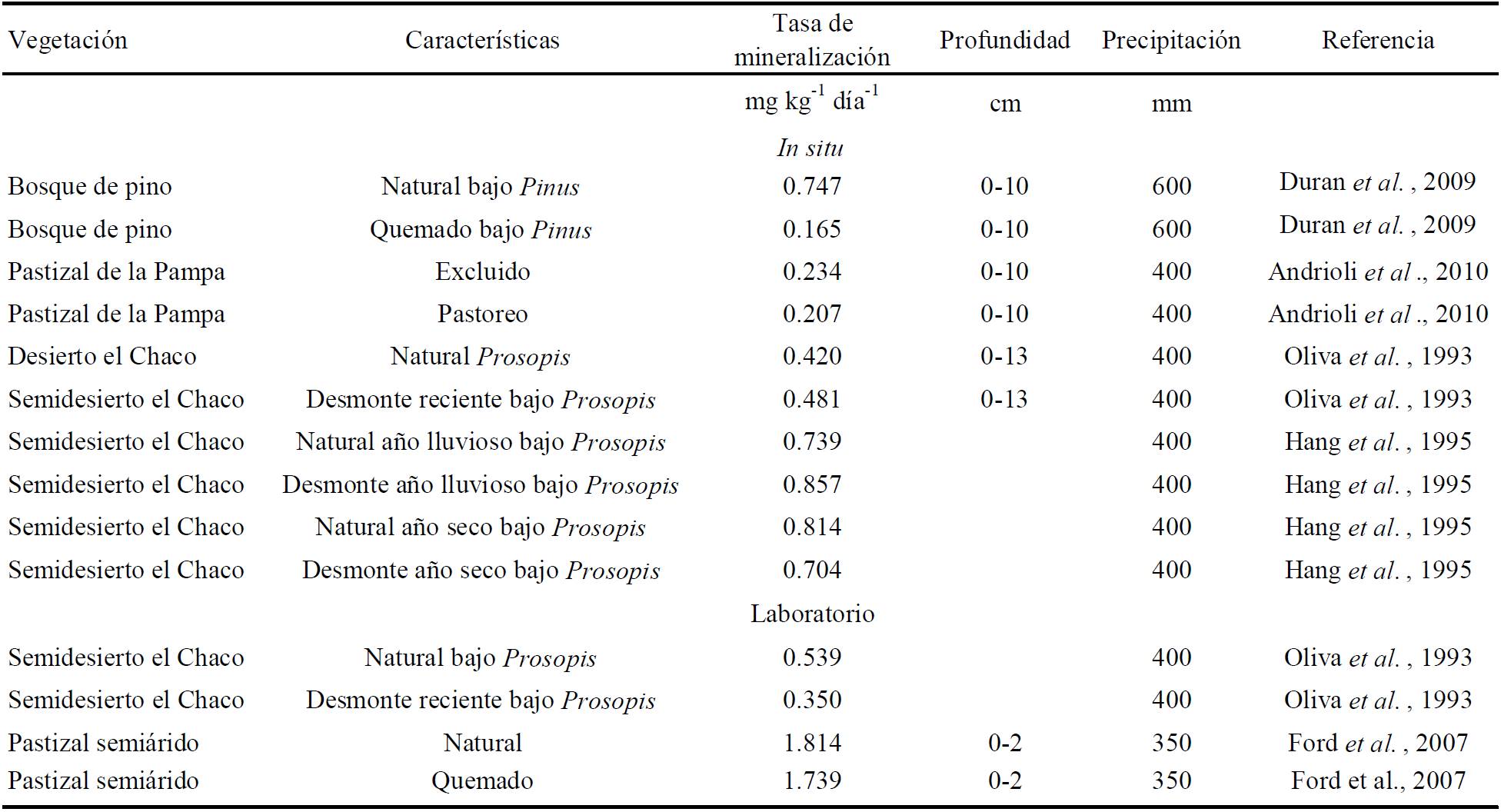
En cuanto a la mineralización neta potencial se presenta la misma tendencia de mayor mineralización en el suelo de la vegetación natural, aunque la diferencia no es tan grande, porque en esta técnica se igualan las condiciones de humedad y temperatura; lo cual nos explica la importancia del microclima, es decir, humedad y temperatura que favorece la vegetación presente y que se ve modificada con la perturbación.
La eliminación de la cubierta vegetal por perturbaciones como: pastoreo excesivo del ganado, desmonte con maquinaria o por fuego, afecta los procesos y el funcionamiento del ecosistema, resultando en modificaciones que pueden llevar a disminuir el contenido de nitrógeno del suelo (Cuadro 3) con el paso del tiempo por pérdidas de nitrógeno del ecosistema.
Aportes de la Hoja y Reabsorción de Nutrientes
La importancia de la caída de las hojas y renovación de raíces en el ciclo de los nutrientes consiste en que retorna materia orgánica y elementos minerales, que constituye el principal proceso de transferencia de nutrientes al suelo y puede llegar a ser de más de 90% del nitrógeno absorbido por las plantas (Chapin et al., 2002). Los nutrientes que pasan al suelo desde la planta pueden reducirse por el proceso de reabsorción o retranslocación de nutrientes, que consiste en la hidrólisis de los nutrientes contenidos en la hoja senescente, y sucede cuando la célula se desintegra junto con el cloroplasto donde se contiene más de 70% de la proteína de la hoja. Metabólicamente la fotosíntesis es reemplazada por el catabolismo de cloroplastos y macromoléculas, con el posterior transporte de los productos resultantes a tejidos en crecimiento o hacia zonas perennes de la planta donde son almacenados hasta su posterior utilización (Gan y Amasino, 1997). Las plantas reabsorben en promedio la mitad de su contenido foliar de nitrógeno antes de la caída de la hoja (Aerts, 1997), aunque la eficiencia en la reabsorción es muy variable.
En diferentes regiones climáticas la descomposición de la hojarasca de árboles y arbustos siempre verdes es más lenta que de los deciduos, esto se debe a la baja concentración de nutrientes en los perennifolios y a la presencia de lignina y otros compuestos secundarios (Wedin, 1999; Aerts y Chapin, 2000).
La producción de hojarasca en la parte central del Desierto Sonorense presenta gran variación espacial, debido a los cambios en composición y estructura de la vegetación, siendo los arroyos los lugares más productivos, acumulando 357 g m-2 año-1, mientras que las planicies solo 60 g m-2 año-1 y los matorrales de ladera 157 g m-2 año-1 (Martínez-Yrízar et al., 1999), presentándose esa producción de hojarasca en otoño y primavera, mostrando la heterogeneidad tanto temporal como espacial del aporte de hojarasca dentro de una misma región.
Pérdidas de Nitrógeno del Ecosistema
En suelos de zonas áridas las pérdidas hacia la atmósfera por erosión, volatilización, nitrificación y denitrificación son altas (Peterjohn y Schlesinger, 1990). La volatilización de amonio ocurre en suelos con pH de 7 o mayores, y como la mayoría de los suelos de los desiertos son básicos, existe un gran potencial de volatilización de amonio en sus suelos (Whitford, 2002), aunque es controlada por la tasa de amonificación (Schlesinger y Peterjohn, 1991). La denitrificación es un proceso anaeróbico que lo llevan a cabo bacterias principalmente, algunos hongos y levaduras (Van Spanning et al., 2005), consiste en la reducción microbiana de nitrato o nitrito a óxido nitroso (N2O) o nitrógeno elemental. Este proceso es afectado por la inmovilización microbiana. Al humedecerse el suelo y en presencia de carbono los microorganismos incorporan nitrógeno a su biomasa, sin embargo en suelos bajos en nutrientes se presentan altas tasas de denitrificación similares a las de suelos de regiones templadas y tropicales posiblemente debido a los pulsos de secado-humedecimiento de los desiertos (Peterjohn y Schlesinger, 1991), por lo cual cobra especial importancia la presencia de plantas que aporten carbono al suelo y favorezcan la conservación de nitrógeno en el suelo por inmovilización en biomasa microbiana. La inmovilización microbiana de nitrógeno es un proceso importante documentado en el suelo del Desierto Chihuahuense (Fisher et al., 1987).
Otras pérdidas importantes son por lixiviación. En el subsuelo de desiertos cálidos y matorrales áridos es donde se ha acumulado más nitrógeno biodisponible, aparentemente por lixiviación, a través de miles de años (Walvoord et al., 2003).
Conclusiones
- La mineralización neta de nitrógeno es un indicador de la disponibilidad de este nutriente, sin ser la disponibilidad real actual. En su interpretación se debe tomar en cuenta la metodología utilizada al comparar los resultados de diferentes autores, así como el nitrógeno disponible y total del suelo. La mineralización neta potencial nos muestra el valor máximo al eliminar la limitante de humedad y temperatura pero bajo condiciones artificiales, mientras que las determinaciones in situ siguen de manera más cercana las condiciones de campo.
- Los tipos funcionales de plantas con capacidad de establecer fijación por simbiosis con bacterias son una importante fuente de aporte de nitrógeno orgánico al suelo de zonas áridas y semiáridas, por ser ecosistemas limitados por nitrógeno. La materia orgánica del suelo es trasformada por los microorganismos con su actividad de descomposición y mineralización, vuelven a hacer disponible nitrógeno para las plantas, además son un reservorio de nutrientes en su biomasa, que inmoviliza elementos y los conserva en el suelo evitando que pasen a procesos de pérdida de nitrógeno del ecosistema.
- La mineralización de nitrógeno en zonas áridas y semiáridas se da en parches de manera heterogénea, condicionada a condiciones de microclima y aporte de materia orgánica por las plantas presentes en las islas de fertilidad por lo que presenta valores mayores bajo el dosel de árboles y arbustos, especialmente leguminosas, comparado con los espacios abiertos adyacentes.
- La perturbación de la cubierta vegetal mediante la eliminación de las islas de fertilidad, afecta muy fuertemente los procesos y el funcionamiento del ecosistema, en particular la dinámica del nitrógeno, resultando en modificaciones que pueden llevar a disminuir su contenido en el suelo, y con el paso del tiempo acentuar las pérdidas de nitrógeno del ecosistema.

