











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La papa es un cultivo importante en la dieta alimenticia de países densamente poblados como China que cultiva 1 000 000 de hectáreas con rendimientos promedio de 13.6 Mg ha-1y México que cultiva 72 000 hectáreas con un rendimiento medio en condiciones de riego de 30 Mg ha-1 y su rendimiento promedio mundial es de 15 Mg ha-1 (FAO, 2008). Las variedades comerciales están adaptadas a fotoperiodos cortos, por ello y por requerimientos de sanidad, se siembran en zonas áridas o semiáridas, cuyos suelos están afectados en mayor o menor grado por procesos de salinización natural o inducida por el hombre a través del riego con aguas de concentración salina superior a 2 g L-1 (Ayers y Westcot, 1987). La salinidad afecta el crecimiento y desarrollo de las plantas de papa desde sus primeros estadios. Moorby y Milthorpe (1983) señalan que la inhibición de la etapa de emergencia de brotes por estrés salino provoca alteraciones metabólicas y fisiológicas en las etapas vegetativas y de llenado del tubérculo, que inciden en la disminución del rendimiento por unidad de superficie cultivada y en la reducción espacial de la frontera del cultivo. Bernstein (1964) clasificó a la papa como una hortaliza moderadamente sensible a salinidad, ya que a concentraciones de 1.92 a 3.84 g L-1 en soluciones de NaCl, redujo en forma notable su rendimiento. Levy (1992) evaluó la tolerancia de catorce cultivares de papa en soluciones de CaCl2 y NaCl, con concentraciones de 3.90 a 4.41 g L-1 y reportó un retraso en la emergencia de plántulas, la formación de tubérculos pequeños y que las variedades tempranas toleraron más la salinidad que las tardías. Martínez et al. (1996) señalan que cv. Alpha fue sensible a NaCl. Sánchez et al. (2003) y Sánchez et al. (2008) investigaron la emergencia y acumulación de biomasa de brotes de tubérculos y de plantas de papa cv. Alpha, respectivamente, en diversas condiciones salinas a concentraciones de 0 a 9.60 g L-1, y concluyeron que los brotes y las plantas fueron sensibles a NaHCO3 y MgSO47H2O, ya que su máxima acumulación se dio a una concentración de 1.13 g L-1, moderadamente sensibles a NaCl y MgCl26H2O (su máxima acumulación de biomasa se registró con 2.48 g L-1), moderadamente tolerantes a Na2SO4, (máxima acumulación de biomasa con 3.84 g L-1) y tolerantes a CaCl22H2O (máxima acumulación de biomasa con 6.40 g L-1). El objetivo del presente trabajo fue evaluar el porcentaje de emergencia de brotes de tubérculos de papa cv. Alpha en condiciones de estrés inducido por un mayor espectro salino que incluye además de las sales y concentraciones indicadas por Sánchez et al. (2003), las salinidades sulfático-clorhídrica, clorhídrico-sulfática y sulfático-sódica presentes en las cuencas endorréicas del Valle de Toluca en México, el altiplano de Nuevo León y las estribaciones de la Laguna del Carmen en Tlaxcala, donde se produce papa en México.
MATERIALES Y MÉTODOS
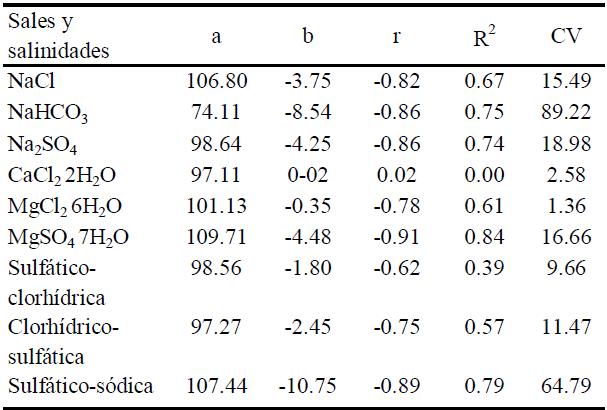
En condiciones experimentales de invernadero se sembraron tubérculos de papa (Solanum tuberosum L.) en macetas de plástico, preparadas con 10 kg de agrolita y una cubierta de tezontle de 2 cm de espesor, a una densidad de dos propágulos por maceta, espaciados 15 cm uno de otro; a cada maceta se aplicó un volumen de 2 L de solución salina. Los tratamientos diseñados como experimentos independientes (una sal por tratamiento), utilizaron sales de: NaCl, NaHCO3, Na2SO4, CaCl22H2O, MgCl26H2O y MgSO47H2O, y tres salinidades cualitativas: sulfático-clorhídrica, clorhídrico-sulfática y sulfático-sódica, más un testigo (C0) con agua destilada y seis niveles de salinidad con concentraciones de 1.13, 2.48, 3.84, 5.19, 6.40 y 9.60 g L-1 que teóricamente corresponden a conductividades eléctricas en la solución de 1.77, 3.88, 6.00, 8.11, 10 y 15 dS m-1 respectivamente y que de acuerdo con Maas y Hoffman (1977) disminuyen el crecimiento o rendimiento en el orden de 0 (C1), 25 (C2), 50 (C3), 75 (C4), 100 (C5) y 150% (C6). Se estableció un modelo bloques al azar con tres réplicas por tratamiento. Las soluciones se prepararon en laboratorio; las de las salinidades sulfático-clorhídrica, clorhídrico-sulfática y sulfático-sódica se obtuvieron mediante las relaciones aniónicas Cl-1/SO4 -2 = 1.5;Cl-1/SO4 -2 = 0.6 y HCO-3/Cl-1 + SO4 -2 = 2 respectivamente, establecidas por Arinushkina (citado por Szabolcs, 1989). En las dos primeras salinidades las fuentes de sales fueron NaCl y MgSO4 y para la salinidad sulfático-sódica las fuentes fueron MgCl2, Na2SO4 y NaHCO3 A las soluciones se les determinó pH con un potenciómetro, conductividad eléctrica (CE) con un conductímetro y potencial osmótico Ψπ (MPa) con un osmómetro. Adicionalmente, y con base en la fórmula de fertilización 40-40-20, se preparó una solución nutritiva Steiner (1984) la cual fue vertida a cada uno de los contenedores de las soluciones salinas que a su vez se suministraron a las macetas sembradas; el riego consistió en la reposición de agua consumida por evapotranspiración cada tercer día, a fin de mantener unidades experimentales a peso constante y a capacidad de campo; en invernadero se registraron temperaturas de 15 a 30 °C. El experimento tuvo una duración de 2 meses (del 18 de febrero al 18 de abril de 2007) periodo suficiente para evaluar la emergencia de brotes. Durante ese lapso los brotes se asperjaron con Ridomil Gold MZ 68 WP, fungicida sistémico, lo que permitió mantener periodos prolongados de protección contra tizón tardío (Phytophtora infestans); cada tercer día se hicieron observaciones para registrar el porcentaje de emergencia de brotes (número de días a la emergencia y número de brotes formados). El porcentaje de brotes emergidos se obtuvo considerando un total de cuatro brotes por tubérculo, es decir, ocho brotes por maceta, obteniéndose los promedios respectivos; los datos recabados se agruparon en fechas de brotación a 16, 23, 30 y 50 días, de esta forma la emergencia de brotes total se registró de forma acumulada en apego al método de Ayers y Hayward (1948). De acuerdo a Moorby y Milthorpe (1983) un brote de papa emergido se consideró como el tallo visible inmediatamente arriba del sustrato. Finalmente se calibró un modelo lineal de predicción del porcentaje total de brotes emergidos por tubérculo (PEB) 50 días después de la emergencia, en función de la CE de las soluciones mediante la ecuación: y = a + bx. Donde: y = porcentaje de brotes emergidos por tubérculo (%), A = ordenada al origen, b = pendiente de la recta o número de unidades (%) que aumenta o disminuye el PEB por unidad de incremento de la CE en la solución, x = CE de la solución (dS m-1). Los análisis de regresión lineal y de varianza, para determinar diferencias significativas con P ≤ 0.05, se efectuaron mediante el programa sistema de análisis estadístico (SAS Institute, 2006).
RESULTADOS Y DISCUSIÓN
Porcentaje de Brotes Emergidos
De los datos del Cuadro 1 se desprende para todos los casos incluyendo el testigo, un retraso en la emergencia de brotes, ya que cerca de 90% de los mismos ocurrió 30 días después de la siembra. La emergencia de brotes en condiciones ambientales adecuadas comprende un periodo de 16 días; en parte tal retraso se atribuye al estado de madurez fisiológica de los tubérculos semilla, pues tubérculos madre con prolongados periodos de almacenamiento tienden a aumentar su lapso de latencia y en consecuencia manifiestan cierto grado de envejecimiento (Moorby y Milthorpe, 1983), sin embargo, el retraso en la emergencia de brotes en lo fundamental es resultado del efecto del tipo de sal y de su concentración, de tal manera que en el rubro de porcentaje de brotes emergidos a los 16 días, se presenta un descenso abrupto de dicho porcentaje en los tubérculos expuestos a NaCl, NaHCO3 y a las salinidades sulfático-clorhídrica, clorhídrico-sulfática y sulfático-sódica a partir de 2.48 g L-1, en tanto que para los tratamientos Na2SO4 y MgSO47H2O, el porcentaje de brotes emergidos fue muy disminuido a 3.84 g L-1; por su parte los tratamientos CaCl22H2O y MgCl26H2O disminuyeron notablemente el porcentaje a una concentración de 5.19 g L-1, lo que denota niveles de tolerancia a sales en los tubérculos semilla.
Cuadro 1 Efecto de la conductividad eléctrica (CE), pH y potencial osmótico (Ψπ) de soluciones salinas experimentales sobre el porcentaje de brotación de tubérculos de papa a diferentes días después de la siembra.
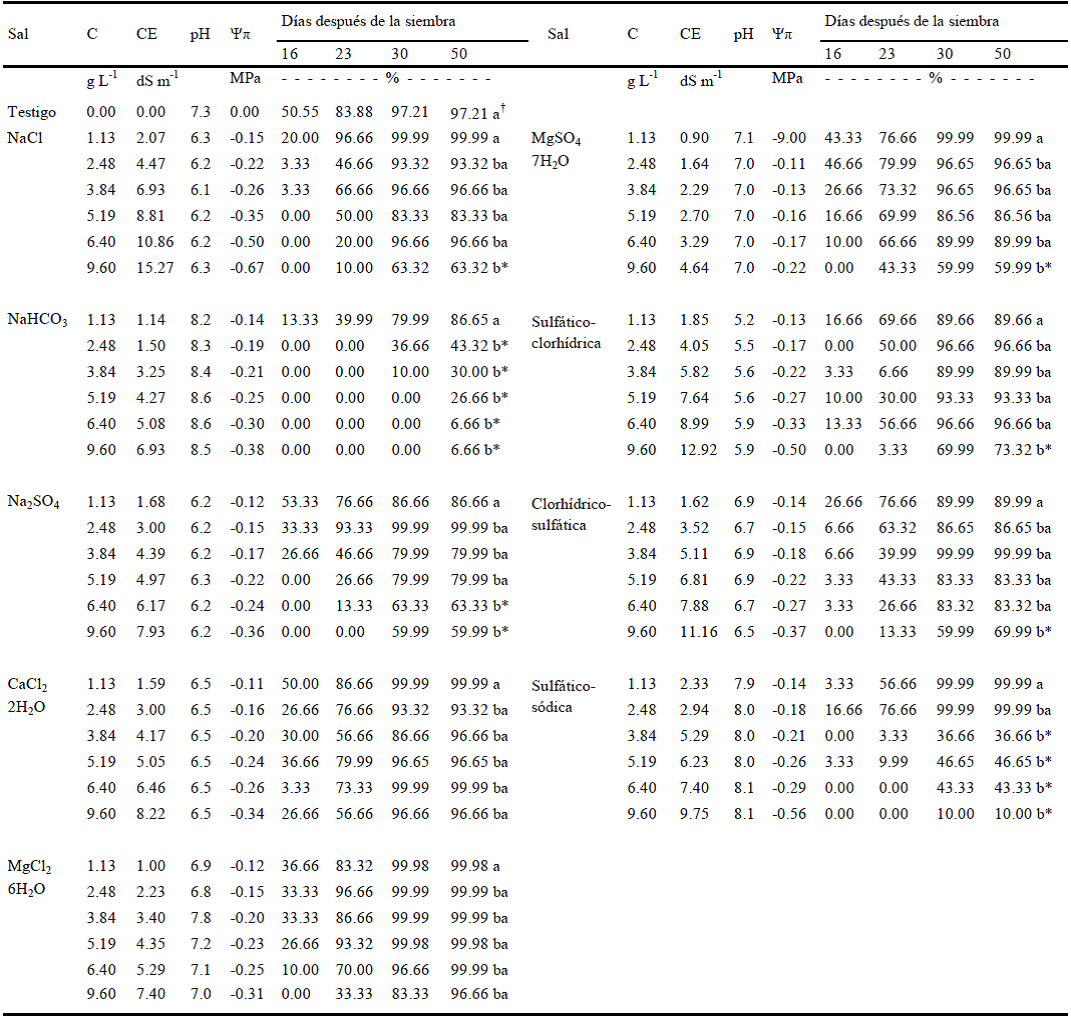
Prueba de medias entre tipo de sal para la variable emergencia de brotes a 50 días de la siembra (P > F = 0.0001); † Prueba de medias entre concentraciones para la variable emergencia de brotes a 50 días de la siembra (P > F = 0.002). * Significancia. Medias con la misma letra no son significativamente diferentes, Tukey, 0.05.
En general, el retraso en la emergencia de brotes es una respuesta de los tubérculos al efecto osmótico y tóxico que producen las sales al interior de este órgano vegetal, ya que el déficit hídrico y la toxicidad iónica inhiben la síntesis de la nitrato reductasa y las poliaminas, alteran los niveles hormonales (mayor producción de ácido absícico que de giberelinas), y deshidratan o desprenden el protoplasma de la pared celular, todo lo cual incrementa el periodo de quiescencia. Una vez que el tubérculo es capaz de adecuarse al estrés, incentiva sus procesos metabólicos y uso de substancias de reserva para promover emergencia de brotes, lo que según Strogonov (1964) y Ashraf (1983) está relacionado con una regulación osmótica al interior del tubérculo o con condiciones hídricas del substrato que favorecen una periodicidad del crecimiento como el riego. El estrés hídrico o tóxico guarda relación con la disminución del potencial osmótico en las soluciones salinas que presentó el siguiente orden: NaCl > salinidad sulfático-sódica > salinidad sulfático-clorhídrica > NaHCO3> salinidad clorhídrico-sulfática > Na2SO4> CaCl22H2O > MgCl26H2O > MgSO47H2O; la secuencia es determinada por las características químicas de cada soluto tales como: su peso molecular, grado de ionización y coeficiente osmótico (Hasegawa et al., 2000); lo que permite establecer criterios definidos sobre el efecto de las sales en el crecimiento y desarrollo de las plantas, el cual puede ser predominantemente osmótico o tóxico (Leidi y Pardo, 2002), de tal manera que el efecto de las sales en la emergencia de brotes de tubérculos de papa, puede ser de carácter osmótico como el producido por la sal NaCl o tóxico generado por la sal MgSO47H2O (Sánchez et al., 2003).
Si se considera el porcentaje total de brotes emergidos 50 días después de la siembra, la sal NaHCO3 y la salinidad sulfático-sódica registraron el menor porcentaje de emergencia de brotes, tal disminución se registró para NaHCO3 a 2.48 g L-1 y para salinidad sulfático sódica a 3.84 g L-1, a mayores concentraciones la emergencia de brotes disminuyó en forma notoria, lo que indica que los tubérculos de papa son muy sensibles para emerger brotes en condiciones sódico alcalinas; al respecto Moorby y Milthorpe (1983) señalan que en condiciones de pH superiores a 7.5 los tubérculos de papa inhiben la emergencia de brotes, situación contraria aloquesucedeenunintervalodepHde6a6.5,quela favorecen, comportamiento que coincide con los datos de pH y porcentajes de emergencia registrados en los tratamientos de sales y salinidades no sódicas (Cuadro 1).
Un efecto diferente produjeron las sales magnésicas (MgCl26H2O y MgSO47H2O) que por su alta solubilidad (353 g L-1 y 275 g L-1) y toxicidad, inhibieron el crecimiento de la yema terminal del tubérculo con la consiguiente pérdida de dominancia apical, que favoreció la formación de múltiples brotes compitiendo entre sí por alimento, luz y agua, caso indeseable y de efectos negativos para la producción de papa. En este caso se registraron hasta siete brotes por tubérculo, de los cuales al menos cuatro presentaban raquitismo, clorosis en su ápice y escaso desarrollo longitudinal (0.5 cm aproximadamente), el resto de los brotes alcanzó una longitud de 2 a 2.5 cm. La pérdida de dominancia apical por estrés hídrico ha sido explicada a detalle por Moorby y Milthorpe (1983). En el tratamiento CaCl22H2O, sal de tipo neutro, cuyo efecto osmótico y tóxico es mínimo, se presentó una emergencia de brotes cercana a la normalidad (cuatro brotes por tubérculo semilla) en todas las concentraciones, con la característica de que estos eran más sanos y vigorosos que los del resto de los tratamientos (4 cm de longitud). En 2003 Sánchez et al. obtuvieron resultados similares. Por su parte, el efecto osmótico producido por las sales NaCl y Na2SO4, así como por las salinidades sulfático-clorhídrica y clorhídrico-sulfática, es la causa del retardo en la emergencia de brotes 16 días después de la siembra; el estrés hídrico además de alargar el periodo de latencia, provocó la pérdida de dominancia apical de la yema terminal, emergiendo hasta cinco brotes por tubérculo semilla, de los que tres al menos presentaron clorosis, raquitismo e inhibición del crecimiento (1 a 1.5 cm de longitud), lo que explica el aumento de porcentaje de emergencia a los 30 días de la siembra; el resto de los brotes denotaron buenas características de vigor (3 cm de longitud). Para este último periodo, las concentraciones salinas de 1.13 a 9.6 g L-1 indujeron en los tubérculos de papa altos porcentajes de emergencia de brotes en los tratamientos MgCl26H2O, CaCl22H2O, MgSO47H2O y NaCl. Las concentraciones de 1.13 a 6.40 g L-1 promovieron altos porcentajes de emergencia de brotes en los tratamientos Na2SO4, salinidad sulfático-clorhídrica y salinidad clorhídrico-sulfática, ello indica que a pesar del retardo en la emergencia y de la pérdida de dominancia apical provocada por la mayoría de estas sales, los tubérculos de papa tienen gran plasticidad para emerger brotes en estas condiciones salinas, lo que está vinculado al efecto nutritivo inicial producido por la concentración de dichas soluciones, ya que de acuerdo a lo señalado por Moorby y Millthorpe (1983) y Sánchez et al. (2008), esta tasa aumenta con el tiempo, entre otros factores con incrementos en la concentración de la solución nutritiva.
Modelos de Predicción de Brotación
Los modelos de calibración del porcentaje total de brotes emergidos en función de la CE de cada solución salina (Cuadro 2) predicen que al aumentar la CE disminuye el porcentaje total de brotes en el siguiente orden: MgCl26H2O < CaCl22H2O < salinidad sulfático- clorhídrica < NaCl < MgSO47H2O < salinidad clorhídrico- sulfática < Na2SO4 < salinidad sulfático-sódica < NaHCO3.
Efecto del Potencial Osmótico de las Soluciones Salinas
Las sales y salinidades disminuyeron el potencial osmótico de las soluciones conforme aumentó su concentración en el orden NaCl (0 a -0.67 MPa) > salinidad sulfático-sódica (0 a -0.56 MPa) > salinidad sulfático-clorhídrica (0 a -0.50 MPa), > NaHCO3 (0 a -0.38 MPa) > salinidad clorhídrico-sulfática (0 a -0.37 MPa), > Na2SO4 (0 a -0.36 MPa), > CaCl22H2O (0 a -0.34 MPa) > MgCl26H2O (0 a -0.31 MPa) > MgSO47H2O (0 a -0.22 MPa), de tal forma que las sales y salinidades que más redujeron el porcentaje de brotes emergidos en el intervalo de 0 a 9.6 g L-1, respecto del testigo, fueron: NaHCO3 y salinidad sulfático-sódica, con promedios de emergencia de 33.3 y 56.1% respectivamente, porcentajes que fueron significativamente diferentes respecto del resto de los tratamientos que presentaron mayores porcentajes de brotes emergidos, en el siguiente orden: Na2SO4 con 78.3%, salinidad clorhídrico-sulfática con 85.5%, MgSO47H2O con 88.3%, NaCl con 88.9%, salinidad sulfático-clorhídrica con 89.9%, CaCl22H2O con 97.2% y MgCl26H2O con 99.4%, en los que no hubo diferencias significativas (Figura 1).
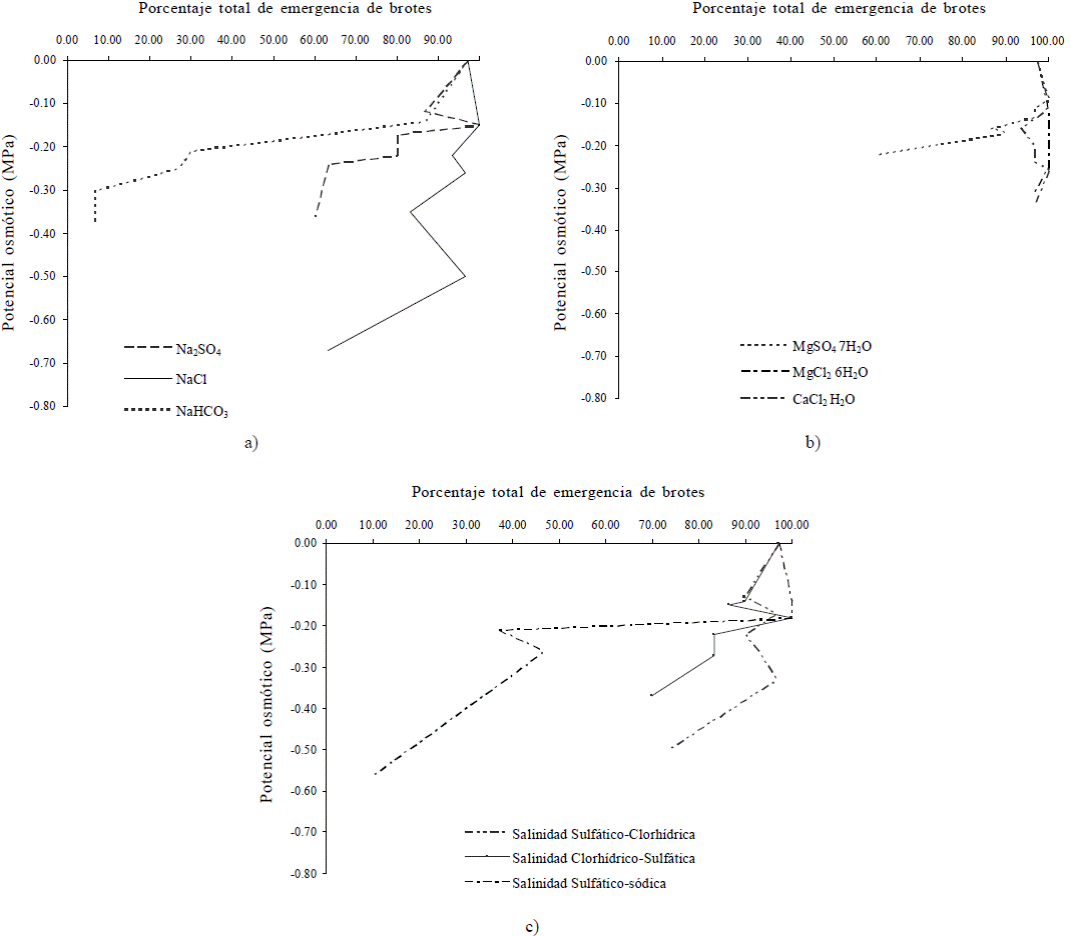
Figura 1 Efecto del potencial osmótico de soluciones salinas de: a) NaCl, NaHCO3, Na2SO4; b) CaCl22H2O, MgCl26H2O, MgSO47H2O; c) salinidad sulfático-clorhídrica, salinidad clorhídrico-sulfática y salinidad sulfático-sódica, en el porcentaje de emergencia de brotes de tubérculos de papa.
Este comportamiento denota el efecto osmótico-tóxico de las sales sódico-alcalinas en los tubérculos madre, cuyos procesos de hidrólisis inhiben la emergencia de brotes, e inducen la destrucción de tejidos vegetales (Strogonov, 1964; Velázquez, 2002). En forma análoga, se presenta el efecto osmótico de NaCl, salinidad sulfático-clorhídrica, Na2SO4 y salinidad clorhídrico- sulfática, que genera la pérdida de dominancia apical y por ende una brotación copiosa anormal, y finalmente el efecto tóxico de las sales MgCl26H2O y MgSO47H2O que inducen una pérdida de dominancia apical más aguda. Las variaciones en potencial osmótico de las sales y salinidades experimentadas, han sido debidamente señaladas por Ortega et al. (1989), su efecto en el crecimiento de algunas plantas cultivadas como el tomate de la misma familia que la papa fue analizado a detalle por Strogonov (1964). El comportamiento osmótico de CaCl22H2O no provocó ni disminución, ni aumento en exceso de la emergencia de brotes, ya que se trata de una sal neutra que generó una brotación de tipo normal, lo que según Davies y Millard (1985) se atribuye al crecimiento celular que promueve el ion calcio absorbido por los tubérculos madre.
CONCLUSIONES
- En la fase de emergencia, los brotes de tubérculos de papa cv. Alpha, son sensibles a NaHCO3 y a la salinidad sulfático-sódica, ya que su máximo porcentaje de emergencia se registró de 2.48 g L-1 a 3.84 g L-1 de concentración; a concentraciones superiores se produjo una disminución notoria del número de brotes emergidos. Son también moderadamente sensibles a NaCl, Na2SO4, MgCl26H2O, MgSO47H2O, salinidad sulfático-clorhídrica y salinidad clorhídrico-sulfática, su máximo porcentaje de emergencia de brotes se registró de 5.19 a 6.40 g L-1 de concentración, esta emergencia de brotes se caracterizó por la pérdida de dominancia apical de los tubérculos. Finalmente se consideran tolerantes a CaCl22H2O, ya que su máximo porcentaje de emergencia de brotes se registró a 9.60 g L-1 de concentración, con brotes normales.
- Se distinguen tres tipos de efectos salinos en la emergencia de brotes: a) hidrólisis alcalina y toxicidad inducida por las sales sódicas en las yemas promotoras de la brotación de tubérculos, que implica destrucción de meristemos y restricción de crecimiento de brotes; b) pérdida de dominancia apical por toxicidad de las sales MgCl26H2O y MgSO47H2O, con formación de múltiples brotes, efecto indeseable y negativo en la producción de papa y c) pérdida de dominancia apical menos pronunciada que la anterior, provocada por el estrés osmótico de las sales NaCl y Na2SO4, así como por las salinidades sulfático-clorhídrica y clorhídrico-sulfática; en estos tratamientos los brotes exhibieron menos daño y más vigor que los de los casos anteriores. Los brotes en CaCl22H2O presentaron características normales.
- La pérdida de dominancia apical de la yema terminal de los tubérculos madre, es resultado de la variación del estrés salino provocada por el riego, lo que induce periodicidad del crecimiento, caso indispensable y de efectos negativos para el cultivo de papa.

