











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La ectomicorriza es una simbiosis que se establece entre alrededor de 2000 especies de plantas y 7000 especies de hongos. Dicha simbiosis resulta fundamental en la estructura y funcionamiento de ecosistemas boreales, templados y algunos tropicales (Pérez-Moreno y Read, 2004). Los hongos ectomicorrízicos (ECM), debido en parte a las conspicuas redes miceliales que establecen, incrementan la adquisición de agua y nutrientes de las plantas con las que se asocian y pueden llegar a constituir también un importante factor de control de patógenos vegetales (Smith y Read, 1997). Debido a esto, la inoculación con hongos ECM constituye una importante herramienta en la propagación masiva de plantas, dado que con frecuencia la ectomicorriza es obligada para los árboles con los que se asocia (Bruns et al., 2002; Castellano y Molina, 1989). Debido a: i) la compatibilidad simbiótica de hongos y plantas, ii) la adaptabilidad ecológica de los simbiontes fúngicos en diferentes sustratos y su habilidad para competir con hongos nativos, iii) los efectos benéficos diferenciales en las plantas hospederas y iv) la facilidad de producción de inóculo, es importante la selección de los simbiontes fúngicos apropiados en programas de inoculación con hongos ECM en vivero (Parladé et al., 2004; Rincón et al., 2001). Adicionalmente, un criterio que ha cobrado importancia a últimas fechas en la selección de hongos ECM a ser utilizados para inocular plantas es su comestibilidad. A nivel mundial, se han registrado más de 200 especies de hongos ECM comestibles con importancia económica. Algunos de ellos tienen un amplio mercado internacional cotizado en billones de dólares anualmente tales como trufas, matsutake, porcini, cantarelos y hongo Cesar (Wang y Hall, 2004) y adicionalmente, existen centenares de especies que son consumidas localmente en diferentes países. Por ejemplo, en México se conocen más de 200 hongos silvestres comestibles los cuales han sido consumidos por siglos (Villareal y Pérez-Moreno, 1989) y en la actualidad son comercializados por diversos grupos étnicos en mercados locales en centenares de toneladas anualmente (Pérez-Moreno et al., 2008, 2010). El uso de productos forestales no maderables, ha recibido recientemente atención como un factor de enorme importancia para la conservación forestal, por su importancia socioeconómica. Por ejemplo en México más de 50% de las especies comestibles conocidas son ectomicorrízicas y podrían potencialmente utilizarse en la inoculación de plantas. Desafortunadamente, los estudios relacionados con la inoculación de muchas de las especies comestibles de hongos ECM y su eficiencia en el crecimiento vegetal son incipientes. El objetivo del presente trabajo fue estudiar el efecto de la inoculación individual o combinada de tres especies de hongos comestibles ECM en plantas de Pinus greggii crecidas en suelos de dos procedencias, vaporizados o no.
Materiales y métodos
Material Biológico y Preparación de Inóculo
La semillas utilizadas de P. greggii proceden de áreas en el municipio Nicolás Romero, en el estado de México. Previo a su siembra, las semillas fueron esterilizadas con H2O2 30% por 20 minutos y lavadas con agua destilada estéril. Para seleccionar las especies de los hongos ECM comestibles a utilizarse, se evaluaron las especies de hongos ECM comercializadas en mercados ubicados en el área de influencia de los Parques Nacionales Izta-Popo y Zoquiapan (ubicados en la parte central de México entre 18° 59’ y 19° 16’ 25’’ N y 98º 34’ 54’’ y 98º 16’ 25’’ W) con mayor potencial de producción de inóculo. Como consecuencia, se seleccionaron tres especies: Hebeloma mesophaeum s.l., Laccaria laccata s.l. y Suillus aff. pseudobrevipes. En el caso de L. laccata y H. mesophaeum se utilizaron exclusivamente los píleos, y en el caso de S. aff. pseudobrevipes los himenios. La razón por la cual se utilizaron exclusivamente los himenios en el caso de S. aff. pseudobrevipes es que el contexto de los cuerpos fructíferos de esta especie se descompone con facilidad, lo que dificulta la preparación del inóculo con los píleos completos. Estas estructuras fueron deshidratadas a 36 °C como máximo, para mantener la viabilidad de los propágulos, molidas y pasadas a través de un tamiz de 1mm para homogeneizar el tamaño de partícula. El inóculo así obtenido fue almacenado a 5 °C hasta su utilización.
Características Físicas y Químicas de los Sustratos
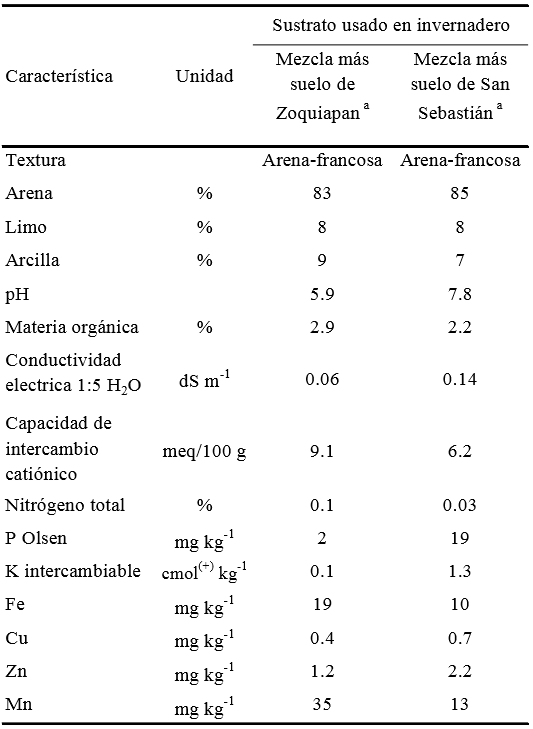
Se recolectaron dos suelos con características físicas y químicas distintas: i) un suelo forestal en el Parque Nacional Zoquiapan y ii) un suelo actualmente degradado donde existió bosque, procedente de la localidad de San Sebastián, Texcoco. Dichos suelos fueron utilizados como sustratos en el experimento en invernadero. Las características físicas y químicas de los sustratos se muestran en el Cuadro 1.
Montaje del Experimento
Los sustratos utilizados fueron tratados con vapor durante 4 h a una presión de 1.3 kg cm-2 y una temperatura de 125 °C. La siembra se efectuó en tubetes de plástico negro de 140 cm3 que contenían una mezcla de aserrín-arena-suelo en proporción 2:2:1. Cada planta fue inoculada con al menos 107 a 108 esporas. Con el inóculo esporal preparado con el procedimiento descrito anteriormente se efectuaron dos inoculaciones, con la finalidad de incrementar la probabilidad de colonización de las plantas. Una porción del inóculo ectomicorrízico a utilizarse fue colocada en una cavidad que se hizo en la superficie de los sustratos contenidos en los tubetes, 30 días después de la siembra. La otra porción del inóculo se incorporó 90 días después de la siembra efectuando una cavidad en la superficie de los sustratos. Para reducir las probabilidades de contaminación se colocó tezontle estéril en la superficie de los tubetes. Las plantas fueron regadas con agua destilada durante 240 días. Al inicio del riego, se aplicó captan en una solución que contenía 2 g por L de agua destilada.
Diseño Experimental y Análisis Estadístico
Se utilizó una distribución completamente al azar y un diseño factorial 5 × 2 × 2. Los tres factores involucrados fueron: inóculo, procedencia del suelo y vaporización del sustrato. El factor inóculo tuvo 5 niveles: i) plantas inoculadas con Hebeloma mesophaeum, ii) plantas inoculadas con Laccaria laccata, iii) plantas inoculadas con Suillus aff. pseudobrevipes, iv) plantas inoculadas con la combinación de los tres hongos y v) plantas sin inocular. El factor procedencia del suelo tuvo dos niveles: i) suelo procedente de Zoquiapan y ii) suelo procedente de San Sebastián, Texcoco. El factor vaporización tuvo dos niveles: i) suelo vaporizado y ii) suelo no vaporizado. Este diseño experimental, entonces, constó de 20 tratamientos, cada uno de los cuales tuvo 5 réplicas lo que originó 100 unidades experimentales. Cada unidad experimental estuvo constituida por una planta. A los datos que se obtuvieron en las diferentes variables evaluadas se les realizaron análisis de varianza y pruebas de comparación de medias de Tukey (α = 0.05) (SAS Institute, 1999). En el caso de los valores de colonización micorrízica que se expresan como porcentajes, los datos se transformaron y se utilizaron sus arcosenos en los análisis de varianza.
Variables Evaluadas
Se efectuaron mediciones periódicas de la altura del tallo de cada planta a los 90, 180 y 240 días después de la siembra. Se evaluó el peso seco de la parte aérea y el radical 240 días después de siembra. Las plantas fueron previamente secadas a 80 °C. Al final del experimento también se determinó el N total de la parte aérea y radical de cada planta, según Bremner (1965) y se evaluó el número de raíces cortas totales y el porcentaje de colonización ectomicorrízica en todas las plantas según el método propuesto por Marx et al. (1994). Se efectuó una caracterización morfológica de las ectomicorrizas encontradas (Agerer, 1991; Deemy, 2008) para evaluar la colonización por los hongos inoculados.
Resultados y discusión
Altura y Peso Seco de las Plantas
La procedencia de los suelos y la inoculación con hongos ECM afectaron significativamente (P < 0.001) todas las variables evaluadas relacionadas con crecimiento vegetal, contenido nutrimental y colonización ectomicorrízica de P. greggii. En contraste, la vaporización del suelo afectó solo la altura y el contenido de N radical (Cuadro 2). En la interacción entre vaporización del suelo e inoculación con hongos ECM se presentan diferencias significativas (P < 0.05) para todas las variables, excepto la colonización ectomicorrízica total. Un fenómeno similar se observó en el caso de la interacción entre procedencia del suelo e inoculación con hongos ECM. En contraste, en la interacción entre procedencia del suelo y vaporización no se observaron diferencias significativas para la mayoría de las variables evaluadas (Cuadro 2).
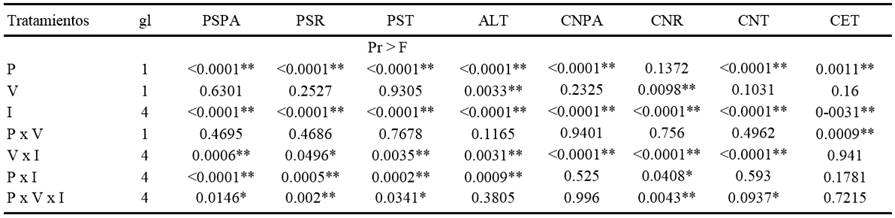
P = procedencia de suelo (San Sebastián y Zoquiapan) , V = vaporización del suelo (suelo vaporizado o no vaporizado), I = inoculación con hongos ectomicorrízicos (plantas sin inocular, plantas inoculadas con Hebeloma mesophaeum, plantas inoculadas con Laccaria laccata, plantas inoculadas con Suillus aff.. pseudobrevipes, plantas inoculadas con los tres hongos), PSPA = peso seco de la parte aérea, PSR = peso seco de la raíz, PST = peso seco total, ALT = altura de las plantas, CNPA = contenido de N de la parte aérea, CNR = contenido de N de la raíz, CNT = contenido de N total de la planta, y CET = colonización ectomicorrízica total. * significativo P ≤ 0.09, ** altamente significativo, P ≤ 0.01.
Cuadro 2. Prueba F de las variables de Pinus greggii en los tratamientos de procedencia de suelos, vaporización e inoculación ectomicorrízica en invernadero.
En términos generales, la inoculación con hongos ECM no produjo diferencias significativas en la altura del tallo de las plantas inoculadas en comparación a las plantas no inoculadas 90 días después de la siembra. Sin embargo, los efectos benéficos reportados al final del experimento, en términos de altura de las plantas (Cuadro 3), fueron evidentes y mostraron diferencias significativas 180 días después de la siembra.
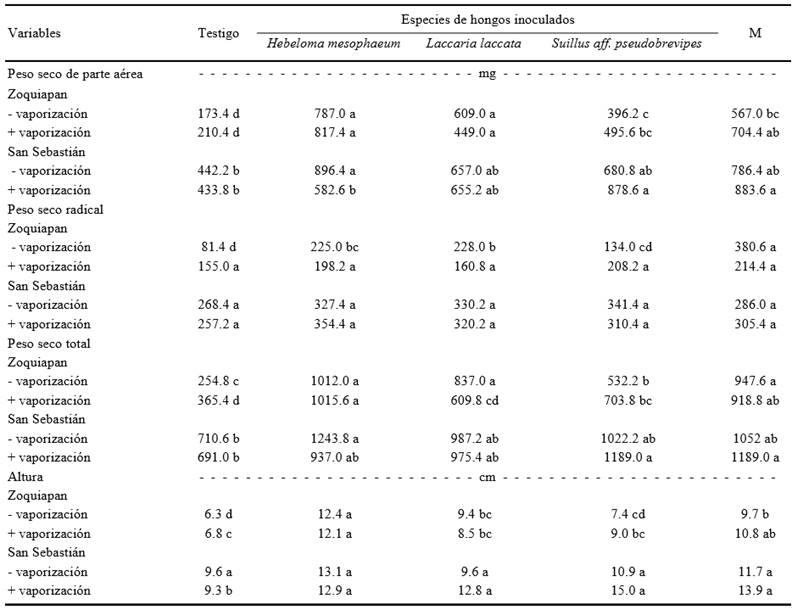
- vaporización = sustrato no tratado con vapor, de acuerdo a como se describe en materiales y métodos; + vaporización = sustrato tratado con vapor como se describe en materiales y métodos. M = Mezcla de H. mesophaeum, L. laccata y S. aff. pseudobrevipes. Valores en la misma línea con la misma letra son iguales según Tukey (P ≤ 0.05). n = 5.
Cuadro 3. Peso seco, diámetro del tallo y altura de plantas de Pinus greggii inoculadas o no con hongos ectomicorrízicos 240 días después de la siembra.
La respuesta de crecimiento vegetal como resultado de la inoculación fúngica dependió del tipo de hongo ECM inoculado. Cuando se inoculó con H. mesophaeum se observó un mayor peso seco total (P = 0.05) independientemente de la procedencia del suelo y de la condición de vaporización. El mayor incremento en peso seco total se observó cuando se utilizó suelo procedente de Zoquiapan sin vaporizar. En este caso, las plantas inoculadas con H. mesophaeum produjeron 3.9 más peso seco total comparadas con las plantas no inoculadas. Cuando se inoculó con S. aff. pseudobrevipes, se observó también un mayor peso seco total en comparación con las plantas no inoculadas. La inoculación con L. laccata solamente produjo mayor peso seco total cuando se utilizó suelo procedente de Zoquiapan sin vaporizar. La inoculación simultánea con las tres especies de hongos ECM evaluados no originó un efecto sinérgico, tampoco se observó un efecto de competencia entre las especies inoculadas (Cuadro 3). El peso seco total de las plantas de P. greggii que fueron inoculadas simultáneamente con las tres especies, fue similar al de las plantas inoculadas solo con H. mesophaeum y siempre superior al de las plantas sin inocular (Cuadro 3).
Previamente, se han documentado incrementos en la biomasa vegetal como resultado de la inoculación con algunos de los géneros de hongos ECM estudiados en el presente trabajo. Por ejemplo, Sudhakara-Reddy y Natarajan (1997) reportaron incrementos de 218% y 182% en el peso seco de la parte aérea y radical, respectivamente como resultado de la inoculación con L. laccata en relación a plantas no inoculadas de Pinus patula, doce meses después de la inoculación en suelo esterilizado. Diversas investigaciones han reportado que la inoculación múltiple con distintos de hongos ECM puede producir mayor incremento en biomasa en las plantas inoculadas que la inoculación individual (Parladé y Álvarez, 1993; Sudhakara-Reddy y Natarajan, 1997). Sudhakara-Reddy y Natarajan (1997) inocularon simultáneamente L. laccata y Telephora terrestris en Pinus patula. Dichos autores encontraron un efecto sinérgico en el peso seco de la parte área como resultado de la inoculación múltiple con dichas especies. Sin embargo, en nuestro caso la inoculación simultánea con las tres especies de hongos ECM estudiados no produjo un efecto sinérgico, a pesar de que se mantuvo, en términos generales un efecto benéfico. Un fenómeno similar ha sido encontrado por otros autores como Chu-Chou y Grace (1985). Estos autores inocularon L. laccata, Rhizopogon luteolus y R. rubescens individualmente o de manera combinada en Pinus radiata. A pesar de que existió un efecto en crecimiento en las plantas inoculadas con cualquiera de las especies de hongos ECM cuando fueron aplicadas de manera individual en relación a las plantas no inoculadas, no existió un efecto sinérgico al efectuar una inoculación con las tres especies fúngicas simultáneamente. Estos incrementos en crecimiento variaron de 109 a 397% dependiendo de las especies vegetales y fúngicas involucradas.
En el presente estudio se observó que, en términos generales, en los primeros dos meses la inoculación con los hongos ECM redujo el crecimiento de las plantas inoculadas en comparación a las plantas no inoculadas (datos no mostrados) y posteriormente se observaron paulatinamente los efectos benéficos reportados al final del experimento, los cuales fueron más evidentes y mostraron diferencias significativas a partir del sexto mes. Previamente, Sudhakara-Reddy y Natarajan (1997) al inocular Pinus patula con L. laccata encontraron que los efectos benéficos, en términos de peso seco de parte aérea y radical, sólo fueron evidentes a partir del octavo mes después de la inoculación. Este fenómeno probablemente puede ser explicado en términos del alto costo inicial que requiere el establecimiento de la simbiosis ectomicorrízica para la planta, en términos del C fijado. Se ha reportado que el costo de la simbiosis ectomicorrízica puede constituir de 7 a 30% del C fijado por las plantas (Finlay y Söderström, 1992; Ek, 1997; Bidartondo et al., 2001). En un inicio este puede ser considerado un alto costo para las plantas, sin embargo, debido a que una vez establecidas las redes miceliales existe la posibilidad de traslocación substancial de N y P (Pérez-Moreno y Read, 2000; 2001a,b) la simbiosis ECM en términos generales es considerada mutualista (Read y Pérez-Moreno, 2003).
Al momento de la cosecha, no existieron conspicuas diferencias entre las plantas crecidas en el sustrato tratado con vapor comparadas con aquellas crecidas en sustrato que no fue tratado con vapor. Sin embargo, diez meses después de la inoculación (datos no mostrados) un juego de plantas paralelas dejadas en invernadero mostraron diferencias entre ambos tipos de sustratos, observándose un mayor crecimiento en las plantas crecidas en el sustrato tratado con vapor. Sudhakara-Reddy and Natarajan (1997) comparando resultados de suelo esterilizado y no esterilizado, encontraron que el número total de ectomicorrizas y peso de las plantas de Pinus patula inoculadas con L. laccata y Thelephora terrestris fue superior en suelo esterilizado que en suelo no esterilizado, lo cual puede ser explicado por la falta de competencia con microorganismos nativos del suelo. Guerin-Laguette et al. (2003) reportaron que la altura y peso seco de Pinus sylvestris inoculadas con el hongo ECM L. deliciosus, produjeron mayor peso seco y altura cuando crecieron en sustrato esterilizado comparadas con aquellas plantas crecidas en sustrato no estéril.
Contenido de N
El contenido total de N varió de 1.50 a 16.7 mg por planta, de acuerdo a los hongos inoculados y las procedencias de los suelos. De manera similar que para el peso seco total y la altura, el contenido total de N fue siempre superior en las plantas inoculadas con hongos ECM que en las no inoculadas. Este fenómeno se observó independientemente de la especie de hongo inoculada y de la procedencia del suelo empleado (Cuadro 4). Los mayores contrastes en contenido de N se observaron entre las plantas inoculadas con H. mesophaeum y aquellas no inoculadas.
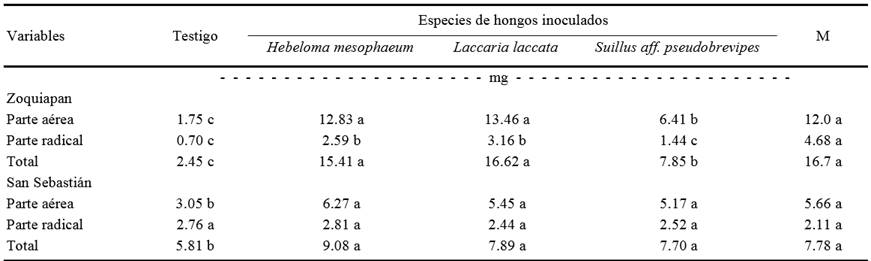
M = mezcla de H. mesophaeum, L. laccata y S. aff. pseudobrevipes. Valores en la misma línea con la misma letra son iguales según Tukey (P ≤ 0.05). n = 5.
Cuadro 4. Contenido de nitrógeno de parte aérea, radical y total de plantas de Pinus greggii inoculadas o no con hongos ectomicorrízicos 240 días después de la siembra.
Diversos autores han reportado que la inoculación con hongos ECM origina incremento en la concentración o contenido total de nutrimentos de árboles y arbustos templados y tropicales, principalmente N (Turjaman et al., 2006; Nara, 2006; Tibbett y Sanders, 2002; Baxter y Dighton, 2001). En nuestro caso, se observó que los altos niveles de colonización ECM de las especies inoculadas estuvieron relacionadas, en términos generales, con incrementos en las concentraciones de N.
Colonización Ectomicorrízica
Al final del experimento, en términos generales, la colonización micorrízica fue superior en las plantas inoculadas con hongos ectomicorrízicos comestibles comparados con plantas no inoculadas independientemente de la procedencia del suelo utilizado en el sustrato y de la condición de vaporización. Los porcentajes de colonización ectomicorrízica correspondiente al hongo inoculado alcanzaron hasta 77% (Cuadro 5), lo cual es un indicador de que las especies inoculadas se establecieron extensivamente en las raíces de los pinos evaluados.
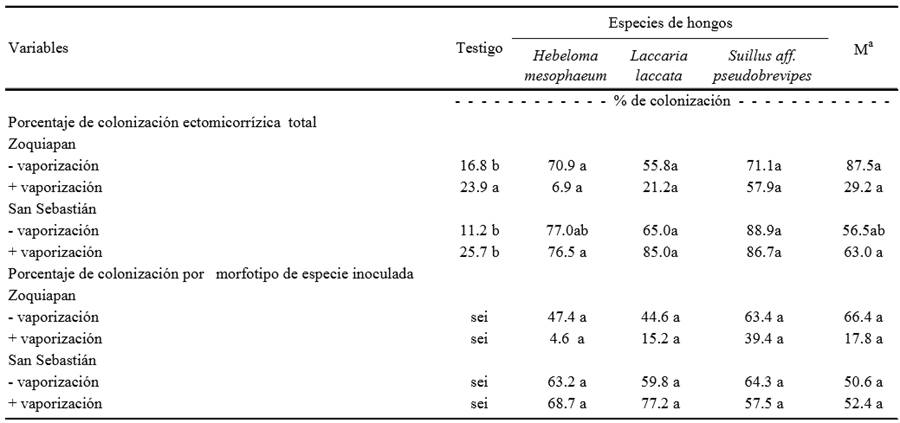
- vaporización = sustrato no tratado con vapor, de acuerdo a como se describe en materiales y métodos; + vaporización = sustrato tratado con vapor. M = Mezcla de H. mesophaeum, L. laccata y S. aff. pseudobrevipes. sei = sin especie inoculada; a cuando se inoculó con tres especies fúngicas se muestran solo los datos de la ectomicorriza de la especie dominante. Valores en la misma línea con la misma letra son iguales según Tukey (P = 0.05). n = 5.
Cuadro 5. Porcentajes de colonización ectomicorrízica total y por morfotipo de especies inoculadas en raíces de plantas de Pinus greggii 240 días después de la siembra.
Colonizaciones más altas a las encontradas en el presente trabajo han sido reportadas por otros autores, por ejemplo, Jonson et al. (2001) registraron colonizaciones de 55 a 78% y de 95 a 99% en Pinus sylvestris inoculados con Hebeloma crustiliniforme y Laccaria bicolor, respectivamente. Rincón et al. (2001) reportaron colonizaciones superiores a 80% al inocular plantas de Pinus pinea con H. crustiliniforme. Otros autores han reportado colonizaciones menores a las observadas en nuestro caso, por ejemplo, Sudhakara-Reddy y Natarajan (1997) encontraron colonizaciones de alrededor de 60% al inocular Pinus patula con Laccaria lacata y Rincón et al. (2001) observaron colonizaciones de 11 al 40% al inocular Pinus pinea con Laccaria lacata. Trocha et al. (2007) reportan que H. mesophaeum coloniza del 18 al 22% de las raíces de Betula pendula creciendo en paredes de ladrillo.
Hasta donde conocemos, ésta es la primera ocasión en la que se evalúa el efecto de la inoculación de hongos ECM en plantas de P. greggii. México constituye un centro de diversidad de especies de árboles ECM, por ejemplo el país posee el mayor número de especies y endemismo de Pinus y Quercus en el mundo, con 72 y 161 taxa y especies respectivamente (Perry, 1991; Valencia, 2004). Sin embargo, una gran cantidad de estos taxa no han sido estudiados desde el punto de vista de sus asociaciones simbióticas ECM. En el presente estudio se observó que la respuesta vegetal varió dependiendo de la especie de hongo inoculada, por lo que debido a la gran diversidad de pinos existentes en México, sería también altamente deseable el desarrollo estudios con otras de pinos mexicanos de importancia forestal.
Conclusiones
Las plantas de pino inoculadas con cualquiera de las tres especies de hongos ECM evaluadas, solas o en combinación, tuvieron mayor biomasa comparadas con plantas no inoculadas. El contenido de N total fue siempre superior en las plantas inoculadas con hongos ECM que en las plantas no inoculadas. La vaporización de los sustratos no tuvo efecto en las variables de crecimiento evaluadas y en el contenido de N de P. greggii. Los altos porcentajes de colonización ectomicorrízica correspondientes morfológicamente a los hongos inoculados son un indicador de que las especies inoculadas se establecieron extensivamente en las raíces de los pinos evaluados. Dado que las 3 especies de hongos estudiados son objeto de comercialización masiva en mercados mexicanos constituyen una importante fuente potencial de inóculo ectomicorrízico para ser utilizado en viveros mexicanos.

