










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkTerra Latinoamericana
versión On-line ISSN 2395-8030versión impresa ISSN 0187-5779
Terra Latinoam vol.28 no.2 Chapingo abr./jun. 2010
División II
Concentración de cationes en frijol común en respuesta a aluminio
Cations concentration in common bean in response to aluminum
Rodolfo Landa Contreras1, Libia I. Trejo Téllez1*, Fernando C. Gómez Merino1, Bernardo Villar Sánchez2 y Cecilia García Osorio1
1 Colegio de Postgraduados, Campus Montecillo. 56230 Montecillo, estado de México. *Autor responsable: tlibia@colpos.mx.
2 INIFAP, Campo experimental Centro Chiapas. Km 3 Carretera Internacional. Ocozocoautla de Espinoza. 29140 Chiapas, México.
Recibido: abril de 2008.
Aceptado: julio de 2009.
RESUMEN
En México, diversas variedades de frijol (Phaseolus vulgaris L.), son cultivadas en suelos ácidos con altos contenidos de aluminio soluble (Al3+), sin que haya sido evaluada su tolerancia a este elemento. En esta investigación se estudió el efecto de 3 niveles de Al3+ (0, 50 y 100 μM) en la solución nutritiva, sobre la concentración de los cationes potasio, calcio y magnesio (K+, Ca2+ y Mg2+) in planta, en 3 variedades mexicanas de frijol negro (Jamapa, Medellín y Grijalva). Después de 23 días de tratamiento, las plantas fueron divididas en hojas, tallos y raíces y se determinó la concentración de los cationes en estos órganos. El factor concentración de Al3+ afectó significativamente la concentración de los cationes en hojas, tallos y raíces. El factor variedad no mostró efecto sobre la concentración de Ca2+ en los órganos evaluados, mientras que la concentración de K+ y Mg2+ si mostraron diferencias estadísticas debidas al factor variedad. La interacción de los factores en estudio ocasionó diferencias estadísticas significativas en la concentración de Ca2+ en los 3 órganos evaluados. La presencia de Al3+ en la solución nutritiva redujo de manera considerable la concentración de K+ en tallos y raíces en las 3 variedades, siendo más evidente en la variedad Medellín. La concentración de Ca2+ mostró tendencia similar a la de K+ en presencia de Al3+ en las variedades Jamapa y Medellín; por el contrario, la concentración de este elemento fue incrementada en hojas de la variedad Grijalva cuando fue tratada con 100 μM de Al3+. El Mg2+ no disminuyó su concentración en tejidos por la adición de Al3+; contrariamente, en la variedad Grijalva su contenido en hoja en el tratamiento con 100 μM Al3+ superó a la del testigo en 23%. Los resultados obtenidos permiten concluir que la variedad Grijalva presentó menor susceptibilidad a Al3+ y la Medellín fue menos tolerante a este elemento.
Palabras clave: Phaseolus vulgaris, toxicidad por aluminio, potasio, calcio, magnesio.
SUMMARY
In Mexico, different varieties of common bean (Phaseolus vulgaris L.) are cultivated on acid soils with high contents of soluble aluminum (Al3+), but these varieties have not been evaluated for their tolerance to Al. Therefore, in this research the effect of three concentrations of Al3+ (0, 50 and 100 μM) in the nutrient solution on the concentration of potassium, calcium and magnesium (K+, Ca2+ and Mg2+) cations in planta, in 3 Mexican varieties of common bean (Jamapa, Medellín and Grijalva) was assessed. After 23 days of Al treatment, plants were harvested and divided into leaves, stems and roots, and cation concentrations contained in each organ was determined. Concentration of Al3+ significantly affected cation concentration in leaves, stems and roots. The factor variety did not affect Ca2+ concentration in any plant organ studied, while K+ and Mg2+ concentrations showed significant differences due to the factor variety. Interaction between factors significantly affected Ca2+ in the 3 organs evaluated. Al3+ concentration drastically reduced concentration of K+ in both stems and roots, which was more notable in Medellín. Ca2+ concentration showed a similar tendency as that observed in K+ in the presence of Al in both varieties Jamapa and Medellín; conversely, Ca2+ concentration increased in leaves of the variety Grijalva when it was treated with 100 μM de Al3+. Mg2+ concentration did not decrease in any organ as an effect of Al; instead, in the variety Grijalva its concentration in leaves of plants treated with 100 μM Al3+ was 23% higher than that showed in control plants. We conclude that the variety Grijalva was less affected by Al3+ concentration in the nutrient solution and that the variety Medellín was less tolerant to Al.
Keywords: Phaseolus vulgaris, aluminum toxicity, potassium, calcium, magnesium.
INTRODUCCIÓN
El aluminio (Al) es considerado uno de los factores que reduce el crecimiento y rendimiento de plantas desarrolladas en suelos ácidos con valores de pH menores a 5 (Samac y Tesfaye, 2003). Se calcula que un 40% de la superficie cultivable del planeta y aproximadamente el 70% del suelo utilizado para la producción de alimentos son suelos ácidos y, por tanto, sujetos a la toxicidad por este metal (Sumner y Noble, 2003). Poscherieder et al. (1992) establecieron que la toxicidad por aluminio (Al3+) reduce de manera significativa la absorción y translocación de elementos esenciales para el desarrollo de la planta, principalmente cationes como el potasio (K+), calcio (Ca2+) y magnesio (Mg2+), lo que repercute sobre el rendimiento. Liu y Luan (2001) mencionan que la inhibición de la absorción de K+ por presencia de Al3+ en Arabidopsis thaliana, es causada por un bloqueo de la superficie interna de la membrana plasmática, lo que conduce a la disminución del 50% de la absorción de este elemento cuando las plantas son tratadas con una concentración de 10 μM de Al3+; una concentración de 50 μM Al3+, la reduce en un 79%. En lo que a Ca2+ respecta, se ha demostrado que el ión Al3+ lo desplaza en lugares de unión en el citoplasma. El desplazamiento de Ca2+ y substitución de éste por Al3+ en la pared celular puede ser suficiente para producir alteraciones en el desarrollo normal de la célula (Schofield et al., 1998).
Se ha reportado que especies de plantas tanto monocotiledóneas como dicotilédoneas presentan diferentes concentraciones de Mg2+ en tejido cuando crecen en condiciones de estrés por Al3+ (Keltjens y Tan, 1993). En vástagos de avena, Grimme (1983) reportó disminución del 30% en la concentración de Mg2+ cuando las plantas fueron tratadas con aluminio, en comparación con las plantas testigo; de la misma manera, en plántulas de abeto rojo (Picea abies Karst.) se observó una fuerte inhibición en la absorción y translocación de Mg2+ cuando fueron tratadas con 1 mM de Al3+ en la solución nutritiva (van Praag et al., 1997). El paso inicial en la interacción Al3+-Mg2+ toma lugar probablemente en el apoplasto radical. En ausencia de Al3+, altas cantidades de Mg2+ se unen a los sitios de intercambio en las paredes corticales de la raíz, conocidos como sitios de intercambio catiónico de la raíz. Como consecuencia, la concentración de Mg2+ incrementa en los alrededores del lado exterior del plasmalema. Cuando existen niveles externos altos de Al3+, una gran parte del Mg2+ unido en el complejo de intercambio de la raíz es desplazado por estos iones, lo que conduce a la deficiencia de Mg2+ (Marschner, 1995). Además, el ión Al3+ puede interferir en procesos fisiológicos en los que participa el Mg2+ debido a que su similitud en radio iónico es una característica aún más importante que su diferencia en cargas dentro de reacciones biológicas (da Silva et al., 2005).
Debido a que el frijol común (Phaseolus vulgaris L.) es una de las especies más cultivadas en zonas tropicales de México con alta incidencia de suelos ácidos y toxicidad por Al3+, y no existen reportes sobre las respuestas de variedades mexicanas a este elemento, el objetivo del presente trabajo fue evaluar el efecto del Al3+ (en 3 concentraciones, adicionadas a la solución nutritiva) sobre la concentración de K+, Ca2+ y Mg2+ en hojas, tallos y raíces de 3 variedades mexicanas de frijol negro.
MATERIALES Y MÉTODOS
Localización geográfica del área experimental
El presente trabajo se realizó en el invernadero del área de Nutrición Vegetal del Colegio Postgraduados, en Montecillo, estado de México, localizado a 19° 29' LN, 98° 53 ' LO y una altitud de 2250 m. El clima es clasificado como C(w0)(w)b(i')g, con temperatura media anual de 15 °C y una oscilación térmica de 5.8 °C (García, 1988).
Descripción del material vegetal
El criterio de selección de las 3 variedades de frijol empleadas fue su adaptación a climas tropicales, donde de manera predominante se encuentran suelos ácidos, y en consecuencia, concentraciones tóxicas de Al3+ en nuestro país. Las 3 variedades seleccionadas (Jamapa, Medellín y Grijalva) son de raza mesoamericana y el color de su testa es negro.
Experimento y diseño de tratamientos
En este experimento se consideraron 2 factores de estudio: a) variedad de frijol y b) concentración de Al3+ en la solución nutritiva. Cada uno de estos factores fue ensayado en 3 niveles: Jamapa, Grijalva y Medellín para las variedades y 0, 50 y 100 μM para el factor concentración de aluminio, lo que generó 9 tratamientos; los tratamientos fueron distribuidos completamente al azar. Una planta fue considerada como unidad experimental, teniéndose 8 plantas por tratamiento, las cuales fueron establecidas en 2 macetas de 3 L de capacidad (4 plantas por maceta). Con los datos obtenidos se realizó un análisis de varianza usando el programa de análisis estadístico SAS (SAS, 1985), empleando el procedimiento ANOVA y una prueba de comparación de media por Tukey (P • 0.05 y P • 0.01).
Manejo del cultivo
Las semillas de las variedades de frijol fueron sembradas en charolas de plástico conteniendo aproximadamente 30 cm3 de agrolita. Las plantas fueron regadas con agua destilada hasta la aparición de las hojas cotiledonares. Los 10 días posteriores fueron regadas con la solución de Hoagland y Arnon (Hoagland y Arnon, 1950) a 1/3 de concentración.
Establecimiento de plántulas en hidroponía
Plántulas de 10 días de edad y de una altura promedio de 9 cm fueron establecidas en macetas de 3 L de capacidad, sostenidas con hule espuma en los orificios de las tapas de las macetas. A la solución nutritiva empleada (Cuadro 1), se le adicionó una mezcla de micronutrimentos (Cuadro 2), en la cual Mn, Cu y Zn fueron suministrados en forma de sulfatos; B como H3BO3; Fe se suministró como quelato (Fe-EDTA) según lo descrito por Steiner y van Winden (1970). Las plantas fueron mantenidas bajo estas condiciones por 5 días (fase de aclimatación), para posteriormente dar inicio con los tratamientos (adición del Al+3 a la solución nutritiva en cada una de las concentraciones a evaluar). El aluminio fue abastecido a las soluciones nutritivas en forma de cloruro de aluminio hexahidratado (AlCl3 6H2O). El pH de las soluciones fue ajustado a 4.2 para garantizar que el Al permaneciera en su forma iónica Al3+. La solución nutritiva fue renovada cada 10 días, mientras que el pH se ajustó cada tercer día.
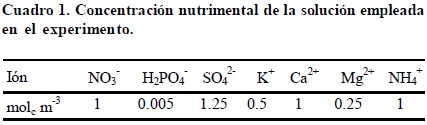
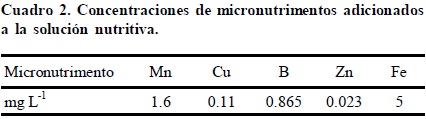
Concentración nutrimental en tejido vegetal
Después de 23 días de tratamiento, las plantas fueron divididas en hojas, tallos y raíces; posteriormente las muestras fueron secadas en una estufa de aire forzado (48 h, 65 °C). Después del secado de las muestras, éstas se molieron usando un molino tipo Wiley de acero inoxidable provisto de un tamiz malla 40. Las concentraciones de K+, Ca2+ y Mg2+ fueron determinadas mediante digestión húmeda del material seco con una mezcla de ácidos perclórico y nítrico (Alcántar y Sandoval, 1999). Posteriormente los extractos obtenidos se leyeron en un equipo de espectrometría de emisión atómica de inducción con plasma acoplado en un equipo ICP-AES VARIAN TM modelo Liberty II.
RESULTADOS Y DISCUSIÓN
Concentración de Potasio
La significancia estadística de las concentraciones de K+ en los órganos evaluados en función de la concentración de Al3+ en la solución nutritiva se presenta en el Cuadro 3. El factor concentración de Al3+ ocasionó diferencias estadísticamente significativas (P • 0.05) sobre la concentración de este elemento en hojas y altamente significativas en tallos y raíces (P • 0.01). El factor variedad ocasionó únicamente diferencias estadísticas significativas en la concentración de K+ en tallo (P • 0.05); en este órgano la interacción de factores ocasionó diferencias estadísticas altamente significativas (P • 0.01).
Las concentraciones de K+ obtenidas en hojas, tallos y raíces de las 3 variedades de frijol sometidas a diferentes concentraciones de Al3+ en la solución nutritiva son mostradas en la Figura 1. Las concentraciones promedio de este elemento en las 3 variedades de frijol en el tratamiento testigo fueron de 27.9, 28.1 y 34.4 g kg-1 para hojas, tallos y raíces, respectivamente. Estas concentraciones son similares a las reportados por Turna et al. (2004) para frijol en hojas (24 g kg1) y en tallos (32 g kg-1); además de coincidir en que la concentración de K en frijol es mayor en tallos que en hojas, lo anterior como respuesta a la función fisiológica de este nutrimento en los procesos de transporte en plantas.
En la Figura 1 también se observa que la variedad Medellín mostró mayor susceptibilidad a la presencia de aluminio. El tratamiento con 100 μM Al3+ redujo la concentración de K+ en casi 17.5% en hojas, en comparación con el testigo; en tallos dicha reducción fue de 44.7% (Figura 1a y b).
La variedad Jamapa mostró resultados similares a la variedad Medellín. La concentración de K+ en raíces (Figura 1c), se redujo considerablemente por la presencia de Al3+ en la solución nutritiva (26 y 46% para las soluciones conteniendo 50 y 100 μM Al3+, respectivamente). En esta variedad, los tallos de plantas tratadas con 100 μM de Al3+, mostraron reducción en 47% en la concentración de este catión, en comparación con los tallos de plantas testigo. En forma contraria a los resultados de tallo y raíz, la variedad Jamapa presentó la menor reducción en la concentración de K+ por efecto de la concentración de Al3+ en la solución nutritiva, siendo sólo 1.15% menor en el tratamiento de 100 μM de Al3+ a la concentración registrada en las plantas que no fueron tratadas con Al3+. Asimismo, en esta variedad la adición de 50 μM Al3+ incrementó la concentración de este catión en 3.47% en hojas, en comparación con el testigo.
Es importante destacar que en la variedad Grijalva la mayor concentración de Al3+ en la solución nutritiva incrementó ligeramente la concentración de K en raíces (1.7% mayor a la del testigo).
El potasio es el catión más abundante en la célula vegetal y presenta una alta movilidad dentro de la planta (Fox y Guerinot, 1998). Incrementos en la concentración de K+ en hojas y raíces por la presencia de Al3+, como en el caso de las variedades Jamapa y Grijalva, han sido previamente reportados en Trifolium repens L. (Lee y Pritchard, 1984), y en raíces de arroz (Tanoi et al., 2006). Estos últimos resultados permitieron concluir que la absorción de K no ocurre por procesos de difusión dentro de las células, sino por transporte activo mediado por transportadores tales como el transportador H+:K+ o por gradiente de protones (H+), el cual es promovido por la presencia de Al3+. Por el contrario, las reducciones en la concentración de K+ a nivel celular, son debidas según Liu y Luan (2001), a la inhibición de la entrada de este elemento causada por el bloqueo de canales de absorción de K localizados en la membrana plasmática interna (Kin) provocada por el Al3+, cuando éste se encuentra en el citosol.
Concentración de Calcio
El Cuadro 4 muestra la significancia estadística de los factores de estudio y la interacción de éstos sobre la concentración de Ca2+ en hojas, tallos y raíces de frijol. La variedad no influyó sobre la concentración de Ca2+ en los órganos evaluados (P > 0.05). Por el contrario, la concentración de Al3+ y la interacción de ésta con el factor variedad ocasionó diferencias altamente significativas (P • 0.01) en la concentración de Ca2+ en hojas, tallos y raíces.
El Ca se encuentra en tejido foliar en concentraciones menores a las del K, las cuales oscilan entre 0.1 y 2% de la materia seca (Fox y Guerinot, 1998). En el caso particular de las variedades de frijol evaluadas en esta investigación, los contenidos promedio registrados de este nutrimento en el testigo fueron de 22.5 g kg-1 tanto para hojas como para tallos, y 6 g kg-1 para raíces (Figura 2).
Entre variedades se observan efectos distintos causados por la presencia de Al3+ sobre la concentración de Ca2+. Es evidente por un lado, la disminución de la concentración de Ca en las variedades Jamapa y Medellín al ser tratadas con Al (Figura 2), respuesta debida a los trastornos ocasionados por el bloqueo de canales en la membrana externa de la célula causados por el Al3+; se argumenta también que el Al es capaz de bloquear los canales ubicados en la membrana interna (Rout et al., 2001). Jones et al. (1998) reportaron que la concentración de Ca en el citosol disminuyó significativamente en células de tabaco después de ser tratadas con Al. Además, se ha reportado que plantas en presencia de Al3+ presentan menor concentración de Ca que aquellas deficientes en este nutrimento (Rengel y Elliot, 1992). En forma contraria a las reducciones en la concentración de Ca2+ como consecuencia del tratamiento con Al3+ anteriormente mencionadas, en la variedad Grijalva el tratamiento con 100 μM de Al3+ incrementó la concentración de este catión en hojas y tallos; lo que conduce a afirmar la existencia de mecanismos intrínsecos de cada variedad para recuperarse del estrés por Al, como lo indican Watanabe y Okada (2005), quienes demostraron que la tolerancia no solo a Al, sino también a estroncio (Sr) y bario (Ba) en arroz, exhibe amplias diferencias entre cultivares. Los mecanismos de tolerancia a Al que hasta ahora han sido documentados en plantas son muy variados. Existen evidencias experimentales que indican que plantas monocotiledóneas y varias especies de plantas dicotiledóneas excluyen la entrada de A13+ a las células de la raíz a través de la exudación de ácidos orgánicos como citrato, malato y oxalato, los cuales inmovilizan al Al3+ en la superficie de la raíz formando complejos estables no tóxicos con éste (Delhaize et al., 1993; Kollmeier et al., 2001, Liu et al., 2007). Kochian et al. (2004) afirman que algunas especies de plantas poseen canales selectivos de cationes que no permiten la entrada de Al a la célula o bien que impiden el transporte de este elemento a la parte aérea; en consecuencia, en estas plantas no se inhibe la absorción de cationes.
Concentración de Magnesio
El factor concentración de Al3+ y la interacción de éste con el factor variedad afectaron en forma altamente significativa (P • 0.01) la concentración de Mg2+ en hojas. Esta misma respuesta se presentó en tallos, en donde adicionalmente existió efecto significativo de la variedad (P • 0.05). Ambos factores (concentración de Al3+ y variedad) tuvieron efecto altamente significativo sobre la concentración de Mg2+ en raíces, pero no sus interacciones (Cuadro 5).
La Figura 3 presenta los resultados obtenidos de concentración de Mg2+ en hojas, tallos y raíces de 3 variedades de frijol tratadas con niveles distintos de Al3+. Las concentraciones de Mg reportadas por Tûma et al. (2004) en tejido foliar de frijol es de 6.4 g kg1 y en tallo de 4.2 g kg-1. Estos resultados coinciden sólo con los encontrados en hojas en esta investigación, con una concentración promedio (considerando las 3 variedades) en el tratamiento sin aluminio de 6.2 g kg-1. En el caso de tallo, la concentración de Mg2+ registrada en este experimento fue similar a la de hojas (6.2 g kg-1), y por lo tanto superior a la reportada por Tûma et al. (2004). Las concentraciones de este nutrimento fueron menores a las registradas para K+ y Ca2+.
Las variedades evaluadas mostraron respuestas diferenciales a la presencia de Al3+, si se considera la concentración de Mg2+. En las variedades Grijalva y Medellín, se registraron incrementos en la concentración de este catión en hojas, cuando las plantas fueron tratadas con 100 μM Al3+, éstos son del orden del 23.4 y 19.8%, respectivamente, en comparación con el testigo; por el contrario, la variedad Jamapa, redujo en el mismo tratamiento la concentración de este elemento en hojas en casi 31.5%.
La variedad Grijalva mostró una baja reducción en la concentración de Mg2+ en el tallo cuando fue tratada con 50 μM de Al3+ (13.1% menor a la concentración registrada en las plantas testigo), y con la mayor concentración de Al3+ (100 μM), la concentración de este catión fue superior en 26% a la de los tallos de plantas que no fueron tratadas con aluminio.
En raíces la presencia de aluminio, independientemente de su concentración, disminuyó la concentración de Mg2+; en el tratamiento con la mayor concentración de Al3+ evaluada la concentración de Mg2+ fue menor en 55, 21 y 24.8% en comparación con el testigo en las variedades Grijalva, Jamapa y Medellín, respectivamente.
Estos resultados permiten aseverar que las concentraciones de Mg2+ mostradas en hoja presentan una relación inversa con las concentraciones en raíces cuando las plantas fueron tratadas con la mayor concentración de Al3+; lo anterior, conduce a afirmar que la translocación de este elemento a hojas no fue afectada por la presencia de Al3+. En este sentido, se ha propuesto que el magnesio y el calcio son cationes que se encuentran involucrados en la reducción de la actividad del aluminio a nivel de la membrana plasmática o bien en la reducción de la competencia por los sitios de unión en el apoplasma radial (Kinraide, 1998). Específicamente en soya, Silva et al. (2001) demostraron que bajas concentraciones de magnesio (intervalos micromolares) fueron altamente efectivas en disminuir los efectos tóxicos del aluminio; no obstante, estos resultados no pudieron ser explicados por cambios en el potencial y en la actividad del Al3+ a nivel de la superficie de la membrana plasmática de la raíz como lo indica el modelo de Gouy-Chapman-Stern, lo que sugiere que el Mg2+ no actúa como un catión indiferente cuando es adicionado a bajas concentraciones, y supone la existencia de un mecanismo que va más allá de efectos estrictamente electrostáticos en la superficie de la raíz. Por otro lado, en presencia de complejos orgánicos de Al3+, existe menos Mg2+ remplazado de los sitios de unión en el apoplasto y en consecuencia la asimilación del Mg2+ es inhibida en menor grado (Kochian et al., 2004). Yang et al. (2007) han demostrado que el Mg aún en concentraciones bajas (10 μM), favorece el eflujo de citrato inducido por la presencia de aluminio en raíces de Vigna μMbellata ([Thunb.] Ohwi & Ohashi). Estos mecanismos incluyen la expresión diferencial de genes que codifican proteínas que regulan la entrada y el transporte de Al3+ en la célula vegetal (Kochian et al., 2004), lo cual debería ser explorado en próximas investigaciones con frijol, especie que ha sido clasificada como muy sensible a bajos valores de pH y con una amplia gama de diferencias genotípicas en lo que a sensibilidad a protones respecta (Rangel et al., 2005).
El análisis de los resultados aquí observados lleva a recomendar la realización de nuevos estudios, en los cuales se cambien las concentraciones de Ca y Mg en el medio de crecimiento para evaluar el efecto real que estos cationes tienen en la inactivación de los iones Al3+. Adicionalmente, se sugiere la identificación de mecanismos de tolerancia a Al3+ como exudación de ácidos orgánicos, acumulación de Al3+ en raíces, y las relaciones de acumulación de Al3+ y cationes (Al3+/ (K+ + Ca2+ + Mg2+)), entre otros, los cuales deben ser evaluados en la variedad Grijalva con concentraciones de Al3+ mayores a 100 μM en soluciones nutritivas con menor fuerza iónica, lo que además permitirá definir si los mecanismos de tolerancia a Al3+ son ocasionados por factores genéticos o por procesos celulares.
CONCLUSIONES
- Los factores variedad, concentración de aluminio (Al3+) y sus interacciones afectaron la concentración de cationes en hojas, tallos y raíces de frijol. El Al3+ modificó significativamente la concentración de potasio (K+) en hojas, y de manera altamente significativa la concentración de este catión en tallos y raíces. Sin embargo, el factor variedad sólo afectó la concentración de K+ en tallo. La interacción de ambos factores fue significativa únicamente para la concentración de K+ en tallos.
- La concentración de Al3+ afectó de manera altamente significativa la concentración de calcio (Ca2+) en los 3 órganos analizados; sin embargo, el factor variedad no tuvo efecto sobre la concentración de este elemento en los órganos analizados, aunque si lo fue la interacción entre estos factores. La concentración de magnesio (Mg2+) fue afectada de manera altamente significativa por el factor concentración de Al3+ en los 3 órganos analizados; en tanto que el factor variedad afectó la concentración de este elemento solo en tallos y raíces; mientras que la interacción entre los 2 factores de estudio afectó la concentración de este elemento en hojas y tallos, pero no en raíces. Se observó que la variedad Grijalva fue la menos afectada por la presencia de Al3+ en la solución nutritiva. En esta variedad, la concentración de Mg2+ en hoja fue incrementada en plantas tratadas con aluminio en comparación con el testigo.
AGRADECIMIENTO
Los autores agradecen al Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) por haber facilitado las variedades de frijol evaluadas y a la Línea Prioritaria de Investigación No. 5 Biotecnología Microbiana, Vegetal y Animal del Colegio de Postgraduados por las facilidades de equipo brindadas para la realización de este trabajo.
LITERATURA CITADA
Alcántar, G. C. y M. Sandoval V. 1999. Manual de análisis químico de tejido vegetal. Guía de muestreo, preparación, análisis e interpretación. Publicación Especial No. 10 de la Sociedad Mexicana de la Ciencia del Suelo A. C. Chapingo, México. [ Links ]
Da Silva, I. R., A. Ferrufino, C. Sanzonowicz, T. J. Smyth, D. W. Israel, and T. E. Carter-Junior. 2005. Interactions between magnesium, calcium, and aluminum on soybean root elongation. Rev. Bras. Cienc. Solo 29: 747-754. [ Links ]
Delhaize, E., P. R. Ryan, and P. J. Randall. 1993. Aluminum tolerance in wheat (Triticum aestivum L.). 2. Aluminum-stimulated excretion of malic acid from root apices. Plant Physiol. 103: 695-702. [ Links ]
Fox, T. C. and M. L Guerinot. 1998. Molecular biology of cation transport in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49: 669-696. [ Links ]
García, E. 1988. Modificaciones al sistema de clasificación climática de Köppen. 4a Edición. Instituto de Geografía. UNAM. México, D.F. [ Links ]
Grimme, H. 1983. Aluminium induced magnesium deficiency in oats. Z. Pflanzenernaeh. Bodenk. 146: 666-676. [ Links ]
Hoagland, D. R. and I. Arnon. 1950. The water culture method for growing plants without soil. Calif. Agr. Expt. Sta. Circ. No. 347. Berkeley, CA, USA. [ Links ]
Jones, D. L., L. V. Kochian, and S. Gilroy. 1998. Aluminum induces a decrease in cytosolic calcium concentration in BY-2 tobacco cell cultures. Plant Physiol. 116: 81-89. [ Links ]
Keltjens, W. G. and K. Tan. 1993. Interactions between aluminium, magnesium and calcium with different monocotyledonous and dicotyledonous plant species. Plant Soil 155: 458-488. [ Links ]
Kinraide, T. B. 1998. Three mechanisms for the calcium alleviation of mineral toxicities. Plant Physiol. 118: 513-520. [ Links ]
Kochian, L. V., O. A. Hoekenga, and M. A. Piñeros. 2004. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Ann. Rev. Plant Biol. 55: 459-93. [ Links ]
Kollmeier, M., P. Dietrich, C. S. Bauer, W. J. Horst, and R. Hedrich. 2001. Aluminum activates a citrate-permeable anion channel in the aluminum-sensitive zone of the maize root apex: a comparison between an aluminum-sensitive and an aluminum-resistant cultivar. Plant Physiol. 126: 397-410. [ Links ]
Lee, J. and W. Pritchard. 1984. Aluminium toxicity expression nutrient uptake, growth and root morphology of Trifolium repens L. cv. "Grasslands Huia". Plant Soil 82: 101-116. [ Links ]
Liu, K. and S. Luan. 2001. Internal aluminum block of plant inward potassium channels. Plant Cell 13: 1453-1465. [ Links ]
Liu, N. G., B. B. Mo, X. L. Yan, and H. Shen. 2007. Physiological mechanisms of soybean and rice in responses to aluminum stress. Ying Yong Sheng Tai Xue Bao 18: 853-858. [ Links ]
Marschner, H. 1995. Mineral nutrition of higher plants. Academic Press. London, UK. [ Links ]
Poschenrieder, Ch., N. Massot, P. Guevara y J. Barceló. 1992. Aluminio: II. Mecanismos de toxicidad en hombre, animales y plantas. Cir. Far. 50: 27-42. [ Links ]
Rangel, A. F., M. Mobin, I. M. Rao, and W. J. Horst. 2005. Proton toxicity interferes with the screening of common bean (Phaseolus vulgaris L.) genotypes for aluminium resistance in nutrient solution. J. Plant Nutr. Soil Sci. 168: 607-616. [ Links ]
Rengel, Z. and D. C. Elliot. 1992. Mechanism of aluminum inhibition of net 45Ca2+ uptake by Amaranthus protoplasts. Plant Physiol. 98: 632-638. [ Links ]
Rout, G. R., S. Samantaray, and P. Das. 2001. Aluminium toxicity in plants: a review. Agronomie 21: 3-21. [ Links ]
Samac, D. A. and M. Tesfaye. 2003. Plant improvement for tolerance to aluminum in acid soils. A review. Plant Cell Tissue Organ Cult. 75: 189-207. [ Links ]
SAS. 1985. SAS user's guide: Statistics. 8th ed. SAS Institute. Cary, NC, USA. [ Links ]
Schofield, R. M. S., J. Pallon, G. Fiskesjö, G. Karlson, and K. G. Malmqvist. 1998. Aluminium and calcium distribution patterns in aluminium-intoxicated roots of Allium cepa do not support the calcium displacement hypothesis and indicate signal mediated inhibition of root growth. Planta 205: 175-180. [ Links ]
Silva, I. R., T. J. Smyth, D. W. Israel, C. D. Raper, and T. W. Rufty. 2001. Magnesium is more efficient than calcium in alleviating aluminum rhizotoxicity in soybean and its ameliorative effect is not explained by the Gouy-Chapman-Stern model. Plant Cell Physiol. 42: 538-545. [ Links ]
Steiner, A. A. and H. van Winden. 1970. Recipe for ferric salts of ethylenediaminetetraacetic acid. Plant Physiol. 46: 862-863. [ Links ]
Sumner, M. E. and A. D. Noble. 2003. Soil acidification: The world story. pp. 1-28. In: Z. Rengel (ed.). Handbook of soil acidity. Marcel Dekker. New York, NY, USA. [ Links ]
Tanoi, K., J. Hojo, K. Suzuki, Y. Hayashi, H. Nishiyama, and T. M. Nakanishi. 2006. Analysis of potassium uptake by rice (Oriza sativa) roots treated with aluminum using a positron emitting nuclide, 38K. Soil Sci. Plant Nutr. 51: 715-717. [ Links ]
Tuma, J., M. Skalicky, L. Tumová, P. Bláhová, and M. Rosulková. 2004. Potassium, magnesium and calcium content in individual parts of Phaseolus vulgaris L. plant as related to potassium and magnesium nutrition. Plant Soil Environ. 50: 18-26. [ Links ]
Van Praag, H. J., F. Weissen, P. Dreze, and M. Cogneau, M. 1997. Effects of aluminium on calcium and magnesium uptake and translocation by root segments of whole seedlings of Norway spruce (Picea abies Karst.). Plant Soil 189: 267-273. [ Links ]
Watanabe, T. and K. Okada. 2005. Interactive effects of Al, Ca and other cations on root elongation of rice cultivars under low pH. Ann. Bot. 95: 379-385. [ Links ]
Yang, J. L., J. F. You, Y. Y. Li, P. Wu, and S. J. Zheng. 2007. Magnesium enhances aluminum-induced citrate secretion in rice bean roots (Vigna umbellata) by restoring plasma membrane H+-ATPase activity. Plant Cell Physiol. 48: 66-73. [ Links ]

