











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Los frutos de calabacita (Cucurbita pepo L.) se consumen principalmente inmaduros, como fruto verdura, tanto en el mercado nacional como en el de exportación. En 2009 se cultivaron 30 629 ha en México, con un rendimiento medio de 15.8 Mg ha-1, inferior al de Holanda (70.0 Mg ha-1), España (68.18 Mg ha-1) y Francia (37.27 Mg ha-1), y apenas superior al promedio mundial (13.62 Mg ha-1) (FAO, 2010), lo que evidencia la necesidad de elevar el rendimiento nacional. En 2009 el país exportó 350 939 Mg (SAGARPA, 2010). El fruto de la calabacita es apreciado porque contiene pocas calorías, es rico en vitaminas (C, E, B1 ,B2 y β-caroteno) y minerales (K, Ca, Fe, Zn, Mn, Mg, P, B, Cu y N) (Danilchenko et al., 2001). Una variedad de calabacita muy cultivada en México es la Gray Zucchini, que se caracteriza por ser herbácea y precoz, y por iniciar la producción 50 d después de la siembra. Su híbrido comercial Tala (Seminis®) tiene la ventaja de presentar plantas uniformes y vigorosas.
La calabacita es una planta C3, las que en general se consideran de manor eficiencia fisiológica que las plantas C4, aunque existen muchas especies con altas tasas de asimilación de CO2 como el arroz (Oryza sativa), la soya (Glycine max) y el girasol (Helianthus annus) (Villalobos, 2001).
Entre los nutrimentos que más influyen en el rendimiento de los cultivos están el N y el K. El N favorece el desarrollo vegetativo e intensifica el color verde de las hojas; es constituyente de componentes celulares esenciales, como aminoácidos, proteínas y ácidos nucleicos; además, es regulador del P, K y otros nutrimentos; y mejora la suculencia de muchos cultivos. Por su parte, el K da vigor al crecimiento, es esencial para la formación del almidón y la hidrólisis de los azúcares, promueve la translocación de asimilados desde las hojas y favorece la fotosíntesis, participa en la regulación del potencial osmótico celular, activa enzimas, participa en la liberación de proteínas desde los ribosomas, es necesario en la síntesis de clorofila, favorece el desarrollo de raíces, y regula el balance del N y P (Taiz y Zeiger, 2006; Alcántar y Trejo, 2007).
En tomate (Lycopersicon esculentum Mill.) se logró incrementar la producciónde biomasa aérea y la calidad de fruto, al aplicar K, a una concentración de 3 a 9 meq L-1 (Bugarín et al., 2002). En brócoli (Brassica oleraceae L.) Vidal et al. (2006) elevaron el rendimiento mediante aplicaciones de K, y en plátano (Musa paradisiaca L.) cv. Gran Enano Orozco y Pérez (2006) detectaron interacción de N y humedad sobre el rendimiento y calidad de fruto.
Mediante fertilización con N se aumentó la concentración de clorofila en hojas de calabacita, en comparación con el testigo (Aroiee y Omidbaigi, 2004); y en calabacita cultivada sin labranza, al aumentar el N permitió incrementar el rendimiento de fruto (Harrelson et al., 2004). Pero en calabaza acanalada (Telfairia occidentalis Hook F.) un mayor contenido de N favoreció la incidencia de la enfermedad mancha foliar causada por Phoma sorghina, mientras que el K no afectó el desarrollo de la enfermedad (Nwufoy Agu, 2001). Dosis de 6 mM de N favoreció una eficiente producción y distribución de biomasa en raíz y vástago en frijol ejotero cv. Strike (Sánchez et al., 2005).
En girasol (Helianthus annuus L.) cultivado con altas dosis de N y densidad de siembra se logró una mayor producción de biomasa y rendimiento de semilla (Vega et al., 2001); similarmente, Escalante (2001) encontró que las aplicación de más N incrementó la biomasa y rendimiento de semilla en dos variedades de girasol. Con dosis de 250 kg ha-1 de N, el tomate elevó su rendimiento y su calidad poscosecha (Villarreal et al., 2002). También en chile Jalapeño (Capsicum annum L.) se encontró que a mayor dosis de N y K se obtuvo mayor rendimiento (Alonso et al., 2002). La fertilización con P y K en calabacita cv. Styriaca aumentaron el rendimiento de frutos y el de semilla (Ali et al., 1999).
Los nutrimentos también pueden mejorar la calidad del fruto y diversas características agronómicas. Por ejemplo, en melón (Cucumis melo L.) cv. Cantaloupe se aumentó rendimiento y contenido de sólidos solubles en fruto mediante aplicación de mayor dosis de N (Pérez et al., 2004).Las aplicaciones de N y etefón en Cucurbita moschata L. cv. Co-1 no solo aumentaron el número de flores pistiladas, el rendimiento de fruto y de semilla, sino también la germinación (Adirai et al., 2001). Las dosis de 476 kg de N, 500 kg P2O5 y 548 kg de K2 O ha-1 incrementaron el rendimiento de fruto comercial, grosor de pulpa, intensidad del color en C. moschata L., en comparación con el testigo sin fertilizar (Damarany y Farag, 1994). En girasol las altas dosis de N produjeron un mayor índice y duración de área foliar, mayor radiación interceptada y más alta producción de biomasa y rendimiento de semilla y aceite (Escalante, 1999). Las hojas de tomate crecidas con bajo suministro de N y con alta relación C/N, contenían más carbohidratos solubles, y fueron 2.5 veces más susceptibles a lesión primaria por Botrytis cinerea, que hojas de plantas crecidas con alta disposición de N y menor relación C/N (Hoffland et al., 1999).
La planta de calabacita es capaz de producir una gran cantidad de botones florales pero apenas siete llegan a convertirse en frutos, en promedio, debido a que el área foliar sólo dura 90 d y con ello se agota la fuente de asimilados (Sedano et al., 2005). Lo anterior ha sugerido la necesidad de alargar la longevidad del follaje que provee fotoasimilados para que más botones florales formen frutos y se aumente el rendimiento. Una alternativa para alargar la vida foliar es mediante el uso de la fertilización nitrogenada que favorece el desarrollo foliar y el rendimiento, y con K que favorece el transporte de asimilados, la actividad enzimática, la asimilación de CO2 y alarga la vida de las plantas (Taiz y Zeiger, 2006; Mengel y Haeder, 1977). Dado que la planta y sus frutos extraen importantes cantidades de nutrimentos (Valadez, 1998), se hace necesario probar altos niveles de N y K. La necesidad de evaluar K es debido a que se necesita elevar la longevidad del cultivo y con ello aumentar el número de frutos a cosechar. Otro aspecto a considerar es la calidad del fruto cosechado y en poscosecha, en especial cuando se destina a la exportación. Actualmente, en la calabacita se consumen dos tamaños de fruto: de 12 a 15 cm de longitud para el mercado nacional, y de 16 a 25 cm para exportación (ASERCA, 1999; Valadez, 1998).
El objetivo de esta investigación fue evaluar el efecto de dosis altas de N y K sobre la eficiencia fisiológica y el rendimiento y calidad de los frutos de calabacita tipo verdura, tanto para el mercado nacional como para el de exportación considerando medir niveles mayores a las recomendaciones de Valadez(1998) y Sedano(2005).
MATERIALES Y MÉTODOS
Sitio Experimental
El campo se ubicó a 19° 27’ 30” N y 98° 54’14” O, una altitud de 2241 m, con temperatura media anual de 14.8 °C y precipitación de 675 mm anuales, cuyo clima es templado, el más seco de los subhúmedos, con oscilación térmica entre 5 y 7 °C (entre el mes más cálido respecto del mes más frío) y régimen térmico tipo ganges (el mes más cálido antes del solsticio de verano), clasificado como C (w0) (w) b (i')g por García (1981). El suelo utilizado es arcilloso clasificado como un vertisoldístrico, del grupo TypicHapluster (USDA, 1994).
Cultivar
La semilla de calabacita cv Tala (Seminis®) se sembró en campo, con riego, durante el ciclo primavera- verano, a una densidad de siembra de 23 000 plantas por ha, en un lote de 2000 m2 en Montecillo, estado de México.
Se utilizó un diseño factorial 3 × 3 (tres dosis de N: 150, 240 y 330 kg ha-1; tres dosis de K2 O: 90, 120 y 150 kg ha-1). Los nueve tratamientos de fertilización se establecieron en el campo en arreglo completamente al azar con tres repeticiones; el tamaño de parcela experimental fue cinco surcos de 90 cm de ancho por 6 m de longitud.
Las fuentes de fertilizantes fueron sulfato de amonio ((NH4)2 SO4), superfosfato triple de calcio (Ca (H2PO4)2) y nitrato de potasio (KNO3). La primera fertilización se aplicó a los 10 d de la siembra, con todo el P y K y la mitad de N (primero aplicando KNO3 y completando con (NH4)2SO4; el resto del N se aplicó tres semanas después con sólo (NH4)2SO4.
Tratamientos y Diseño Experimental
En este experimento se probaron nueve dosis de fertilización, resultantes de combinar tres dosis de N (150, 240 y 330 kg ha-1), con tres dosis de K2O (90, 120 y 150 kg ha-1). Considerando el terreno uniforme, los nueve tratamientos se dispusieron en un diseño completamente al azar con arreglo factorial 3 × 3, con tres repeticiones.
Variables Medidas
Asimilación de CO2. Se hicieron dos mediciones de tasas de asimilación neta de CO2 en la hoja madura más joven, registradas al medio día y a pleno sol, a los 67 y a los 83 días después de la siembra (dds), en plena fructificación y final del ciclo del cultivo, en tres repeticiones en cada parcela con el equipo LI-6200® (LI-COR Lincoln, NE, EUA).
Rendimiento. Para cuantificar el rendimiento en peso fresco y el número de frutos por planta, en cada parcela se etiquetaron cuatro plantas con competencia completa. Los frutos se clasificaron en frutos tamaño 1 y 2; tamaños en los que se consume y comercializa la calabacita. Esta clasificación es: frutos para mercado nacional (Tipo 1) de 12 a 15 cm y de exportación (Tipo2) de 16 a 25 cm de longitud. Los frutos se cortaron cada 2 d desde el inicio de fructificación (50 dds) hasta el final del ciclo (105 dds).
Calidad del fruto. Las variables consideradas fueron: color externo, firmeza y contenido de azúcares solubles totales, midiendo cinco frutos de los dos tamaños por tratamiento. El color se midió con un colorímetro de reflexión Hunter Lab® (Reston, VI, EUA), en dos lados opuestos del fruto. Se registraron los valores L, a y b, con los que se calcularon las variables denominadas: croma = √a2+b2) y °Hue = (tan-1 b/a), L indica la claridad o luminosidad, a es la dirección del rojo, -a es la dirección del verde, b es la del amarillo, y -b es la del azul, c es croma y hue es el ángulo de color (McGuire, 1992). Los azúcares totales (mg mL-1) se determinaron, por el método de antrona (Witham et al., 1971). La firmeza de los frutos se midió con un texturómetro Chatillón (Warner Instruments, GR, EUA) y se registraron los datos de resistencia a la penetración en kg cm-2.
Manejo Agronómico del Cultivo
Se proporcionaron cinco riegos, uno a la siembra y los otros cuatro cada 10 d, posteriormente ya no fue necesario regar por que se había establecido la temporada de lluvias. Se eliminó la maleza 8 d después de emergencia, de forma manual con azadón; se aplicaron bactericidas y fungicidas preventivos (Terramicina® y Rydomil®), para prevenir infecciones causadas por Erwinia spp y por Ryzoctonia spp; Rovral® y Neem® contra cenicilla vellosa (Pseudoperonospora cubensis) y Neem (PHC™ NEEM) contra mosquita blanca (Bemisia tabaci Gennadius).
Análisis Estadísticos
Las figuras y las curvas de regresión se trazaron mediante el programa de Excel (Microsoft Excel). Los análisis de varianza, regresión y las pruebas de medias de Tukey (P ≤ 0.05 y P ≤ 0.10) para fotosíntesis, rendimiento y calidad de fruto, se hicieron con el programa estadístico SAS (SAS Institute, 1999), bajo un diseño experimental completamente al azar.
RESULTADOS Y DISCUSIÓN
Rendimiento y Número de Frutos
El análisis de varianza realizado para observar el efecto de N y K sobre el rendimiento de frutos de calabacita cv. Tala de tamaño 1 (para el mercado nacional), mostró que los niveles de K y la interacción N × K no afectaron a esta variable (P ≤ 0.10), pues sólo hubo diferencias (P ≤ 0.05) para dosis de N (Cuadros 1 y 2). Al respecto, el rendimiento del nivel más alto de N (330 kg ha-1) superó al del nivel más bajo (150 kg ha-1), aunque no hubo diferencias (P ≤ 0.10) entre tratamientos de N y K en el número de frutos de tamaño 1 por planta; por tanto, las diferencias en rendimiento podrían atribuirse a diferencias en el peso individual de los frutos. La curva de respuesta de frutos de tamaño 1 a la dosis de N, ajustada mediante una ecuación cuadrática (Figura 1), muestra que el rendimiento de este tamaño de fruto aumenta al elevar la dosis de N de 150 a 240 kg ha-1, que al subirla de 240 a 330 kg ha-1. Es de suponer entonces, que la dosis de 240 kg ha-1 de N es cercana a la dosis que maximiza el rendimiento de este cultivar. En cuanto al número de frutos de tamaño 1, todos los tratamientos produjeron al menos el doble de frutos (14.71 frutos por planta) que el obtenido por Sedano et al. (2005) (siete frutos por planta). En rendimiento y número de frutos del tamaño 2 (Cuadros 1 y 2) no hubo efecto significativo de N y K ni de la interacción N × K (P ≤ 0.10). Sin embargo, el número de frutos del tamaño 2 (9.48) fue inferior al número de frutos de tamaño 1 (Cuadro 1), lo que se atribuye a que los frutos grandes compiten más entre sí por asimilados, que los frutos pequeños, como se pudo constatar previamente, cuando se dejaron plantas sin cosechar, observándose que solo crecía un fruto y los frutos pequeños abortaban.
Cuadro 1 Rendimiento y número de frutos de calabacita de tamaño 1 (12-15 cm) para el mercado nacional, y tamaño 2 (16-25 cm) para mercado de exportación.
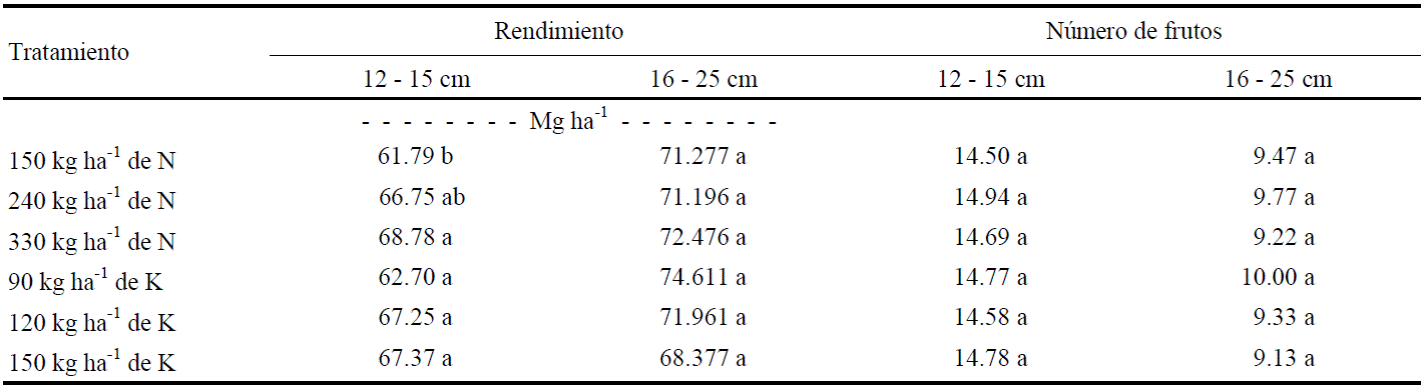
Medias con letras distintas en la misma columna son estadísticamente diferentes (P ≤ 0.05).
Cuadro 2 Valores de probabilidad de error de los análisis de varianza efectuados para rendimiento y número de frutos de los tamaños 1(consumo nacional) y 2 (de exportación), y para tasas de fotosíntesis neta en dos fechas (06/06/05 y 22/06/05) de evaluación.
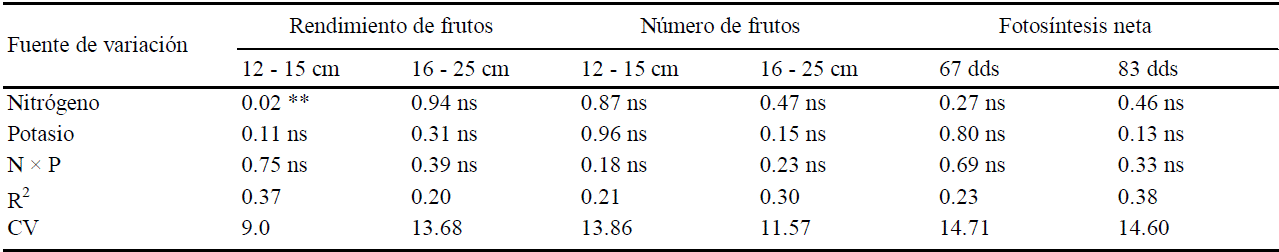
ns, *, ** y *** = no significativo y significativo a 0.1 0.05 y 0.01 de probabilidad, respectivamente; CV = coeficiente de variación; dds = días después de la siembra.
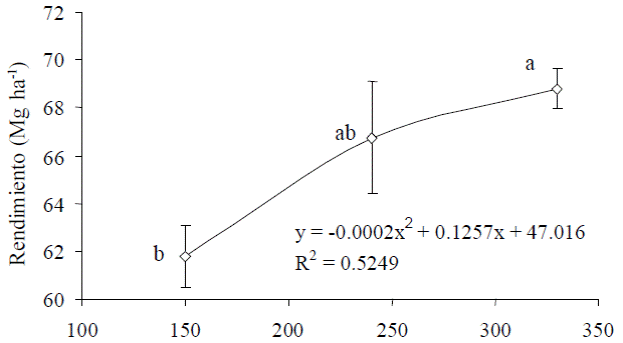
Figura 1 Relación entre dosis de N y rendimiento de fruto de calabacita de tamaño 1. Las barras verticales representan la desviación estándar (n = 4). Letras distintas al lado de cada dato indican diferencias significativas (P ≤ 0.05) entre tratamientos.
Los mayores rendimientos obtenidos en frutos de tamaño 1 al aplicar más N, coinciden con lo reportado por Harrelson et al. (2004) para calabacita con cero labranza cuando se aumentó la dosis de N, y por Damarany y Farag (1994) y Adirai et al. (2001) en Cucurbita moschata. En girasol (Helianthus annus L.), Escalante (1999, 2001) también encontró que altas dosis de N aumentaron biomasa y el rendimiento de semilla. De igual modo, Vega et al. (2001) reportaron que altas dosis de N y alta densidad de siembra aumentaron la biomasa y rendimiento de semilla de girasol; y Pérez et al. (2004) encontraron que al aumentar dosis de N se incrementaba el rendimiento de fruto de melón. Pero no coincide con lo encontrado en calabacita cv. Styriaca por Alí et al. (1999) en respuesta a incrementos de K que sí aumentó el rendimiento y número de frutos y el rendimiento de semilla.
El rendimiento de fruto de calabacita de tamaño 2 (para exportación) no presentó diferencias significativas (P ≤ 0.10) entre las dosis de N y K probadas. Aún así, destaca que el rendimiento mínimo obtenido de este fruto (68 Mg ha-1) es 3.2 veces mayor al reportado (21 Mg ha-1) por Sedano et al. (2005) y 4 veces mayor al promedio nacional de 15.8 Mg ha-1 (FAO, 2010).
Los presentes resultados coinciden parcialmente con los de otros investigadores en cuanto al efecto de N y K en el rendimiento de cultivos. Por ejemplo, en plátano Gran Enano Orozco y Pérez (2006) detectaron efecto de interacción de N y humedad sobre rendimiento y calidad de fruto. Vidal et al. (2006) encontraron aumento en rendimiento en brócoli var. Itálica por efecto del K. Al elevar la dosis de N, Villarreal et al. (2002) obtuvieron alta calidad postcosecha y rendimiento de tomate. En chile Jalapeño Alonso et al. (2002) obtuvieron mayor rendimiento de fruto con aumentos en las dosis de N y K. También al aumentar N, P y K se incrementó el rendimiento de fruto de C. moschata (Damarany y Farag, 1994). Esta aparente falta de concordancia con otros autores se atribuye a que en el presente estudio solamente se evaluaron altas dosis de N y K, puesto que la dosis más baja evaluada en este estudio (150N- 90P-90K), corresponde a una dosis alta de fertilización en comparación con las dosis comerciales para calabacita, como las recomendadas por Valadez (1998) de 80-60-00, 130-90-00 y 120-80-00 de N-P-K, respectivamente para alcanzar un rendimiento de 19.71 Mg ha-1.
Fotosíntesis
En las dos fechas en que se midió la tasa neta de asimilación de CO2, no se detectaron diferencias significativas entre tratamientos (P ≤ 0.10). No obstante, se ha reportado que la fotosíntesis se favorece por el N porque este elemento aumenta la cantidad de clorofila (Taiz y Zeiger, 2006; Buckman y Brady, 1980; Aroiee y Omidbaigi, 2004). El índice de área foliar y la radiación interceptada (Escalante, 1999). También el K favorece el desarrollo de la clorofila y regula el balance de N y P (Taiz y Zeiger, 2006; Buckman y Brady, 1980). Lo anterior sugiere que aún la dosis más baja de fertilización aquí aplicada (150N-90P-90K) fue suficiente para lograr una alta tasa de asimilación, de 41 y 38 μmoles de CO2 m-2 s-1 en la primera y segunda evaluación, respectivamente.
Calidad de fruto
Azúcares totales. En contenido de azúcares totales en fruto para consumo nacional no hubo diferencias (P ≤ 0.10) entre tratamientos de N y K (Cuadro 3), lo que coincide con lo reportado en plátano cv. Gran Enano por Orozco y Pérez (2006), cultivo en el que el N no se afectó la calidad del fruto, aunque la interacción de N y humedad tuvo efecto positivo en rendimiento y calidad de fruto. También coincide con Salgado et al. (2001) quienes reportaron que la calidad del jugo de caña de azúcar no fue afectada por las dosis de fertilizantes probadas. En contraste, Pérez et al. (2004) encontraron que al aumentar dosis de N se incrementaba el rendimiento y contenido de sólidos solubles en fruto de melón.
Cuadro 3 Probabilidades de error para las variables de calidad del fruto, azúcares totales y firmeza, en frutos de tamaños 1 (consumo nacional) y 2 (de exportación), según los análisis de varianza.
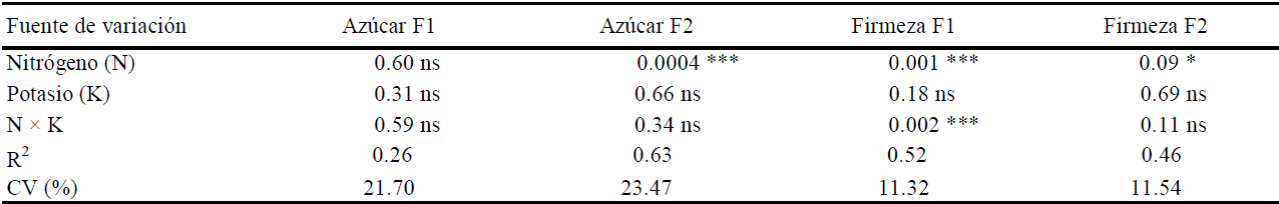
F1 = fruto1; F2 = fruto2, ns, *, ** y *** = no significativo y significativo a 0.1, 0.05, y 0.01 de probabilidad, respectivamente; CV = coeficiente de variación.
En el fruto de tamaño 2, el contenido de azúcares totales (Figura 2) se elevó (P ≤ 0.05) solamente al subir la dosis de N de 240 a 330 kg ha-1 pero no al aumentar la dosis de N de 150 a 240 kg ha-1. Mediante regresión poli-nomial se determinó que dicha respuesta al N se ajustó a una ecuación cuadrática con una R2= 0.88. En cambio, los niveles de K (de 90 a 150 kg ha-1) y la interacción N × K no afectaron el contenido de azúcares, a pesar de que este nutrimento participa en la hidrólisis de azúcares (Taiz y Zeiger, 2006; Buckman y Brady, 1980).

Figura 2 Azúcares totales en fruto de calabacita de tamaño 2 (de exportación 16-25 cm de longitud), en respuesta a la dosis aplicada de N.
Firmeza de frutos. En frutos de tamaño 1 esta variable fue afectada significativamente (P ≤ 0.05) por efecto de N y de la interacción N × K (Cuadro 3). En cuanto a la interacción, los máximos grados de firmeza se detectaron con la combinación de máxima dosis de N (330 kg ha-1) con la mínima de K (90 kg ha-1) o al contrario (Cuadro 4).
Cuadro 4 Efecto de la interacción N × K sobre la firmeza de fruto de calabacita de tamaño 1 (consumo nacional).
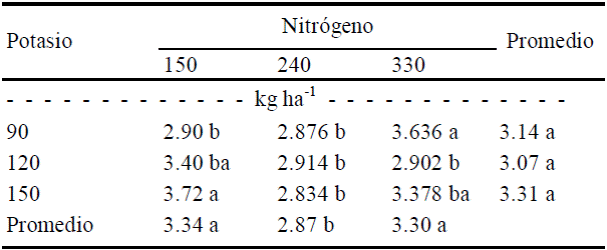
Valores con letra diferente en la misma columna, son diferentesestadísticamente (Tukey, P ≤ 0.10).
En cambio, el fruto de tamaño 2 solamente fue afectado por N (P≤ 0.1), y no por el K ni la interacción N×K (Cuadro 5). El efecto del N solamente fue significativo con la menor dosis de N y la mayor de K, esto es, conforme se incrementó el nivel del N, la firmeza del fruto disminuyó (Cuadro 5).
Cuadro 5 Efecto de la interacción N × K sobre la firmeza del fruto de calabacita de tamaño 2 (de exportación).
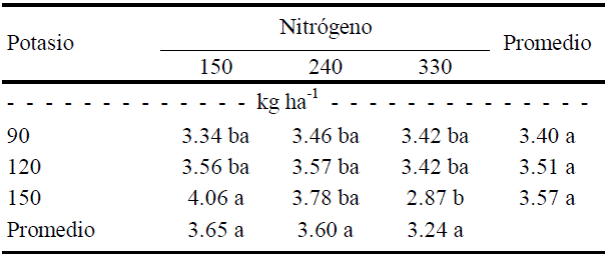
Valores con letra diferente en la misma columna, son diferentes estadísticamente (Tukey, P ≤ 0.10).
Color de fruto. En los frutos de tamaño 1 la luminosidad (L) no se vio afectada por los tratamientos de N y K aquí aplicados (Cuadro 6), aunque sí lo fueron otras características del color del fruto (P ≤ 0.10).
Cuadro 6 Análisis de varianza para color de frutos de calabacita de tamaño 1 (consumo nacional).
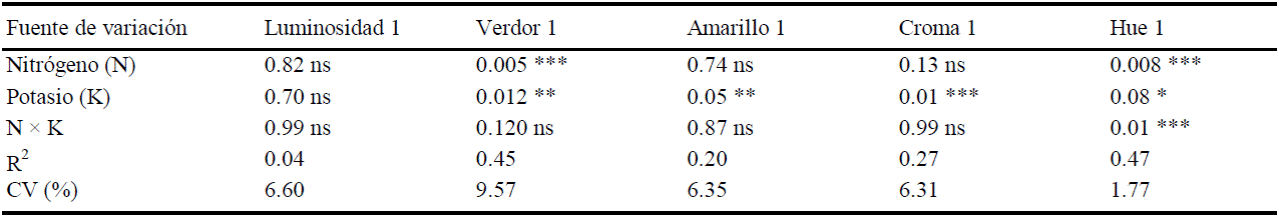
ns, *, ** y *** = no significativo y significativo a 0.1, 0.05, y 0.01 de probabilidad respectivamente. CV = coeficiente de variación.
Así el verdor de estos frutos (-a) mostró cambios significativos (P ≤ 0.05) por efecto simple de N y K, pero no de su interacción. Al respecto, el verdor aumentó en una unidad conforme aumentó la dosis de N de 150 a 330 kg ha-1 (Figura 3), y también aumentó una unidad conforme la dosis de K se elevó de 90 a 150 kg ha-1 (Figura 4).
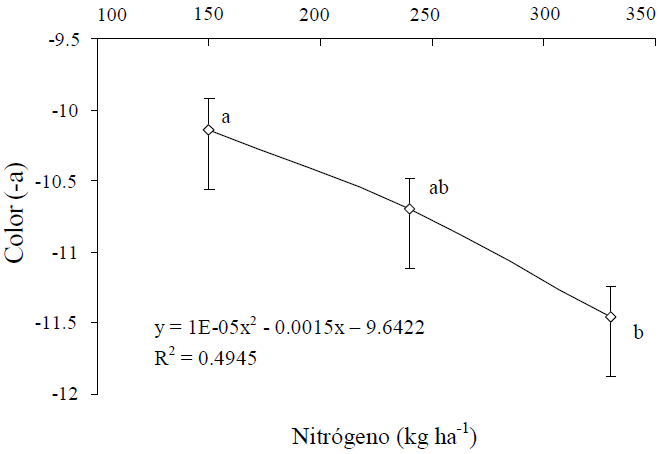
Figura 3 Efecto del N sobre el color verde (-a) del fruto de calabacita de tamaño 1 (para consumo nacional).

Figura 4 Efecto del K sobre el color verde (-a) del fruto de calabacita tamaño 1 (consumo nacional).
En el color amarillo (b) de los frutos de tamaño 1 sólo se detectó una tendencia a disminuir conforme aumentó la dosis de K (promedio, Cuadro 7), efecto que no resultó significativo (P ≤ 0.10) en la prueba de comparación de medias de Tukey, aunque la prueba de F marcó diferencias entre dosis de K (Cuadro 5).
Cuadro 7 Efecto de la interacción N × K sobre el color amarillo (b) del fruto de calabacita de tamaño 1 (consumo nacional).
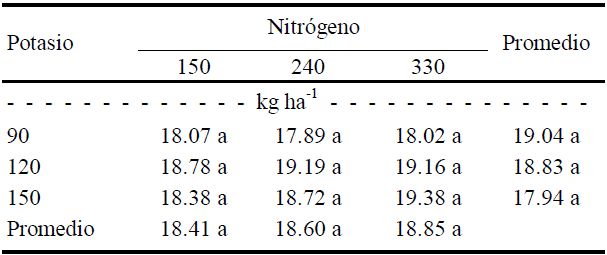
Valores con la letra diferente en la misma columna, indican diferencias estadísticas significativas (Tukey, P ≤ 0.10).
En la saturación del color o croma (c) de los frutos de tamaño 1 solamente hubo diferencias (P ≤ 0.05) por efecto de las dosis de K, pero no por efecto de N ni de la interacción N × K. Tal efecto consistió en la croma aumentó en una unidad (5%) al subir la dosis de K de 90 a 120 kg ha-1), pero la c ya no se modificó al aplicar más K (Figura 5). Es decir, los frutos con menor saturación de color fueron los que se cosecharon de plantas que recibieron la menor dosis de K (90 kg ha-1).
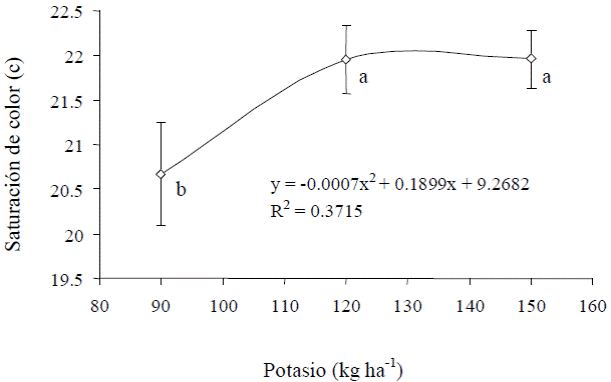
Figura 5 Curva de saturación de color en frutos de calabacita de tamaño 1 (consumo nacional), en respuesta a dosis de K. Letras distintas al lado de valores de c indican diferencias estadísticas significativas (P ≤ 0.05).
En contraste, los valores de hue (h) que también se relacionan con el verdor del fruto, resultaron afectados significativamente por las dosis de N (P ≤ 0.01) y por la interacción N × K (P ≤ 0.01), como se muestra en el Cuadro 6. Estos resultados confirman que las dosis de N aquí evaluadas combinadas con altos niveles de K favorecen el verdor de fruto de tamaño 1 y solo la combinación de 150 kg de Ncon 90 kg de K presentan menor valor de h (Cuadro 8).
Cuadro 8 Efecto de la interacción N × K sobre el color verde (h) de fruto de calabacita de tamaño 1 (consumo nacional).
Valores con letra diferente en la misma columna, indican diferencias estadísticas significativas (Tukey, P ≤ 0.10).
En los frutos de tamaño 2 la luminosidad (L) tampoco fue afectada por los tratamientos de N y K (P ≤ 0.01) (Cuadro 9), aunque las demás variables indicadoras del color de fruto sí fueron afectadas.
Cuadro 9 Probabilidades de error de los análisis de varianza para variables del color de frutos de calabacita de tamaño 2 (de exportación).

ns, *, ** y *** = no significativo y significativo a 0.1, 0.05, y 0.01 de probabilidad respectivamente. CV = coeficiente de variación.
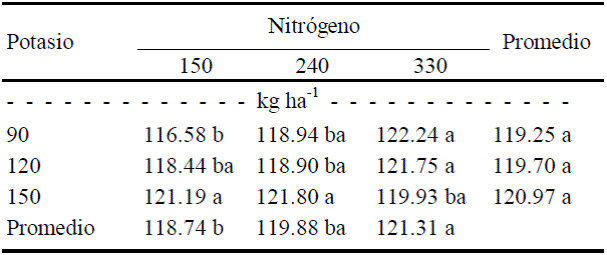
En el tipo de fruto 2 se detectó que el color verde aumentó (P ≤ 0.05) al elevar la dosis de N de 150 a 240 kg ha-1, y presenta menor valor absoluto en el tratamiento de 150 kg ha-1 de N y 120 de K, de modo que el máximo verdor se logró con la combinación de máximas dosis de N y K (Cuadro 10).
Cuadro 10 Efecto de la interacción N × K en el color verde (-a) del fruto de tamaño 2 (de exportación).
Valores con letra diferente en la misma columna, indican diferencias estadísticas significativas (Tukey, P ≤ 0.05).
Respecto al color amarillo (b), en frutos del tamaño 2 no hubo diferencias entre dosis de N o de K (P ≤ 0.05), pero si por efecto de la interacción N × K (Cuadros 9 y 11). Dicha interacción consistió básicamente en que el mayor grado de amarillamiento se dio con la combinación de máximas dosis de N (330 kg ha-1) y de K (150 kg ha-1), mientras que el mínimo amarillamiento resultó con la mínima dosis de N (150 kg ha-1) combinada con la máxima de K (150 kg ha-1). Estos resultados muestran que en el fruto de tamaño 2 también predomina el color verde, pero posee más tendencia al color amarillo que el fruto tamaño 1.
Cuadro 11 Efecto de la interacción N × K sobre el color amarillo (b) en frutos del tamaño 2 (de exportación).
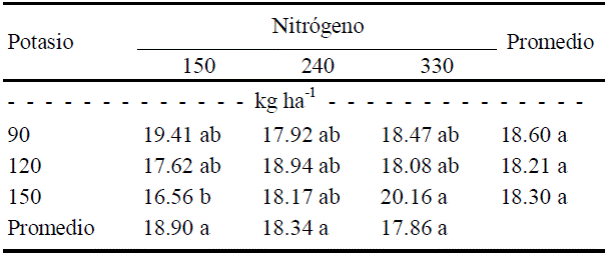
Medias con letra diferente en la misma columna, indican diferencias estadísticas significativas (Tukey, P ≤ 0.10).
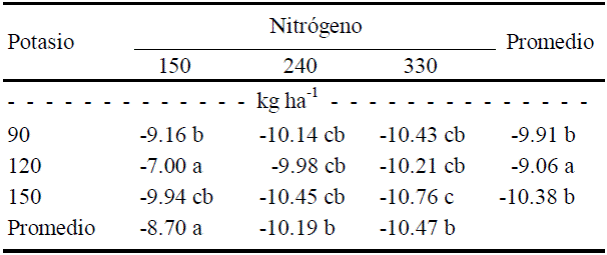
Con respecto a la saturación del color o croma (c) se registraron diferencias significativas (P ≤ 0.5) entre dosis de N y por efecto de la interacción (Cuadro 9). La saturación del color de fruto aumentó con las dosis de N (de 150 a 330 kg ha-1), mientras que la saturación se redujo al mínimo con las altas dosis de K (120 y 150 kg ha-1) combinadas con la menor de N (150 kg ha-1), como se muestra en el Cuadro 12. No se observa efecto de K, es decir, para lograr mayor saturación de color del fruto es conveniente una dosis de 90 kg ha-1 de K combinada con 330 kg ha-1 de N.
Cuadro 12 Efecto de la interacción N × K sobre el croma (c) de frutos de calabacita de tamaño 2 (para exportación).
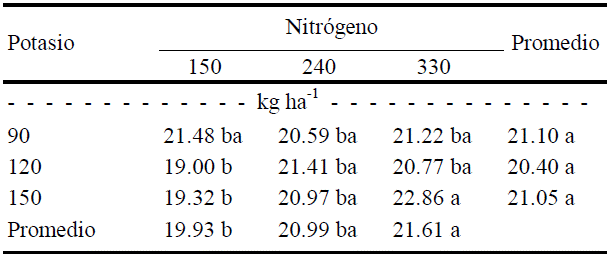
Valores con letra diferente en la misma columna, indican diferencias estadísticas significativas (Tukey, P ≤ 0.05).
La variable hue que indica el color verde del fruto también mostró diferencias (P ≤ 0.05) entre dosis de N y K y por efecto de la interacción N × K (Cuadro 9). Ello se debió a que el color verde se incrementó al aumentar la dosis de N de 150 a 240 kg ha-1; también hubo diferencias en los niveles de K, habiendo solamente un menor valor de h con el menor nivel de N (150 kg ha-1 y 120 kg ha-1 de K) (Cuadro 13).

CONCLUSIONES
- El rendimiento de frutos de tamaño 1 para consumo nacional se incrementó de 62 a 68 Mg ha-1 al aumentar la dosis de N de 150 a 330 kg ha-1, pero no cambió al elevar la dosis de K de 90 a 150 kg ha-1. En cambio, el rendimiento de fruto de tamaño 2 para exportación, no cambió significativamente entre esas dosis de N y K. No hubo diferencias entre dosis de N y K con relación al número de frutos por planta de calabacita en cualquiera de los dos tamaños de fruto, lo que indica que las diferencias en rendimiento se debieron a diferencias en el peso individual de los frutos. La tasa de asimilación de CO2 no fue afectada por las dosis de N y K evaluadas, pero dichas tasa fueron altas par una planta C, con valores de 41 y 38 μmoles de CO m-2 s-1, en dos fechas de medición, ya que en la mayoría de plantas C3 son tasas menores o iguales a 30 μmoles de CO2 m-2s-1.
- En cuanto a la calidad del fruto, el contenido de azúcares totales del fruto de tamaño 1 para consumo nacional, no fue afectado por los tratamientos de N y K, pero en los de tamaño 2 para exportación este valor aumentó al subir la dosis de N de 150 a 330 kg ha-1. La firmeza de los frutos fue más alta con el menor nivel de N y con el mayor de K, en ambos tamaños de fruto. En color del fruto, todos los tratamientos favorecen el color verde y el tratamiento que alcanzó el menor verdor fue el de la menor tasa de N (150 kg ha-1) y la intermedia de K (120 kg ha-1 de K).

