










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkTerra Latinoamericana
versión On-line ISSN 2395-8030versión impresa ISSN 0187-5779
Terra Latinoam vol.27 no.2 Chapingo abr./jun. 2009
División II
Modelo expo–lineal del crecimiento y equivalencia de la productividad de un tomate cultivado en invernadero
Expolinear growth model and productivity equivalence of a greenhouse grown tomato
Nahielli Solorio Elizalde1, Fernando Paz Pellat1*, Magali Odi Lara1 y Martín A. Bolaños González1
1 Colegio de Postgraduados. Campus Montecillo. 56230 Texcoco, Estado de México. *Autor responsable (pellat@colpos.mx).
Recibido: noviembre de 2007.
Aceptado: agosto de 2008.
RESUMEN
El modelo expo–lineal del crecimiento de las plantas en la etapa vegetativa ha sido propuesto como un esquema semi–empírico de modelación, con resultados aceptables en su aplicación a datos experimentales. En el presente trabajo, este modelo se revisa en su forma general y se aplica a un experimento de tomate cultivado en invernadero sujeto a diferentes combinaciones de niveles de estrés hídrico y nitrógeno. Los resultados muestran que los ajustes del modelo son buenos para la materia aérea seca y el área foliar, pues validan su uso en condiciones que difieren de las óptimas determinadas. Asimismo, se discute y revisa el concepto de equivalencia ambiental en la productividad usando los datos experimentales. Los resultados obtenidos son aplicables, lo que define un camino para una modelación compacta del crecimiento de los cultivos.
Palabras clave: equivalencia ambiental en la productividad, tomate en invernadero.
ABSTRACT
The vegetation expolinear grown model in the vegetative stage has been proposed as a semi–empirical scheme for modeling, where acceptable results are obtained when applied to experimental data. In this paper, this model is reviewed in its general form and applied to a greenhouse tomato under a combination of different levels of water stress and nitrogen. The results show that the adjustments of the model are good for aerial dry matter and foliar area, validating in this form its use for conditions away from the optimum. Also, the environmental equivalence in productivity concept is discussed and reviewed using experimental data, where good results were obtained in its applicability, defining a way of modeling the growth of very compact vegetation.
Keywords: environmental equivalence in productivity, greenhouse tomato.
INTRODUCCIÓN
La modelación del crecimiento de los cultivos es una tarea compleja y difícil de realizar por la gran cantidad de información requerida. Debido al impacto económico del crecimiento de cultivos en invernadero, se debe contar con herramientas de modelación sólidas y relativamente simples. El tomate (Lycopersicon esculentum Mill.) es uno de los cultivos de invernadero que posee gran productividad y un manejo eficiente.
La productividad (principalmente materia y área foliar) de la vegetación depende del medio ambiente (temperatura y radiación solar, entre otros) en que se desarrolla, además de factores de estrés (agua, nutrientes, competencia, plagas y enfermedades, etc.). La modelación de los factores incidentes en la productividad puede hacerse usando modelos mecanicistas semi–empíricos. Para el caso del tomate en invernadero existen varios: HORTSIM (Gijzen et al., 1998); TOMGRO (Jones et al., 1991); SUKAM (Heuvelink, 1995). La alternativa a la modelación mecanicista es usar funciones matemáticas con bases empíricas (Hunt, 1982). En el caso de los modelos mecanicistas, el número de variables es muy alto para predecir una o dos variables de salida de interés, además de la necesidad de conocer su variabilidad temporal. En la modelación matemática empírica, las funciones matemáticas no tienen un significado biofísico, por lo que resultan muy restringidas en las aplicaciones.
Como una alternativa a lo antes mencionado, en este trabajo se analizan las bases del modelo expo–lineal del crecimiento (Ferrandino, 1989; Goudriaan y Monteith, 1990) y se usa para modelar la etapa vegetativa del crecimiento en invernadero. La fase lineal del modelo es usada para analizar el concepto de equivalencia en la productividad.
Modelo expo–lineal del crecimiento
En el espacio del tiempo la modelación del crecimiento de la vegetación, índice de área foliar (IAF) y materia (M) en peso por unidad de área; representadas en forma general por la variable V en la etapa vegetativa, puede aproximarse como compuesta de dos fases: una exponencial y una lineal.
Crecimiento exponencial:

Crecimiento lineal:

donde R es la tasa relativa de crecimiento, C es tasa máxima crecimiento y tb es el tiempo obtenido al proyectar la fase lineal para intersectar el eje del tiempo.
Las dos fases de la etapa vegetativa pueden ser analizadas en forma independiente haciéndolas compatibles en los puntos de transición (Lieth et al., 1996) o podemos generar un modelo con una transición gradual entre ambos tipos de crecimiento. En el último caso, en el momento de la transición del crecimiento exponencial al lineal, las tasas de crecimiento deben ser iguales:

El modelo expo–lineal es simplemente la solución de los dos tipos de crecimiento representados por las ecuaciones diferenciales anteriores sujeta a la condición dada por la relación de igualdad de las tasas de crecimiento:

Con las siguientes características:

donde tT es el tiempo donde la condición (3) se cumple.
La Figura 1a muestra el modelo expo–lineal del crecimiento de la vegetación, mostrando las características definidas previamente. La Figura 1b muestra una transformación invariante (conservación de la métrica asociada) del modelo.
En la Figura 2 se muestra el crecimiento del índice de área foliar (IAF) (Huete et al., 1985) durante la fase reproductiva, donde las propiedades geométricas u ópticas varían con relación a la fase expo–lineal.

La transición de la fase exponencial a la reproductiva (representada por una función exponencial) puede ser modelada de forma similar en el caso de la transición exponencial a la lineal (Goudriaan, 1994); aunque el modelo es un sigmoide simétrico que puede generalizarse a uno asimétrico (Yin et al., 2003), pero con una pérdida de interpretación biofísica. Los modelos expo–lineales y sus generalizaciones discutidas utilizan la hipótesis de medios ambientes relativamente similares (principalmente la radiación fotosintética activa), por lo que es posible generalizarlos a medios ambientes variables (Monteith, 2000). No obstante, Yuan y Bland (2004) mostraron que los beneficios del modelo ambientalmente variable no mejora apreciablemente los resultados de ambiente constante, por lo que la estructura matemática del modelo expo–lineal (Ecuación 4) puede ser usada en términos generales.
Bases del modelo expo–lineal del crecimiento
De acuerdo a Monteith (1972) podemos definir una relación entre la materia y la radiación interceptada:

donde B es la materia seca, fIRFA, es la fracción de la radiación fotosintéticamente activa (RFA) interceptada y εM es el factor de eficiencia de conversión de la RFA interceptada por las plantas en materia seca. En realidad, la fracción que importa es la absorbida por la vegetación; sin embargo, el uso de la interceptada por la absorbida representa solo errores pequeños en la región de la RFA (Monteith, 1981).
En forma similar que para el caso de B, podemos plantear una ecuación para el IAF:

La fIFRA puede estimarse para un medio radiativamente turbio (Ross, 1981) como:

en donde K es un coeficiente de extinción.
Así, la Ecuación 8 puede ser reemplazada en función de IAF y diferenciada en relación al tiempo:

Igualando las Ecuaciones 7 y 8 y resolviendo para IAF tenemos:

Con las siguientes características:


donde RL es una tasa relativa de crecimiento, que para un medio ambiente constante implica:

De la estimación del IAF, es necesario asociar estos valores con la materia (M) de los cultivos. En la literatura relacionada con el análisis del crecimiento de las plantas, se utiliza el concepto de la razón de área foliar o RAF (LAR, leaf area ratio; por sus siglas en inglés):

Otro índice utilizado es el del área foliar específica o AFE (SLA, specific leaf area; por sus siglas en inglés):

El índice AFE puede usarse en forma instantánea, por lo que es más útil para su uso en la solución de ecuaciones diferenciales. Goudriaan y Monteith (1990) utilizaron la hipótesis de que la materia producida por el cultivo es proporcional al IAF:

en donde pf es la proporción del crecimiento de materia seca que se distribuye a la formación de nuevas hojas.
Cuando pf y AFE son constantes, entonces RAF = (pf)(AFE). La relación definida parte del concepto de que el incremento en materia es función de una tasa C dependiente del tiempo:

en donde CM es el valor máximo de C que puede ser logrado si toda la radiación incidente en el cultivo fuera interceptada (fIRFA = 1).
Usando una estrategia similar al caso del desarrollo de la ecuación expo–lineal para el IAF, se obtiene:

en donde RM es la tasa relativa de crecimiento en la fase exponencial, la cual se incrementa hasta alcanzar su valor máximo CM, al inicio de la fase lineal y está definida por:

La ecuación expo–lineal para el IAF y la materia, están relacionadas por:

El modelo expo–lineal de Goudriaan y Monteith (1990) fue desarrollado para la materia seca total de los cultivos; aunque es válido también para la materia seca (y verde) aérea.
Equivalencia de la productividad
Sin pérdida de generalidad podemos analizar sólo la fase lineal del modelo expo–lineal discutido anteriormente o una transformación aproximadamente lineal del modelo, tal como la mostrada en la Figura 1b. Para definir el contexto de las propiedades de los patrones del crecimiento de la vegetación, fase lineal, podemos analizar un conjunto de líneas rectas que se intersectan en un punto cualquiera (Figura 3a). Bajo esta consideración, las intersecciones ak y las pendientes bk de las líneas rectas (V = ak + bkt) tienen una relación lineal (Figura 3b), definiendo un espacio metaparamétrico dependiente de las constantes A y B (bk = A + Bak).
En el caso particular del modelo expo–lineal podemos suponer que la tasa de crecimiento relativa R es más o menos constante (los niveles del estrés, hasta un umbral crítico, tienen poco efecto en el espacio del tiempo), por lo que se espera que las fases lineales tengan un punto aproximado de intersección, tal como se muestra en la Figura 3a.
Considerando la variable de productividad Vi,j, donde i se refiere al nivel de estrés para un medio ambiente j (j = 1, por ejemplo, en nuestro caso), la Figura 4a muestra la situación donde el nivel de estrés i se manifiesta en un cambio de la pendiente bi,j en el espacio del tiempo. Haciendo un cambio de origen en el punto donde las fases lineales aproximadamente se intersectan (Figura 4a), podemos suponer que un cambio de pendiente implica un cambio en la productividad de la vegetación. En la Figura 4b se ha conservado el origen inicial de la Figura 4a, pero haciendo que las ai,j sean positivas (multiplicadas por –1) y la relación lineal entre ai,j y bi,j sea cambiada restando a los valores de bi,j la intersección con ese eje. Así, con la estimación de la pendiente de las fases lineales de la etapa vegetativa del crecimiento podemos estimar la productividad de la vegetación, ya que para una fecha determinada la pendiente es equivalente a una productividad V dada.
En la situación donde el medio ambiente y los niveles de estrés o manejo varían simultáneamente, la Figura 5a muestra la variación de la pendiente de las líneas rectas de la fase lineal (o la fase expo–lineal transformada linealmente) del crecimiento de la vegetación. En la Figura 5b (línea punteada más gruesa) se muestra el concepto de equivalencias ambientales en la productividad, donde para un valor de ésta, las intersecciones (ambientes y niveles de estrés) en la fase lineal del crecimiento de la vegetación pueden ser múltiples y equivalentes, dependiendo de i y j.
En la Figura 5a se muestra una línea recta radial, para cada diferente estrés, asociada a un ambiente potencial donde la vegetación puede alcanzar su potencial genético productivo, Figura 5b.
MATERIALES Y MÉTODOS
Experimento con tomate en invernadero
Para validar el comportamiento antes discutido, se realizó un experimento con tomate variedad Río Grande a través de la siembra de 500 semillas en un almácigo de charolas de plástico llenas con turba. La germinación ocurrió a los 3 días de la siembra y permaneció en el almácigo durante 42 días, tiempo durante el cual se le proporcionaron 14 riegos.
A continuación, se realizó el transplante en macetas de plástico con forma de cono truncado, conteniendo un sustrato de tezontle graduado a 2 mm de diámetro, con un orificio para drenar excedencias de agua y un filtro que impidió el deslave.
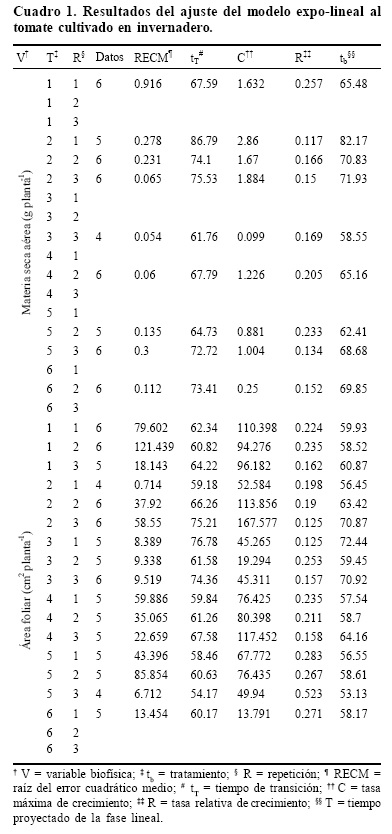
Los tratamientos al cultivo consistieron en variaciones de nitrógeno y humedad, en niveles óptimos y restrictivos. De estos factores, se propusieron tres niveles de nitrógeno (100, 50 y 10% respecto a la dosis óptima requerida por la planta) combinados con dos niveles de humedad (100 y 27% del consumo de la humedad aprovechable a capacidad de contenedor, determinada por mediciones directas), de donde se obtuvieron seis tratamientos. Para dosificar el N se procedió conforme al equivalente a la solución de Steiner (Steiner, 1984). De este modo, en el Cuadro 1, se muestran los tratamientos definidos según las variaciones de humedad y N. Se realizaron tres repeticiones de cada tratamiento, obteniéndose un total de 18 parcelas experimentales, las cuales fueron distribuidas al azar dentro del invernadero.
Con respecto al control de plagas del cultivo, fue necesario realizar 10 aplicaciones preventivas principalmente contra mosquita blanca usando PlenumMR (ingredientes activos: picloram 8% y fluoxipir 8%) diluído en agua (3 g L–1) y Dap–PlusMR (ingredientes activos: ácido fosfórico 32% y nonil fenol etoxilado 38%) (1 mL L–1), aplicado con mochila aspersora, a razón de 0.5 y 1 L por parcela experimental, dependiendo del crecimiento de las plantas.
RESULTADOS Y DISCUSIÓN
Ajuste del modelo expo–lineal del crecimiento del tomate en condiciones de invernadero
Dado que las mediciones del IAF o B son destructivas, la hipótesis de representatividad de estos datos para un tratamiento y repetición particular fueron revisadas usando diferentes espacios de representación y variables medidas. Sólo en aquellos casos donde existía coherencia de patrones se retuvieron los datos para análisis posteriores. Las repeticiones de cada tratamiento fueron consideradas en forma individual, para evitar la hipótesis de homogeneidad física. Finalmente, el modelo expo–lineal fue ajustado a través de un proceso de minimización no lineal del error cuadrático de estimación y sólo las curvas con dos o más datos en la fase lineal fueron analizadas. El usar toda las mediciones, destructivas y a nivel de plantas individuales, implica una hipótesis de homogeneidad física (estacionariedad en términos estadísticos) que difícilmente puede ser sostenida. No obstante lo anterior, si se utilizan todas las mediciones disponibles, los resultados no se alteran y, por el contrario, muestran mejores ajustes a los mostrados.
El Cuadro 2 muestra los resultados del ajuste del modelo expo–lineal para la materia aérea seca (g planta–1) y área foliar (cm2 planta–1). En general, el ajuste del modelo expo–lineal fue bueno (cuando había datos suficientes) ante todas las condiciones de estrés de humedad y de N analizadas, por lo que este modelo se puede usar en forma robusta y confiable. Para evaluar el ajuste del modelo se usó la raíz del error cuadrático medio (RECM), que mide el error de estimación en términos absolutos.
En la Figura 6a, se muestra el peor ajuste del modelo exponencial para la materia aérea seca y la Figura 6b, muestra lo mismo para el área foliar. Como se observa, los ajustes muestran que el patrón del modelo expo–lineal es representativo del crecimiento en la etapa vegetativa de la planta. Resultados similares a los mostrados se obtuvieron de la materia fresca aérea y la materia total, seca y fresca.
Equivalencia de la productividad del tomate
Usando la fase lineal del modelo expo–lineal se estimaron las constantes ak (intersección) y bk (pendiente) de las rectas, para analizar sus patrones y compatibilidad con el concepto de la equivalencia de la productividad (medio ambiente fijo). En la Figura 7, se muestran los resultados de los patrones entre ak y bk para el caso de la materia seca aérea y área foliar. Es importante señalar que los resultados obtenidos usando todos los datos (no mostrados), sin eliminar inconsistencias, tienen una R2 mayor que los presentados.
La Figura 7 muestra que el modelo de equivalencia ambiental se cumple, indicando que la tasa de crecimiento relativo R puede considerarse constante y el efecto del estrés se manifiesta en el tiempo de transición dado por la Ecuación 5; es decir, donde el cultivo alcanza su máxima tasa de crecimiento, evidenciándose en los valores de tb o tiempo perdido (Monteith, 2000).
CONCLUSIONES
– La aplicación del modelo expo–lineal al caso de un experimento de tomate cultivado en invernadero con diferentes combinaciones de niveles de estrés hídrico y por nitrógeno mostró buenos ajustes a los datos experimentales, fundamentando su uso en el caso general de condiciones alejadas de un óptimo (no estrés).
– En el caso del concepto de la equivalencia ambiental, los resultados para el caso de un ambiente fijo (invernadero) son compatibles con su formulación.
LITERATURA CITADA
Ferrandino, F. J. 1989. Spatial and temporal variation of a defoliating plant disease and reduction in yield. Agric. For. Meteorol. 47: 273–289. [ Links ]
Gijzen, H. E., E. Heuvelink, H. Challa, E. Dayan. L. Marcelis, S. Cohen, and M. Fuchs. 1998. HORTSIM: a model for greenhouse crop and greenhouse climate. Acta Hortic. 456: 441–449. [ Links ]
Goudriaan, J. 1994. Using the expolinear growth equation to analize resource capture. pp. 99–110. In: J. L. Monteith, R. K. Scott, and M. H. Unsworth (eds.). Resource Capture by Crops. Nottingham University Press. Nottingham, UK. [ Links ]
Goudriaan, J. and J. L. Monteith. 1990. A mathematical function for crop growth based on light interception and leaf area expansion. Anal. Bot. 66: 695–701. [ Links ]
Heuvelink, E. 1995. Dry matter production in a tomato crop: measurements and simulation. Ann. Bot. 75: 369–379. [ Links ]
Huete, A. R., R. D. Jackson, and D. F. Post. 1985. Spectral response of a plant canopy with different soil backgrounds. Remote Sens. Environ. 17: 35–53. [ Links ]
Hunt, R. 1982. Plant growth curves. Edward Arnold. London, UK. [ Links ]
Jones, J., E. Dayan, L. Allen, K. H. Van, and H. Challa. 1991. A dynamic tomato growth and yield model (TOMGRO). Transactions of the ASAE 34: 633–673. [ Links ]
Lieth, J. H., P. R. Fisher, and R. D. Heins. 1996. A phasic model for the analysis of sigmoid patterns of growth. Acta Hortic. 417: 113–118. [ Links ]
Monteith, J. L. 1972. Solar radiation and productivity in tropical ecosystems. J. Appl. Ecol., 9: 747–766. [ Links ]
Monteith, J. L. 1981. Does light limit crop production? pp. 23–38. In: C. B. Johnson (ed.), physiological processes limiting plant productivity. Butterworths, London, U.K. [ Links ]
Monteith, J. L. 2000. Fundamentals equations for growth in uniform stands of vegetation. Agric. For. Meteorol. 104: 5–11. [ Links ]
Ross, J. K. 1981. The radiation regime and architecture of plant stands. Dr. W. Junk Publishers. Norwell, MA, USA. [ Links ]
Steiner, A. A. 1984. The universal nutrient solution. pp. 633–650. In: Proccedings of the 6th International Congress on soilles culture. Wageningen, The Netherlands. [ Links ]
Yin, X., J. Goudriaan, E. A. Latinga, J. Vos, and H. J. Spiertz. 2003. A flexible sigmoid function of deterministic growth. Ann. Bot. 91: 361–371. [ Links ]
Yuan, F. M. and W. Bland. 2004. Light and temperature modulated expolinear growth model for potato (Solanum tuberosum L.). Agric. For. Meteorol. 121: 141–151. [ Links ]

