










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkTerra Latinoamericana
versão On-line ISSN 2395-8030versão impressa ISSN 0187-5779
Terra Latinoam vol.28 no.2 Chapingo Abr./Jun. 2010
División I
Salinity effect on Prosopis laevigata seedlings
Efecto de la salinidad sobre plántulas de Prosopis laevigata
Ramiro Ríos-Gómez1*, Carlos Enrique Salas-García1, Arcadio Monroy-Ata2 and Eloy Solano1
1 Facultad de Estudios Superiores Zaragoza, Unidad de Investigación en Sistemática Vegetal y Suelo. *Autor responsable: riosgr55@yahoo.com.mx.
2 Unidad de Investigación en Ecología Vegetal, UNAM. Apartado Postal 9-020, 09230, México, D.F.
Recibido: junio de 2008.
Aceptado: abril de 2010.
SUMMARY
The effect of salinity on emergence and development of Prosopis laevigata seedlings was studied in a greenhouse. Five saline mixtures and a control of distilled water were used throughout the experimental stage. The mixtures were chloride, sulfate-chloride, chloride-sulfate, sulfate, and sulfate-sodium, which generated electric conductivities (EC) of 1.0, 2.0, 3.5, 5.0, 7.5, and 9.0 dS m-1. No effect on emergence, relative growth rate, cover, number and length of roots or root and shoot weight was observed in relation to the control for chloride or sulfate-chloride treatments with EC above 5 dS m-1. Treatments with chloride-sulfate and sulfate mixtures were detrimental to emergence and initial development at EC above 5 dS m-1. Our results suggest that in an EC below 5 dS m-1, P. laevigata could be used for rehabilitation of soils affected by salinity.
Keywords: electrical conductivity, emergence, saline mixture, saline soils.
RESUMEN
En el estudio se evaluó el efecto de la salinidad sobre la emergencia y el desarrollo de plántulas de Prosopis laevigata bajo condiciones de invernadero. Se emplearon 5 mezclas salinas, clorhídrica, sulfático-clorhídrica, clorhídrico-sulfática, sulfática y sulfático-sódica; las cuales generaron conductividades eléctricas (CE) de 1.0, 2.0, 3.5, 5.0, 7.5, 9.0 dS m-1 y un testigo con agua destilada. Durante su emergencia y desarrollo, las plántulas de P. laevigata tratadas con las mezclas clorhídrica y sulfático-clorhídrica, no redujeron emergencia, tasa relativa de crecimiento, cobertura, número y longitud de raíces, ni el peso de raíz y vástago en relación con el testigo cuando la CE fue superior a 5 dS m-1. Los tratamientos con las mezclas clorhídrico-sulfática y sulfática, disminuyeron la emergencia y el desarrollo inicial de las plántulas a una CE superior a 5 dS m-1. Los resultados sugieren que por debajo de una CE de 5 dS m-1 P. laevigata podría ser utilizada en la rehabilitación de suelos afectados por salinidad.
Palabras clave: conductividad eléctrica, emergencia, mezclas salinas, suelos salinos.
INTRODUCTION
Several authors have studied the response of Prosopis to saline stress, though they have been mainly focused on effects produced by sodium chloride (Villagra, 1997; Cony and Trione, 1998; Cazebonne et al., 1999); in the nature, however, salinity is caused by the interaction of more than one salt, for example, salt mixtures composed by sodium chloride/magnesium sulfate (as in coastal zones) or sodium bicarbonate/sodium chloride plus magnesium sulfate (as in sodic soils). The germination of Prosopis is negatively affected by NaCl (Rhodes and Felker, 1988; Catalán et al., 1994; Perez and Vieira, 1994) and it has been shown that the magnitude of the damage due to NaCl varies among species (Felker et al., 1981; Pandey and Thakarar, 1997; Velarde et al., 2003). Reinoso et al. (2000) found that germination is the most sensitive phase in P. farcta, P. strombulifera and P. flexulosa. Similarly, Serrato et al. (1992) and Reinoso et al. (2004) reported that NaCl causes structural alteration of the roots, hypocotyl, epicotyl, and leaves of P. cineraria, P. tamarugo and P. strombulifera.
Reinoso et al. (2000) showed that salinity damage may vary among different salts; they observed that germination and growth reduction of P. juliflora and P. strombulifera seedlings was less affected by Na2SO4 than by NaCl. Nonetheless, Jarrell and Virginia (1990) found that P. glandulosa var. torreyana seedlings can extract water at saline soils with EC higher than 20 dS m-1. The ability to assimilate water with elevated salt content is aided by the capacity of Prosopis species to exclude salt from its aboveground biomass, accumulating it in the roots instead (Jarrell and Virginia, 1984; Zhao and Harris, 1992). Furthermore, P. chilensis, P. flexuosa and P. alba seedlings exposed to NaCl developed more root than shoot tissue in order to evade an elevated saline content. This behavior has also been identified in P. laevigata (Meloni et al., 2004).
Given that some Prosopis species are tolerant to salinity and that Mexico has an elevated quantity of affected soils, these species could be utilized for rehabilitation. Of the nine species of Prosopis found in Mexico, Prosopis laevigata is particularly noteworthy. It is typically found in the central regions of Mexico (Rzedowski, 1988; Palacios et al., 2000), and is widely used for a variety of purposes including firewood, forage, human consumption, and construction (Galindo and García-Moya, 1986; Villanueva, 1993). Additionally, Prosopis laevigata contributes to the maintenance of ecological balance in semi-arid ecosystems by generating islands of resources with high nutrient concentrations in the soil and allowing the development of other organisms under its canopy (Olalde et al., 2000; Montaño et al., 2006, Perroni-Ventura et al., 2010). In this context, the goal of the current study was to answer the following: How does salinity influence the emergence and development of Prosopis laevigata seedlings? For this purpose, we conducted an experimental approach to assess the effect of five saline mixtures on the emergence and initial growth of Prosopis laevigata seedlings under greenhouse conditions.
MATERIALS AND METHODS
Collection of Biological Material
Herbarium samples and fruits from 15 to 20 trees of Prosopis laevigata were collected from xeric scrub habitat between 1800 and 1900 of altitude. San Nicolás and Tasquillo in the Hidalgo State, Mexico. Herborized samples were determined taxonomically and deposited in the "Facultad de Estudios Superiores Zaragoza (FEZA)" Universidad Nacional Autónoma de México herbarium with the number 5601. In this area P. laevigata grows on soils with electrical conductivities (CE) varying from 2 to 41 dS m-1, and 4 or more dS m-1 are considered as saline according to Ríos-Gómez (unpublished data). P. laevigata is used as forage, human food, medical item and firewood in the Mezquital Valley, Hidalgo State (Meraz et al., 1998). P. laevigata is usually a tree with a height from 5 to 10 m and up to 50 cm in diameter, often flowering when smaller; herbage sparingly pubescent; spines from 0.3 to 4 cm long, stout, divergent pinnae one or two pairs; leaflets 1-2 mm wide, linear; spikes 39 cm long; corolla between 2.5 and 4 mm long; fruit compressed or turgid, more or less constricted among seeds, stipitate 5.10 mm, nearly one cm wide, 15-20 cm long, with up to 20-25 seeds.
Experimental Design
The experimental study was carried out in a greenhouse at the "Facultad de Estudios Superiores Zaragoza (FEZA)" Universidad Nacional Autónoma de México, during July-August 2001. Experimental treatments were performed under greenhouse conditions. Black plastic 1 L pots filled with 100 g of perlite were used as experimental units. In each pot 13 seeds were uniformly distributed on the perlite and covered with a gravel volcanic layer, and 330 mL of the corresponding saline solution was added. The ten pots set were weighted every 48 hours in order to know the water loss; the average weight was subtracted from the weight of two days before and the mass difference pointed the water amount that should be added. By this way, the water content and salinity levels were maintained by the constant addition every 48 h of the quantity of water necessary to replace losses by evapotranspiration. The experiment was continued for 19 days post-emergence. The experimental design was a randomized complete block design of two factors: five mixtures and seven EC levels, with five replicates for a total of 175 experimental units.
Saline Mixture
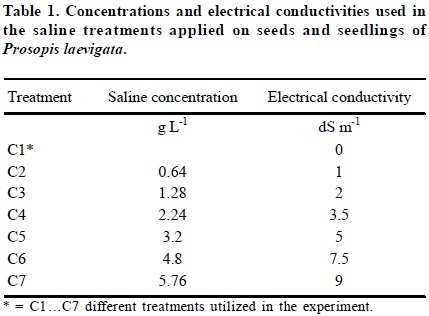
Saline mixtures were prepared according to the method of Arinushkina (1970), in which it is established that a salt concentration of 0.64 g L-1 generates an EC of 1.0 dS m-1. Elaboration of the treatments considered were determined previously based on EC values from the saline soils in which Prosopis laevigata is found in the Mezquital Valley, Hidalgo State, Mexico (Table 1). The quantity of salt in each mixture was calculated using these EC values and the previously described method. Table 2 shows the saline mixtures used and the corresponding ionic relations.

Response Variables
Seedlings were considered as emerged when their cotyledons rose above the surface of the substrate. Emerged seedlings were counted daily until the number remained unchanged for three days. Five seedlings from each experimental unit were selected to evaluate relative growth rate (RGR) registering the height of seedlings daily, from root crown to the shoot apex. The plant cover was estimated by mean of the average of canopy diameter line and its perpendicular diameter, thus the medium radius was obtained and used to calculate seedling circle area cover. The number of secondary roots, length of the primary root, and fresh and dry weight of shoot and root were estimated by direct counts and measurements or weight differences, respectively.
To determine weight, the root was separated from the shoot at the height of the root crown; each part was weighed, oven-dried at 70 °C for 48 h, and then weighed again. Plant mortality was quantified daily.
Emergence was determinated by direct counts and it was expressed as percentage and the Maguire's index for the emergence rate was also calculated (González-Zertuche and Orozco-Segovia, 1996). Length and width of cotyledons and true leaves were measured, and the average of the two measurements was used to calculate cover. The RGR was calculated according to Hunt (1990). Secondary root numbers were counted and the length of the primary root was measured with a ruler graduated in millimeters. The root/shoot ratios (R/S) were calculated using fresh and dry weight of the root and shoot and with the formula proposed by Böhm (1979).
Statistical Analysis
To understand the effect of saline mixtures on the response variables and because the data set did not behave as a normal distribution, a Kruskal-Wallis test was employed, using a 95% confidence interval (Montgomery, 1991). The study of root and shoot dry weight was carried out by an analysis of covariance with 95% confidence intervals, where root and shoot weight were the covariables. Statistical analyses were performed in the software program StatGraphics Plus version 4.0.
RESULTS AND DISCUSSION
Emergence
In the chloride and sulfate-chloride mixtures neither the percentage of emergence nor the rate of emergence were reduced as concentration increased, while the sulfate treatment reduced emergence up to 57% (Figure 1).
The literature indicates that NaCl reduces the germination in Prosopis (Villagra and Galera, 1992; Pandey and Thakarar, 1997; Villagra, 1997; Cazebonne et al., 1999). However in this study, chloride and sulfate-chloride saline mixtures (characterized by an ionic relation higher than that of sulfates) had no effect on the germination. Emergence was significantly affected by saline mixtures in which sulfate ions had higher concentration than chlorine. Previously, Reinoso et al. (2000) and Pérez and Tambelini (1995) found that the germination in P. strombulifera and P. juliflora was more sensitive to Na2SO4 than to NaCl, which suggests that the genus Prosopis is sensitive to sulfate ions, as was shown in this study.
Relative Growth Rate (RGR)
The RGR in the chloride and sulfate-chloride mixtures was not affected by treatments C1 to C4, and in fact occurred an increase in C7 treatment, though it was not significant at the 95% level of confidence (Figure 2). In contrast, in the sulfate and chloride-sulfate mixtures, as salinity increased, RGR was reduced by more than 50% with respect to the control (Figura 2).
Cover
Chloride and sulfate-chloride mixtures did not reduce seedling cover (Figure 3), but it was reduced up to 54 and 70% (P • 0.05) with respect to the control in the chloride-sulfate and sulfate mixtures, respectively. Cazebonne etal. (1999) obtained results similar to those found in this study. In these studies salinity reduced both growth and cover of seedlings. In our study, the reduction observed in both variables was statistically significant (P < 0.05) only when sulfate ions dominated and not for chloride or sulfate-chloride mixtures in which chlorine predominated. Similarly, Reinoso et al. (2000) and Pérez and Tambelini (1995) observed that seedling growth of P. strombulifera and P. juliflora was more affected by Na2S04 than by NaCl; our results suggest that P. laevigata is sensitive to the sulfate ion, due to that presumably the sulfates promote the entrance of sodium and restrict the assimilation of calcium so that sulfate and sodium act synergistically to reduce plant growth (JoshiandNaik, 1980).
Number and Length of Roots
Chloride and sulfate-chloride mixtures did not reduce the number or length of seedling roots (Figures 4 and 5).
In contrast, the chloride-sulfate and sulfate mixtures decreased root production up to 78%, while radical growth was reduced up to 93% relative to the control (P • 0.05). Meraz etal. (2000) found that salinity caused by NaCl reduced the number and length of roots in seedling of Prosopis laevigata, but in our study these variables were only significantly affected by the chloride-sulfate and sulfate mixtures. The results showed that the chloride and sulfate-chloride mixtures did not reduce the number or length of roots, because the elevated proportion of the chlorine ion that could be generating a water content superior to 80% in vegetative tissues, which regulated osmotic pressure, when exposed to excessive salinity as occurs in halophytes plants (Levitt, 1972; Bernstein, 1975; Kramer, 1989; White and Broadley, 2001).
Fresh and Dry Weight
In the chloride and sulfate-chloride mixtures fresh and dry weight of root and shoot were not significantly reduced, and seedlings exhibited water content higher than 80%. In contrast, the chloride-sulfate and sulfate mixtures significantly decreased the above-mentioned variables (P < 0.05), and seedling water content was less than 80% (Table 3). All mixtures, however, had R/S ratios less than one for dry root weight, which suggests a greater resources allocation to the shoot of seedlings.
On the other hand, the chlorine-sulfate mixture reduced fresh weight due to the scarce growth of shoot and root influenced by the sulfate ion; an even greater quantity of sulfate ions (to which seedlings of Prosopis laevigata are sensitive) was found in the sulfate mixture (Table 3). Strogonov (1964) found that sulfate ions may promote a weaker accumulation of water compared to chlorine, since the respiration increases, the water potential of the leaves is reduced, osmotic pressure is elevated and the viscosity of the protoplasm is depleted.
In previous studies with Prosopis flexuosa, P. chilensis and P. laevigata have documented R/S ratios greater than one as a response to low hydric potential (Catalán et al., 1994; Pandey and Thakarar, 1997). In the current investigation the R/S ratios were below one, influenced by the saline mixtures which provoked oxidation and necrosis in the subapical meristem as concentrations increased. Below an EC of 5 dS m-1, the germination, emergence, and growth of P. laevigata seedlings were not affected, regardless of the salinity type present in the medium around the roots. Under these conditions, this species could be used for rehabilitation of soils affected by salinity.
CONCLUSIONS
- The seeds and seedlings of Prosopis laevigata showed low sensitivity to chloride-sulfate and chloride mixtures. There was no reduction in seedling emergence, relative growth rate (RGR), cover, number, or longitude of the roots or on the weight of the roots and shoot, when the electrical conductivity (EC) was lower than 8 dS m-1. However, these variables show a reduction when the EC was higher than 5.0 dS m-1 based on mixtures of chloride-sulfate, sodium-sulfate and sulfate. The greatest mortality of seedlings occurs in mixtures with chloride-sulfate and sulfate salinities.
- The Prosopis laevigata seedlings could grow on soils with salinity problems, including those with an EC of up to 5.0 dS m-1, because of seedling development is reduced when the salinity increases. Across the different salinities here studied, the root was the most affected organ; its longitude and weight was reduced when the concentration and time of exposure of sulfate ions increased. The aboveground had the greatest biomass.
ACKNOWLEDGEMENTS
We thank A. Cervantes Sandoval for statistical advice and Ma. del C. López Barriga for linguistic improvement on the manuscript. This study was supported by PAPIIT IN216610, UNAM. We also thank the helpful comments of anonymous reviewers, who helped us to improve the manuscript for publication.
REFERENCES
Arinushkina, E. V. 1970. Manual de análisis químico de suelos. Parte II. Ed Universidad de Moscú. URSS. [ Links ]
Bernstein, L. 1975. Effects of salinity and sodicity on plant growth. Annu. Rev. Phytopathol. 13: 295-312. [ Links ]
Böhm, W. 1979. Methods of studying root systems. Springer-Verlag. Berling, Germany. [ Links ]
Catalán, L., M. Balzarini, E. Taleisnik, R. Sereno, and U. Karlin. 1994. Effects of salinity on germination and seedling growth of Prosopis flexulosa (D.C.). For. Ecol. Manage. 63: 347-357. [ Links ]
Cazebonne, C., A. I. Vega, D. A. Varela y L. A. Cardemil. 1999. Efectos de la salinidad en la germinación y crecimiento de Prosopis chilensis. Rev. Chil. Hist. Nat. 72: 83-91. [ Links ]
Cony, M. A. and S. O. Trione. 1998. Inter-and intraespecific variability in Prosopis flexuosa and Prosopis chilensis: seed germination under salt and moisture stress. J. Arid Environ. 40: 307-317. [ Links ]
Felker, P., P. R. Clark, A. E. Laag, and P. F. Pratt. 1981. Salinity tolerance of the tree legumes: mesquite (Prosopis glandulosa var. torreyana, P. velutina y P articulata) algarrobo (P. chilensis), Kiawe (P. pallida) and tamarugo (P. tamarugo) grown in sand culture on nitrogen-free media. Plant Soil 61: 311-317. [ Links ]
Galindo, A. S. y E. García-Moya. 1986. Usos del mezquite (Prosopis L.) en el Altiplano Potosino. Agrociencia 63: 7-15. [ Links ]
González-Zertuche, L. y A. Orozco-Segovia. 1996. Métodos de análisis de datos en la germinación de semillas, un ejemplo: Manfreda brachystachya. Bol. Soc. Bot. Méx. 58: 15-30. [ Links ]
Hunt, R. 1990. Basic growth analysis. Academic Press. London, UK. [ Links ]
Jarrell, W. M. and R. A. Virginia. 1984. Salt tolerance of mesquite. Calif. Agric. 38: 28. [ Links ]
Jarrell, W. M. and R. A. Virginia. 1990. Response of mesquite to nitrate and salinity in a simulated phreatic environment: water use, dry matter and mineral nutrient accumulation. Plant Soil 125: 185-196. [ Links ]
Joshi, G. V. and G. R. Naik. 1980. Response of sugarcane to different types of salt stress. Plant Soil 56: 255-263. [ Links ]
Kramer, P. J. 1989. Relaciones hídricas de suelos y plantas. Harla. México, D.F. [ Links ]
Levitt, J. 1972. Response of plants to environmental stresses. Academic Press. New York, NY, USA. [ Links ]
Meloni, D. A., M. R. Gulotta, C. A. Martínez, and M. A. Oliva. 2004. The effect of salt stress on growth, nitrate reduction proline and glycinebetaine accumulation in Prosopis alba. Braz. J. Plant Physiol. 16: 39-46. [ Links ]
Meraz V., S., J. Orozco V., J. A. Lechuga C., F. Cruz S. y J. Vernon C. . 1998. El mezquite, árbol de gran utilidad. Ciencias 51: 20-21. [ Links ]
Montaño-Arias, N. M., R. García-Sánchez, G. Ochoa-de la Rosa y A. Monroy-Ata. 2006. Relación entre la vegatación arbustiva, el mezquite y el suelo de un ecosistema semiárido en México. Terra Latinoamericana 24: 193-205. [ Links ]
Montgomery, D. C. 1991. Diseño y análisis de experimentos. Iberoamericana. México, D.F. [ Links ]
Olalde, V., J. T. Frías, A. L. Aguilar, N. Pescador, y L. I. Aguilera. 2000. Caracterización microbiológica de suelos de islas de fertilidad de mezquite [Prosopis laevigata (Humb. & Bonpl. ex. Wild) M. C. Johnst.] en ambientes semiáridos. pp. 95-107. In: V. Frías-Hernández, E. Olalde, and J.Vernon (eds.). El mezquite árbol de usos múltiples: estado actual del conocimiento de México. Universidad de Guanajuato. Guanajuato, México. [ Links ]
Palacios, R. A., P. S. Hoc, A. D. Burghardt y A. E. Vilela. 2000. Prosopis L. Biodiversidad y clasificación, estrategias adaptativas reproductivas e importancia económica. pp. 13-34. In: V. Frías-Hernández, E. Olalde, and J.Vernon (eds.). El mezquite árbol de usos múltiples: estado actual del conocimiento de México. Universidad de Guanajuato. Guanajuato, México. [ Links ]
Pandey, A., N. and N. K. Thakarar. 1997. Effect of chloride salinity on survival and growth of Prosopis chilensis seedlings. Trop. Ecol. 38: 145-148. [ Links ]
Pérez, S. C. J. G. de A. y J. A. P. V. de Moraes. 1994. Estresse salino no processo germinativo da algarobeira e atenuacao de seus efeitos pelo uso de reguladores de crescimiento. Pesquisa Agropecuária Brasileira 29: 389-396. [ Links ]
Pérez, S. C. J. G. de A. y M. Tambelini. 1995. Efeito dos estresses salino e hidrico e do envelhecimento precoce na germinacao de algarobeira. Pesquisa Agropecuária Brasileira 30: 1289-1295. [ Links ]
Perroni-Ventura, Y., C. Montaña, and F. García-Oliva. 2010. Carbon-nitrogen interactions in fertility island soil from a tropical semi-arid ecosystem. Funct. Ecol. 24: 233-242. [ Links ]
Reinoso, H., L. Sosa, C. Combo, F. Ochoa, and V. Luna. 2000. Morphological and physiological responses of Prosopis strombulifera (Lam.) bent to increasing salt conditions. pp. 201-202. In: D. Bush, D. Cosgrove, R. Hangarter, R. Jorgensen, D. Delmer, P. Springer, W. Lucas and D. Schnell (eds.). Plant biology. 2000. American Society of Plant Physiology. San Diego, CA, USA. [ Links ]
Reinoso, H., L. Sosa, L. Ramírez, and V. Luna. 2004. Salt-induced changes in the vegetative anatomy of Prosopis strombulifera (Leguminosae). Can. J. Bot. 82: 618-628. [ Links ]
Rhodes, D. and P. Felker. 1988. Mass screening of Prosopis (mesquite) seedlings for growth at seawater salinity concentrations. For. Ecol. Manage. 24: 169-176. [ Links ]
Rzedowsky, J. 1988. Análisis de la distribución del complejo Prosopis (Leguminoseae, Mimosoideae) en Norteamérica. Acta Bot. Mex. 3: 7-19. [ Links ]
Serrato V., G., L. Melone, O. Orsi, and F. Riveros. 1992. Anatomical changes in Prosopis cineraria (L.) Druce seedlings growing at different levels of NaCl salinity. Ann. Bot. 70: 399-404. [ Links ]
Strogonov, B. P. 1964. Physiological basis of salt tolerance of plants (as affected by various types of salinity). Akad. Nauk. SSSR. Isr. Progr. Sci. Transl. Jerusalem, Israel. [ Links ]
Velarde, M., P. Felker, and C. Degano. 2003. Evaluation of Argentine and Peruvian Prosopis germoplasm for growth at seawater salinities. J. Arid Environ. 55: 515-531. [ Links ]
Villagra, P. E. 1997. Germination of Prosopis argentina and P. alpataco seeds under saline conditions. J. Arid Environ. 37: 261-267. [ Links ]
Villanueva, J. D. 1993. Distribución actual y características ecológicas del mezquite (Prosopis laevigata H. & B. Johnst.), en el estado de San Luis Potosí. INIFAP. México, D.F. [ Links ]
White, P. J. and M. R. Broadley. 2001. Chloride in soils and its uptake and movement within the plant: a review. Ann. Bot. 88: 967-988. [ Links ]
Zhao, K. and P. J. C. Harris. 1992. The effects of iso-osmotic salt and water stresses on the growth of halophytes and non-halophytes. J. Plant Physiol. 139: 761-763. [ Links ]

