










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkTerra Latinoamericana
On-line version ISSN 2395-8030Print version ISSN 0187-5779
Terra Latinoam vol.26 n.2 Chapingo Apr./Jun. 2008
División III
Sucesión en el matorral desértico de Larrea tridentata (DC.) Cov. en la Sierra de Catorce, San Luis Potosí, México*
Succession in the Microphyll Desert Scrub of Larrea tridentata (DC.) Cov. in the Sierra de Catorce, San Luis Potosi, Mexico
Jesús Ugalde Ávila1, Diódoro Granados-Sánchez2‡ y Arturo Sánchez-González3
1 Universidad Nacional Autónoma de México. México, D. F.
2 División de Ciencias Forestales, Universidad Autónoma Chapingo. 56230 Chapingo, Estado de México. ‡Autor responsable (didorog@latinmail.com)
3 Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo. 42184 Pachuca, Hidalgo, México.
Recibido: septiembre de 2006.
Aceptado: diciembre de 2007.
RESUMEN
Se estudió el proceso de regeneración del matorral desértico micrófilo (MDM) dominado por Larrea tridentata (DC.) Cov. en la Sierra de Catorce, San Luis Potosí, México. Para el trabajo de campo se seleccionaron siete sitios de muestreo afectados por actividades agrícolas desde uno hasta 50 años. En cada sitio se establecieron cinco parcelas de 4 m2 para determinar la composición y la densidad de las especies arbustivas y herbáceas; adicionalmente, se cuantificaron algunas variables edáficas. Los métodos de crono-secuencias y de análisis multivariable permitieron reconocer que en el MDM el proceso de sucesión ecológica no ocurre en el sentido clásico, en el que ensambles más o menos definidos de especies son sustituidas por otros hasta formar una comunidad madura. En su lugar ocurrió una autosucesión: la mayoría de las especies colonizadoras fueron dominantes en todos los sitios de la crono-secuencia analizada y en la comunidad madura. La composición de especies y el tiempo de recuperación post-disturbio se correlacionó con cambios en las características edáficas, principalmente en los valores del potencial de hidrógeno, la materia orgánica y los cationes del suelo. L. tridentata y otras especies estructuralmente importantes mostraron características tanto de especies pioneras como clímax en la crono-secuencia, lo cual sugiere que las interacciones bióticas son muy importantes en el tiempo de regeneración del MDM en las zonas semiáridas de México.
Palabras clave: zonas semiáridas, ordenación, crono-secuencias, autosucesión, gobernadora.
ABSTRACT
In the Sierra de Catorce, San Luis Potosi, Mexico, the regeneration process of microphyll desert scrub (MDS) dominated by Larrea tridentata (D.C.) Cov. was studied. Seven sample sites were selected for the field work. These sites had been affected by agricultural activities from one to 50 years. In each site five 4 m2 plots were established in which composition and density of the shrub and herbaceous species were determined. Furthermore, some edaphic variables were quantified. Chrono-sequential and multivariate analysis suggested that in the MDS the ecological succession process does not occur in the classical sense in which more or less definite species ensembles are replaced by other ensembles until a mature community is shaped. Instead, auto-succession occurred: most of the colonizing species were dominant in all the plots of the analyzed chrono-sequence and in the mature community. Species composition and recovery time after disturbance were correlated with changes in edaphic characteristics, mainly within the hydrogen potential values, organic matter, and soil cations. L. tridentata and other structurally important species showed characteristics of both pioneer and climax species in the chrono-sequence, which suggests that biotic interactions are important in the regeneration time of MDS in the semiarid areas of Mexico.
Keywords: semiarid lands, ordination, chronosequences, auto-succession, creosote bush.
INTRODUCCIÓN
La vegetación de las regiones de clima seco de México se conoce como matorral xerófilo (Rzedowski, 1978), pero dentro de esta denominación general se incluyen numerosos tipos de vegetación, entre los que destaca el matorral desértico micrófilo, propio de terrenos planos y de las partes inferiores de los cerros de una gran zona del altiplano potosino-zacatecano, en el desierto chihuahuense (Miranda, 1964; Marroquín et al., 1981; Granados-Sánchez y Sánchez-González, 2003). En el matorral desértico micrófilo predominan los elementos arbustivos de hoja pequeña que incluyen casi siempre a Larrea tridentata y Flourensia cernua (Rzedowski, 1978). L. tridentata "La Gobernadora" es un arbusto de 2 a 3 m de altura que se distribuye en forma casi ininterrumpida desde Nevada, Utah, Nuevo México y Texas (en Estados Unidos) hasta Guanajuato, Querétaro e Hidalgo (en México) donde es una de las especies más abundantes y conspicuas (Rzedowski y Calderón, 1988).
Grandes extensiones (más de 100 000 ha anuales en las últimas cuatro décadas) de los matorrales de las zonas muy secas y secas de México están experimentando procesos de sucesión secundaria, debido, en gran parte, al abandono continuo y extensivo de las tierras de cultivo (SNIARN, 2005); ello provee de una excelente oportunidad para estudiar este proceso de cambio en la composición y dinámica de la vegetación. En forma tradicional, se considera que la sucesión secundaria en las zonas secas es muy lenta: se necesitan décadas o aun siglos para restablecer las relaciones naturales entre los recursos del suelo y la vegetación, donde las condiciones ambientales imperantes son la causa principal (Vasek, 1979/1980; Webb et al., 1987; Tilman 1988; Bonet, 2004; Miranda et al., 2004). Sin embargo, la sucesión en estos ecosistemas tiene múltiples vías de recuperación relacionados con factores locales y regionales (Focht y Pillar, 2003; Pugnaire et al., 2004; Armas y Pugnaire, 2005), por lo que no es, en todos los casos, un proceso lento (Shreve, 1929; Costello, 1944; MacMahon, 1981; Foster y Motzkin, 2003).
La variación espacial y temporal en la composición de especies puede ser considerada análoga (Lehman y Tilman 2000; Alados et al., 2005), de ahí que el remplazo del espacio por el tiempo tenga el objeto de formular modelos descriptivos e hipótesis acerca de los mecanismos que determinan los cambios durante la sucesión vegetal. En la presente investigación se analizó la sucesión secundaria en el MDM de L. tridentata con el esquema de crono-secuencias (Pickett, 1989), en tierras de cultivo abandonadas en distintos tiempos. Se planteó como hipótesis que es posible reconstruir los procesos temporales de cambio en la composición de especies en el matorral desértico micrófilo a partir del análisis del mosaico espacial en un medio semejante con tiempos de recuperación post-disturbio diferentes. El objetivo principal de este trabajo fue contribuir al conocimiento de la sucesión y, por ende, de la regeneración ecológica en el matorral desértico micrófilo de México.
MATERIALES Y MÉTODOS
La Sierra de Catorce se localiza entre 23° 24' y 23° 47' N y entre 100° 46' y 100° 57' O, en la zona noreste del altiplano potosino-zacatecano (dentro del desierto chihuahuense) y forma parte de las sierras inferiores de la provincia fisiográfica de la Sierra Madre Oriental (Barboza-Gudiño et al., 2004). El intervalo altitudinal en la Sierra de Catorce varía de 1800 m (en el valle de Wadley) a 3060 m (en la cima del cerro del Barco). Presenta un clima semíseco templado, con lluvias de verano, con porcentaje de precipitación invernal mayor de 10.2 mm (la precipitación media anual es de 306.4 mm) y es extremoso, con oscilación de la temperatura media mensual entre 7 y 14 °C; la temperatura media anual es de 17.34 °C (Zárate del Valle, 1982; García et al., 1985). En la Sierra de Catorce se encuentran expuestas rocas sedimentarias, ígneas y metamórficas, cuyas edades pertenecen al Triásico, Jurásico y Cretácico (Zárate del Valle, 1982; Barboza-Gudiño et al., 2004). Debido a los procesos de intemperización, transportación y sedimentación, los suelos en el área son, en general, de poca profundidad. Los perfiles más desarrollados son los del grupo de los Xerosoles que se caracterizan por la presencia de sales; este suelo se presenta en las partes bajas de las cuencas donde el drenaje endorreico y la acumulación de material arcilloso provocan la deficiencia en la filtración e impiden la lixiviación de los perfiles. Otros tipos de suelo importantes por su extensión son los Litosoles con fase petrocálcica que se encuentran relacionados estrechamente a elevaciones, donde el material rocoso está cerca de la superficie o se encuentra aflorando (Hernández, 1988). Los tipos de vegetación más importantes son: matorral desértico micrófilo, matorral desértico crasicaule, matorral desértico rosetófilo, encinar arbustivo y piñonar (Sánchez-González y Granados-Sánchez, 2003).
Para analizar la sucesión secundaria en el matorral desértico micrófilo de L. tridentata se seleccionaron seis sitios de muestreo, con base en el tiempo de abandono después de la actividad agrícola. La información fue proporcionada por los campesinos quienes utilizaron el matorral para el cultivo de maíz. Los sitios elegidos fueron de uno, tres, siete, 10, 20 y 40 años post-cultivo. Como comunidad madura o clímax se eligió un sitio sin señales de perturbación evidentes.
En cada sitio se colocaron, al azar, cinco parcelas de muestreo de 4 m2, en las que se estimó la densidad, la cobertura y la frecuencia de especies arbustivas y herbáceas. La suma de los valores relativos de los tres variables anteriores permitió estimar el valor de importancia relativa (VIR) de las especies (Matteucci y Colma, 1982). Los ejemplares recolectados se depositaron en el herbario de la División de Ciencias Forestales de la Universidad Autónoma Chapingo. Por sitio de muestreo se determinó la altitud, la pendiente, la exposición, la pedregosidad y las siguientes características edáficas: potencial de hidrógeno (pH), materia orgánica, porcentaje de arena, limo y arcilla, densidad aparente, nitrógeno total, potasio y sodio intercambiables (Cuadro 1). La altitud (1780 a 1830 m), pendiente (0 a 10%) y orientación de los sitios de muestreo se consideraron como constantes.
La semejanza en la composición de especies entre sitios y la relación entre las especies y las variables ambientales en la crono-secuencia se estimó mediante el análisis de correspondencia canónica, con el programa de cálculo PC-ORD, Versión 4.10 (McCune y Grace, 2002).
RESULTADOS Y DISCUSIÓN
Se colectaron en total 34 especies, las más abundantes y conspicuas. La riqueza de especies por sitio incrementó respecto al tiempo de recuperación: la riqueza más baja se registró en los sitios con poco tiempo de recuperación y la más alta en los sitios con más de 10 años de recuperación. El sitio considerado como comunidad madura o clímax presentó el mayor número de especies (Cuadro 2). Otras investigaciones mencionan que la edad post-cultivo del sitio se correlaciona positivamente con la riqueza y abundancia de las especies (Wali, 1999). En ecosistemas mediterráneos secos de España se ha documentado que la recuperación gradual de la diversidad de especies se alcanzó después de 30 años de abandono de las tierras agrícolas (Alados et al., 2005). Cuando el disturbio en el suelo no es muy severo, el tiempo de retorno a la comunidad madura en estos ecosistemas se ha estimado en 50 años o menos (Costello, 1944).
Conforme transcurre la sucesión secundaria en el matorral desértico micrófilo de la Sierra de Catorce, el número de especies y la densidad de individuos por especie incrementan progresivamente hasta que, en las tierras de cultivo con 20 ó más años de abandono, la recuperación natural se completa con la presencia constante de las especies estructuralmente importantes en todas las parcelas. Foster y Motzkin (2003) mencionan que en ecosistemas áridos de Nueva Inglaterra, en EE. UU., la tasa de recuperación de las especies perennes está influenciada por el tipo de suelo, el tipo de agricultura y por las características de las especies de la sucesión.
La composición de especies después de una perturbación está definida, en gran medida, por las especies que existían antes de la alteración y por la vegetación circundante (Egler, 1954; Milton, 1995; Tirado y Pugnaire, 2005); el hecho de que las especies importantes en la estructura del matorral desértico micrófilo sean constantes a través de la crono-secuencia analizada confirma que la "autosucesión" es un fenómeno común en zonas secas (Shreve, 1929; Muller, 1940; MacMahon, 1981). La sucesión secundaria en estas zonas no implica, necesariamente, el remplazo de unas especies por otras; las especies que componen la comunidad madura están presentes desde el primer año y la invasión por otras especies es muy limitada (Shreve, 1929; Hanes, 1971; Miranda et al., 2004).
Mientras que algunos estudios en zonas muy secas de Norteamérica mencionan una tasa de incorporación lenta y colonización tardía (Webb et al., 1987; McAuliffe, 1991; Bowers et al., 1997) para L. tridentata, otros (Shreve, 1942; Vasek, 1979/1980) coinciden en que las plántulas de la gobernadora aparecen desde el inicio de la sucesión. En el presente trabajo L. tridentata fue constante en todos los sitios de la crono-secuencia, lo que demuestra que tiene características tanto de especie serial inicial como serial tardía y que es, además, una de las mejor adaptadas a las condiciones del medio seco, con un intervalo de distribución muy amplio comparado con el de otras especies (Muller, 1940; Reynolds, 1986).
En las parcelas de muestreo varias especies se presentaron en conjunto con L. tridentata; destacan por su constancia: Atriplex canescens, Dyssodia setifolia, Euphorbia thymifolia, Flourensia cernua, Opuntia cantabrigiensis, O. leptocaulis, O. microdasys, Parthenium incanum, Zaluzania robinsonii y Zinnia acerosa (Cuadro 2). Las especies con mayor VIR fueron Euphorbia thymifolia, L. tridentata, Parthenium incanum y Zaluzania robinsonii con casi el 50% del total (Cuadro 3); la única especie arbustiva de las cuatro L. tridentata alcanzó hasta el 90% de la cobertura total en los sitios con 20 o más años de recuperación.
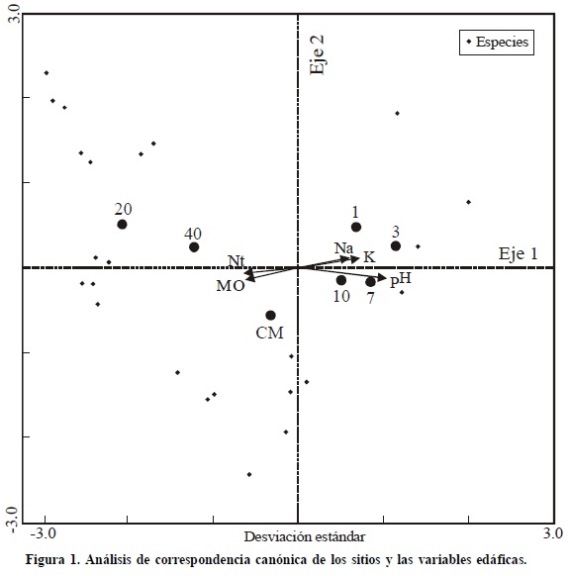
El valor de las raíces características (Figura 1, Cuadro 4) muestra que sólo el primer eje de la ordenación se relaciona significativamente (P < 0.05) con la variación en la composición de especies entre los sitios de la crono-secuencia. En el diagrama de la ordenación de los sitios, las especies y los factores edáficos se observa que los sitios con tiempos de uno, tres, siete y 10 años son más semejantes entre sí que los sitios con 20 ó más años de recuperación. Este patrón, definido con datos de presencia-ausencia de las especies en los sitios de muestreo, sugiere un remplazo en la composición de especies conforme transcurre la sucesión. Sin embargo, los datos del Cuadro 2 muestran que ocho de las 12 especies consideradas como seriales tempranas, por estar presentes en los sitios con uno y tres años de recuperación: Atriplex canescens, Dyssodia setifolia, Euphorbia thymifolia, Flourensia cernua, Larrea tridentata, Parthenium incanum, Zaluzania robinsonii y Zinnia acerosa, se presentaron también en las etapas seriales intermedias/tardías y son, además, las que definen la estructura del matorral desértico micrófilo (VIR: 65%; Cuadro 3). Estos resultados sugieren que la composición florística inicial (sensu Egler, 1954), después de un disturbio, puede jugar un papel determinante en el tiempo de recuperación de la comunidad madura (Figura 2).

La distribución de las plantas en el espacio y en el tiempo está relacionada y depende de factores bióticos y abióticos múltiples (Whittaker, 1967; Focht y Pillar, 2003; James et al., 2005).
Aunque es complejo distinguir, por su naturaleza dinámica e interactiva, los efectos de las características físicas y químicas del suelo sobre la composición y diversidad de especies (Schoenholtz et al., 2000), los resultados del análisis de correspondencia canónica muestran que la variación en la composición de especies durante la sucesión en el matorral desértico micrófilo de L. tridentata se relaciona positivamente con la cantidad de materia orgánica y el nitrógeno (N) total, y negativamente con el pH, el potasio y el sodio intercambiables. La correlación entre las variables edáficas analizadas y el primer eje de la ordenación fue alta en todos los casos (Cuadro 4). Algunas investigaciones en sistemas desérticos indican que cuando el suministro de agua para las plantas es adecuado, el N y el P pueden convertirse en factores limitantes (García-Moya y McKell, 1970; Smith et al., 1997; Drenovsky y Richards, 2004; James et al., 2005; James y Richards, 2006; Perkins et al., 2006).
Perturbaciones como las que resultan de la actividad agrícola, al modificar la disponibilidad de nutrimentos y la humedad del suelo, pueden causar un alto impacto sobre la vegetación. Con la pérdida de la cobertura arbustiva los suelos desnudos son susceptibles a la erosión por agua y viento, y la remoción neta de los nutrimentos de los parches degradados aumenta. En este escenario adquiere importancia el proceso de auspicio que involucra, entre otras cosas, el incremento en la disponibilidad de agua y nutrimentos (Perkins et al., 2006). El suelo bajo las plantas nodrizas tiene un contenido más alto en nutrimentos "islas de fertilidad" (sensu García-Moya y McKell, 1970) que el de las áreas abiertas circundantes y, de esta manera, induce un incremento relativo en el crecimiento de las plántulas (Holmgren et al., 1997; Pugnaire et al., 2004; James y Richards, 2006; Perkins et al., 2006). En medios semisecos las interacciones positivas y negativas entre especies vegetales contribuyen a la formación de micro-sitios donde ocurren sucesiones cíclicas (Yeaton, 1978); el balance entre ambos tipos de interacciones puede cambiar en intensidad y signo a través del tiempo y el espacio, dependiendo de la disponibilidad de recursos y de las historias de vida de las plantas (Armas y Pugnaire, 2005; James y Richards, 2006).
CONCLUSIONES
La variación en la composición de especies entre los sitios de muestreo está relacionada con el tiempo de abandono de los terrenos después de la actividad agrícola. La crono-secuencia analizada sugiere que la regeneración natural del matorral desértico micrófilo se inicia desde el primer año, pero es evidente hasta los siete años, cuando la mayoría de las especies presentes persisten hasta la comunidad madura. Como los sitios con 20, 40 ó más años de recuperación no muestran cambios significativos en composición y estructura, es probable que la restauración ocurra después de los 20 años, tiempo alrededor del cual los valores de las características físicas y químicas del suelo (potencial de hidrógeno, la materia orgánica y los cationes) son los óptimos. En la comunidad vegetal analizada la sucesión no ocurrió en el sentido clásico estricto, en su lugar ocurrió una autosucesión: la mayoría de las especies pioneras se presentaron en etapas posteriores de la crono-secuencia y en la comunidad madura. Es probable que la reposición de las especies características de la comunidad madura ocurra a través de cambios cíclicos a lo largo del tiempo, en los que Larrea tridentata probablemente desempeñe un papel relevante como planta nodriza.
AGRADECIMIENTOS
A Don Ceferino Charcas e hijos, campesinos que amablemente proporcionaron la información sobre el tiempo de abandono de las tierras después de cultivo. A los revisores y el editor, por su contribución para mejorar la estructura y el contenido del manuscrito.
LITERATURA CITADA
Alados, C. L., Y. Pueyo, D. Navas, B. Cabezudo, A. González, and D. C. Freeman. 2005. Fractal analysis of plant spatial patterns: a monitoring tool for vegetation transition shifts. Biodiversity Conserv. 14: 1453-1468. [ Links ]
Armas, C. and F. I. Pugnaire. 2005. Plant interactions govern population dynamics in a semi-arid plant community. J. Ecol. 93: 978-989. [ Links ]
Barboza-Gudiño, J. F., M. Hoppe, M. Gómez-Anguiano y P. R. Martínez-Macías. 2004. Aportaciones para la interpretación estratigráfica y estructural de la porción noroccidental de la Sierra de Catorce, San Luís Potosí, México. Revista Mexicana de Ciencias Geológicas 21: 299-319. [ Links ]
Bonet, A. 2004. Secondary succession of semi-arid Mediterranean old-fields in south-eastern Spain: insights for conservation and restoration of degraded lands. J. Arid Environ. 56: 213-233. [ Links ]
Bowers, J. E., R. H. Webb, and E. A. Pierson. 1997. Succession of desert plants on debris flow terraces, Grand Canyon, Arizona, U.S.A. J. Arid Environ. 36: 67-86. [ Links ]
Costello, D. F. 1944. Natural revegetation of abandoned plowed land. Ecology 25: 312-326. [ Links ]
Drenovsky, R. E. and J. H. Richards. 2004. Critical N:P values predicting nutrient deficiencies in desert shrublands. Plant Soil 259: 59-69. [ Links ]
Egler, F. E. 1954. Vegetation science concepts. 1. Initial floristic composition - a factor in old-field vegetation development. Vegetatio 4: 412-417. [ Links ]
Focht, T. and V. D. Pillar. 2003. Spatial patterns and relations with site factors in a campos grassland under grazing. Brazilian J. Biol. 63: 423-436. [ Links ]
Foster, D. R. and G. Motzkin. 2003. Interpreting and conserving the openland habitats of coastal New England: insights from landscape history. For. Ecol. Manage. 185: 127-150. [ Links ]
García, E., R. Vidal y C. M. E. Hernández. 1985. Aspectos climáticos de las zonas áridas del norte de la altiplanicie mexicana. Boletín del Instituto de Geografía de la UNAM 15: 41-74. [ Links ]
García-Moya, E. and C. M. McKell. 1970. Contribution of shrubs to the nitrogen economy of a desert-wash plant community. Ecology 51: 81-88. [ Links ]
Granados-Sánchez, D. y A. Sánchez-González. 2003. Clasificación fisonómica de la vegetación de la Sierra de Catorce, San Luis Potosí, a lo largo de un gradiente altitudinal. Terra 21: 321-332. [ Links ]
Hanes, T. L. 1971. Succession after fire in the chaparral of Southern California. Ecol. Monogr. 41: 27-49. [ Links ]
Hernández, S. 1988. Caracterización de suelos y aguas y sus posibles usos en el área del Salado, Estados de Zacatecas y San Luis Potosí. Geografía y Desarrollo 1: 19-26. [ Links ]
Holmgren, M., M. Scheffer, and M. A. Huston. 1997. The interplay of facilitation and competition in plant communities. Ecology 78: 1966-1975. [ Links ]
James, J. J. and J. H. Richards. 2006. Plant nitrogen capture in pulse-driven systems: interactions between root responses and soil processes. J. Ecol. 94: 765-777. [ Links ]
James, J. J., R. L. Tiller, and J. H. Richards. 2005. Multiple resources limit plant growth and function in a saline-alkaline desert community. J. Ecol. 93: 113-126. [ Links ]
Lehman, C. L. and D. Tilman. 2000. Biodiversity, stability, and productivity in competitive communities. Am. Nat. 156: 534-552. [ Links ]
MacMahon, J. A. 1981. Successional processes: comparisons among biomes with special reference to probable roles of and influences on animals. pp. 277-304. In: D. C. West, H. H. Shugart, and D. B. Botkin (eds.). Forest succession. Springer-Verlag. New York, NY, USA. [ Links ]
Marroquín, S. J., G. L. Borja, R. C. Velázquez y C. J. A. Cruz. 1981. Estudio ecológico dasonómico de las zonas áridas del norte de México. Publicación especial 2. Instituto Nacional de Investigaciones Forestales. México, D. F. [ Links ]
Matteucci, S. D. y A. Colma. 1982. Metodología para el estudio de la vegetación. Serie Biología, Monografía 22. Secretaría General de la Organización de los Estados Americanos, Programa Regional de Desarrollo Científico y Tecnológico. Washington, DC, USA. [ Links ]
McAuliffe, J. R. 1991. Demographic shifts and plant succession along a late Holocene soil chronosequence in the Sonoran Desert of Baja California. J. Arid Environ. 20: 165-178. [ Links ]
McCune, B. and J. B. Grace. 2002. Analysis of ecological communities. MjM Software Design. Gleneden Beach, OR, USA. [ Links ]
Milton, S. J. 1995. Spatial and temporal patterns in the emergence and survival of seedlings in arid Karoo shrubland. J. Appl. Ecol. 32: 145-156. [ Links ]
Miranda, F. H. E. 1964. Las zonas áridas del centro y noroeste de México y el aprovechamiento de sus recursos. Instituto Mexicano de Recursos Naturales Renovables. México, D. F. [ Links ]
Miranda, J. D., F. M. Padilla y F. I. Pugnaire. 2004. Sucesión y restauración en ambientes semiáridos. Ecosistemas URL: http://www.aeet.org/ecosistemas/041/investigacion4.htm (Consulta: julio 15, 2006).
Muller, C. H. 1940. Plant succession in the Larrea-Flourensia climax. Ecology 21: 206-212. [ Links ]
Perkins, S. R., K. C. McDaniel, and A. L. Ulery. 2006. Vegetation and soil change following creosotebush (Larrea tridentata) control in the Chihuahuan Desert. J. Arid Environ. 64: 152-173. [ Links ]
Pickett, S. T. A. 1989. Space-for-time substitution as an alternative to long term studies. pp. 110-135. In: G. E. Likens (ed.). Long term studies in Ecology. Springer-Verlag. New York, NY, USA. [ Links ]
Pugnaire, F. I., C. Armas, and F. Valladares. 2004. Soil as a mediator in plant-plant interactions in a semi-arid community. J. Vegetation Sci. 15: 85-92. [ Links ]
Reynolds, J. 1986. Adaptative strategies of desert shrubs with special reference to the creosotebush. pp. 63-71. In: Pattern and process in desert ecosystems. University of New Mexico Press. Albuquerque, NM, USA. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa. México, D. F. [ Links ]
Rzedowski, J. y G. Calderón de Rzedowski. 1988. Dos nuevas localidades de Larrea tridentata (Zygophyllaceae) en el Centro de México y su interés fitogeográfico. Acta Bot. Mex. 1: 7-9. [ Links ]
Sánchez-González, A. y D. Granados-Sánchez. 2003. Ordenación de la vegetación de la Sierra de Catorce a lo largo de gradientes ambientales. Terra 21: 311-319. [ Links ]
Schoenholtz, S. H., H. V. Miegroet, and J. A. Burger. 2000. A review of chemical and physical properties as indicators of forest soil quality: challenges and opportunities. For. Ecol. Manage. 138: 335-356. [ Links ]
Shreve, F. 1929. Changes in desert vegetation. Ecology 10: 364-373. [ Links ]
Shreve, F. 1942. The desert vegetation of North America. Bot. Rev. 8: 195-246. [ Links ]
Smith, S. D., R. K. Monson, and J. E. Anderson. 1997. Physiological ecology of North American desert plants. Springer. Berlin, Germany. [ Links ]
SNIARN (Sistema Nacional de Información Ambiental y de Recursos Naturales). 2005. Informe de la situación del medio ambiente en México. Compendio de Estadísticas Ambientales. Secretaría de Medio Ambiente y Recursos Naturales. México, D. F. [ Links ]
Tilman, D. 1988. Plant strategies and the dynamics and structure of plant communities. Princeton University Press. Princeton, NJ, USA. [ Links ]
Tirado, R. and F. I. Pugnaire. 2005. Community structure and positive interactions in constraining environments. Oikos 111: 437-444. [ Links ]
Vasek, F. C. 1979/1980. Early successional stages in Mojave Desert scrub vegetation. Israel J. Bot. 28: 133-148. [ Links ]
Wali, M. K. 1999. Ecological succession and the rehabilitation of disturbed terrestrial ecosystems. Plant Soil 213: 195-220. [ Links ]
Webb, R. H., J. W. Steiger, and R. M. Turner. 1987. Dynamics of Mojave Desert shrub assemblages in the Panamint mountains, California. Ecology 68: 478-490. [ Links ]
Whittaker, R. H. 1967. Gradient analysis of vegetation. Biol. Rev. 42: 207-264. [ Links ]
Yeaton, R. I. 1978. A cyclical relationship between Larrea tridentata and Opuntia leptocaulis in the Northern Chihuahuan desert. J. Ecol. 66: 651-656. [ Links ]
Zarate del Valle, P. F. 1982. Geología y análisis metalogénico de la Sierra de Catorce, S. L. P. Boletín de la Sociedad Geológica Mexicana 43: 1-21. [ Links ]
* Publicado en Terra Latinoamericana 26: 153–160.

