










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkTerra Latinoamericana
On-line version ISSN 2395-8030Print version ISSN 0187-5779
Terra Latinoam vol.26 n.1 Chapingo Jan./Mar. 2008
División I
Nota de investigación
Variación de la velocidad de infiltración media en seis ecosistemas inalterados*
Variation of the Mean Infiltration Rate in Six Unperturbed Ecosystems
Martha Alejandra García–Hernández1, Mónica Alejandra García–Hernández1, Iván Castellanos–Vargas1, Zenón Cano–Santana1‡ y Cinthya M. Peláez–Rocha1
1 Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México. Ciudad Universitaria. 04510 México, D. F. ‡ Autor responsable (zcs@hp.fciencias.unam.mx)
Recibido: septiembre de 2005.
Aceptado: junio de 2007.
RESUMEN
Se midió la velocidad de infiltración media (VIM) de agua en el suelo de seis ecosistemas de la Reserva de la Biosfera Sierra Gorda. En el bosque de pino, adicionalmente, se observó la VIM en sitios con mantillo y sitios con la capa de mantillo removida vertiendo 100 mL de agua en tubos de PVC de 10.16 cm de diámetro. La profundidad de mantillo varió de 4 a 5 cm en los bosques de pino y de encino, así como en la selva baja, mientras que en el pastizal y en los matorrales xerófilos esta capa estaba ausente. Se registró 31.30 ± error estándar 4.02 mL cm–2 min–1 en el bosque de pino con mantillo, 21.02 ± 3.20 mL cm–2 min–1 en el bosque de encino, 20.55 ± 2.25 mL cm–2 min–1 en el pastizal, 15.45 ± 2.60 mL cm–2 min–1 en la selva baja caducifolia, 7.74 ± 1.70 mL cm–2 min–1 en el matorral xerófilo dominado por Larrea tridentata, 6.01 ± 0.80 mL cm–2 min–1 en el bosque de pino con mantillo removido y 2.61 ± 0.73 mL cm–2 min–1 en el matorral xerófilo de Acacia constricta. Se concluyó que la presencia del mantillo es uno de los factores determinantes que incrementan la VIM de agua en el ecosistema. Otros factores que parecen afectar la tasa de infiltración son: la cercanía a los cuerpos de agua, la textura edáfica y la edad del ecosistema.
Palabras clave: mantillo, Sierra Gorda.
ABSTRACT
The mean infiltration rate (MIR) of water in the soil of six ecosystems from the Reserva de la Biosfera Sierra Gorda was measured. Additionally, in pine forest the MIR was observed in sites with litter and in sites where litter was removed that emitted 100 mL water in tubes of 10.16 cm diameter. Litter depth varied from 4 to 5 cm in the pine and oak forest, as well as in the low tropical forest, while in the grassland and xeric scrub this layer was absent. We registered 31.30 ± standard error 4.02 mL cm–2 min–1 in pine forest with litter, 21.02 ± 3.20 mL cm–2 min–1 in oak forest, 20.55 ± 2.25 mL cm–2 min–1 in grassland, 15.45 ± 2.60 mL cm–2 min–1 in dry forest, 7.74 ± 1.70 mL cm–2 min–1 in Larrea tridentata xeric scrub, 6.01 ± 0.80 mL cm–2 min–1 in pine forest without litter and 2.61 ± 0.73 mL cm–2 min–1 in Acacia constricta xeric scrub. The presence of litter is a determinant factor that increases the water infiltration rate in the ecosystem. Other factors that seem to affect the MIR are: the proximity of water bodies, the edaphic texture and ecosystem age.
Keywords: litter, Sierra Gorda.
INTRODUCCIÓN
La mayor parte del agua que las plantas utilizan para sus procesos fisiológicos proviene del suelo y con frecuencia el exceso o déficit de ella suele actuar como un factor limitante de su crecimiento.
La velocidad de infiltración es el proceso que define la entrada y el movimiento, en general vertical, del agua a través de la superficie del suelo en una unidad de tiempo definida (cmmin–1 o bien, cmh–1) (Tamhane et al., 1978; Porta et al., 1994; Cerdá, 1996). La velocidad a la que el agua penetra en el suelo es, en general, elevada al inicio de la infiltración debido a que el gradiente matricial de succión al inicio es elevado en las capas superficiales en particular cuando el suelo está seco, y esta velocidad tiende a decrecer a medida que transcurre el tiempo (Buol et al., 1981; Porta et al., 1994). Por su parte, la percolación es el movimiento del agua a través de un perfil edáfico que comprende tres direcciones en el flujo de ésta (vertical, horizontal y radial) (Osuna y Padilla, 1998). El agua que no puede penetrar se desplaza por la superficie y a menudo provoca erosión edáfica, dando como resultado una pérdida de la fertilidad y la disminución del aporte de agua al suelo (Sarukhán y Maass, 1990; Osuna y Padilla, 1998). La relación entre la velocidad de infiltración y la precipitación determina la cantidad de agua que penetra al suelo y la que se pierde por escorrentía (Tamhane et al., 1978; Osuna y Padilla, 1998; Návar y Synnott, 2000; Franzluebbers, 2001). Un desbalance en esta relación puede ocasionar el escurrimiento excesivo de agua lo que provoca inundaciones en tierras bajas (Sarukhán y Maass, 1990).
La velocidad de infiltración es afectada por las características del perfil edáfico, como la textura, la estructura, la cantidad de materiales orgánicos e inorgánicos, así como por el tamaño y volumen total del espacio poroso (Tamhane et al., 1978; Gurovich, 1985). Daubenmire (1979), Buol et al. (1981) y Gregory et al. (1987) discuten que la presencia de raíces de las plantas es un factor determinante para incrementar el espacio poroso del suelo y, a su vez, aumentar la permeabilidad. Según estos autores, más de la mitad del volumen de un suelo es espacio poroso y eso es principalmente propiciado por la fase de intercambio entre raíz–suelo. El tamaño, la forma y la continuidad de los poros son, en gran medida, los factores determinantes del movimiento del aire y del agua. Las características de los poros están determinadas por la estructura edáfica de los agregados y ésta también interviene en el grado de interconexión de los poros lo cual influye en la infiltración y la permeabilidad de los suelos.
La velocidad de infiltración es considerada como un indicador clave de la calidad del suelo (Weixelmann et al., 1996; Arshad y Martin, 2002). Buol et al. (1981) y Porta et al. (1994) reconocen algunas subdivisiones del proceso de infiltración en el suelo. Ellos destacan el concepto de velocidad de infiltración instantánea como el volumen de agua que se infiltra a través de una superficie de suelo horizontal unitario, en un periodo de tiempo infinitamente pequeño. Asimismo, identifican a la velocidad de infiltración básica como la tasa constante registrada en un suelo después de un periodo de prueba que comprende ensayos de 3 a 4 h de duración (Buol et al., 1981; Porta et al., 1994).
El objetivo del presente trabajo fue conocer las velocidades de infiltración media (VIM) que adicionalmente comprende la percolación vertical, horizontal y radial del agua en el suelo de seis diferentes ecosistemas de la Reserva de la Biosfera Sierra Gorda: 1) bosque de pino (con dos tratamientos en el mantillo del suelo); 2) bosque de encino; 3) selva baja caducifolia; 4) pastizal, 5) matorral xerófilo de Larrea tridentata; y 6) matorral xerófilo de Acacia constricta. En el bosque de pino la VIM se observó en sitios con mantillo y en sitios con la capa de mantillo removida. La VIM se define como la cantidad acumulada de agua percolada por unidad de superficie, a lo largo del tiempo que ha durado el proceso (Porta et al., 1994). Se espera encontrar una mayor VIM en los suelos de los ecosistemas con abundante mantillo y vegetación, por ser condiciones favorables para este proceso. Del mismo modo, se espera que en los ecosistemas con suelos desnudos (como en el matorral xerófilo) se registren los menores valores de VIM.
MATERIALES Y MÉTODOS
La Reserva de la Biosfera Sierra Gorda está ubicada en el noreste del estado de Querétaro (entre 20° 50' y 21° 45' N, y entre 98° 50' y 100° 10' O; comprende una extensión de 383 567.45 ha). Forma parte de la Sierra Madre Oriental y se encuentra ubicada entre el Plano Costero del Golfo y el Altiplano Mexicano; otros estados que también comprenden la distribución de esta sierra son San Luis Potosí, Guanajuato e Hidalgo. En la parte central el clima que predomina es el semicálido–subhúmedo, al suroeste el clima es seco y semiseco semicálido y al noroeste y oeste los climas son templados subhúmedos; en todos los casos el régimen de lluvias es de verano y presentan una precipitación media anual entre 313.3 y 883.33 mm y una temperatura media anual entre 13.1 y 24.3 °C (INE, 1999).
Los sitios de matorral xerófilo se localizaron en la autopista México–Querétaro, desviación San Juan del Río–Tequisquiapan, en una localidad entre Vizarrón y Peña Blanca, km 88 (20° 56' N, 99° 44' O; a una altitud de 1600 m). En estos sitios el clima es seco con una temperatura media de 26 °C y suelos sin mantillo y con drenaje deficiente (Rzedowski, 1988). En esta localidad los suelos son de origen sedimentario con predominancia de grandes afloramientos de rocas calizas y presencia de lutitas abigarradas (INE, 1999); son suelos desnudos con muy poca profundidad y sin presencia evidente de materia orgánica (mantillo). El manejo cultural al que han estado sometidos estos suelos es en principio la extracción de rocas calizas y pizarras en lajas para la construcción. El primer sitio se localizó en la zona de lomeríos con dominancia de Larrea tridentata; la pendiente aproximada de este lugar es de 12.5°. El segundo sitio se ubicó a 500 m del cauce de un río temporal donde el estrato arbóreo alcanzó los 4 m de altura con la predominancia de Acacia constricta; en este sitio la pendiente del terreno alcanzó 10°. En ambos sitios la textura del suelo se caracterizó por ser francolimosa.
El sitio de bosque de pino se ubicó en el municipio de Pinal de Amoles (21° 8' N, 99° 37.5' O y una altitud de 2200 m). El suelo de esta localidad es de origen volcánico y su clasificación corresponde a la de un Andosol maduro y con buena profundidad (INE, 1999). El horizonte O se caracterizó por presentar una capa de acículas de pino que alcanzó 5 cm de profundidad y estas estructuras se observaron sin ningún rastro de fragmentación evidente. El elemento florístico predominante en esta localidad fue Pinus patula (Rzedowski, 1988). El terreno de esta localidad presentó una pendiente pronunciada que alcanzó los 12°. El manejo cultural al que antes se sometió el suelo de esta localidad fue el aprovechamiento forestal; 30 años atrás, en este sitio se sembraron los árboles de P. patula que en la actualidad alcanzan una altura de 5.5 m y cubren el suelo con sus acículas.
El sitio de selva baja caducifolia se ubicó en la carretera que une Jalpan de Serra con Arroyo Seco (21° 15' N, 99° 40' O, a una altitud de 600 m). El clima se cataloga como cálido y, en la actualidad, este sitio está dominado por especies arbóreas de hoja caduca. El suelo es de origen sedimentario con presencia de algunos elementos rocosos metamorfoseados (INE, 1999), la textura del suelo que predomina es la limosa con arenas finas y la capa de mantillo tiene un espesor de 4 cm. La pendiente del terreno de esta localidad fue de 13.5°. Los elementos florísticos de esta localidad están representados por Bursera sp., Acacia sp., Mimosa sp. y Prosopis sp., entre algunas otras (Rzedowski, 1988). El manejo cultural al que suele someterse este tipo de suelos es el desmonte a través de la técnica de roza–tumba–quema para abrirlos a la agricultura por períodos de 3.5 años. En años posteriores a la alta productividad que de ellos se extrae, son terrenos usados como potreros para la crianza del ganado. En este sitio, ya no se realizan estas dos prácticas.
El sitio de pastizal se localizó en el km 203 de la carretera Jalpan–Xilitla (21° 10' N, 99° 24' O, a una altitud de 950 m), en una zona de potreros asentados en suelos de origen sedimentario con nula presencia de mantillo (INE, 1999). El pastizal se ubicó sobre una planicie (de 5 ha), sin importantes variaciones de pendiente en la elevación del terreno. En un extremo de la localidad existe un abrevadero artificial para el ganado. El sitio ostenta un suelo compacto y de textura francoarcillosa. La vegetación actual está compuesta por pastos que alcanzan una altura de 5 cm. Según Rzedowski (1988), la vegetación original fue una selva baja caducifolia asentada en una zona de clima semiárido; de hecho, la selva baja es un bioma que aún se puede observar bien representado circundando este pastizal (a 3.5 km).
El sitio de bosque de encino se localizó en el km 228 de la carretera Jalpan–Xilitla (21° 17' N, 99° 13' O, a una altitud de 1300 m). Es un bosque abierto donde predomina Quercus spp. con algunos elementos de Juniperus sp. y de Brahea moorei (Rzedowski, 1988). El suelo de esta localidad fue de origen volcánico con elementos depositados por acarreos aluviales y eólicos (INE, 1999), se asienta sobre un sustrato basáltico con suelo profundo, con textura franca–arenosa y con una capa de mantillo de 4 cm de profundidad. El valor de la pendiente en el terreno de esta localidad fue de 11.5°. El principal manejo cultural practicado en esta localidad es forestal (extracción de tierra de encino para macetas).
En abril de 2005 (época de sequía para todas las localidades de la Sierra Gorda y del centro de México) se seleccionaron en cada ecosistema de 19 a 24 puntos al azar dentro de un área de 0.25 ha por cada localidad. En ellos se colocó a 10 cm de profundidad en el suelo un tubo de PVC hidráulico de 15 cm de altura y 10.16 cm de diámetro interno. Se vertieron 100 mL de agua dentro del tubo (cantidad equivalente a una carga hidráulica de 13 mm) y se cronometró el tiempo que tardó en percolarse. En el bosque de pino se tomó el tiempo de infiltración en suelo con mantillo y en suelo donde éste se retiró. En el matorral xerófilo se tomó el tiempo de percolación en dos zonas, una dominada por Larrea tridentata y, otra, dominada por Acacia constricta, cercana a un lecho donde se origina un río de temporal. Aunque en la literatura se sugiere que para el cálculo de la velocidad de infiltración, la aplicación de la ecuación de Philip es robusta (Philip, 1957; 1969), este trabajo tomó en consideración para el cálculo de la VIM la fórmula modificada de Porta et al. (1994):
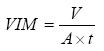
donde: V = volumen de agua, A = área interna del tubo y t = tiempo de infiltración.
Lo anterior, a razón de que se ajusta mejor al objetivo de este trabajo, dado que éste le asigna especial énfasis a la medición de la percolación del agua en intervalos de tiempo muy cortos y, a diferencia de uno de los supuestos paramétricos de la ecuación de Philip (Osuna y Padilla, 1998), este trabajo no tiene aplicación con fines productivos o utilitarios.
Análisis Estadístico
Se aplicó un análisis de varianza de una vía para determinar el efecto del tipo de ecosistema (incluyendo presencia/ausencia de mantillo en el bosque de pino) sobre la VIM. Se realizó una prueba de comparación múltiple de promedios de Tukey (Zar, 1999) para determinar las diferencias entre ecosistemas.
Para corroborar las diferencias en las tasas de infiltración de las zonas de matorral xerófilo, se aplicó una prueba de t de Student para muestras independientes (Zar, 1999).
RESULTADOS Y DISCUSIÓN
Se encontró un efecto significativo del tipo de ecosistema sobre la VIM (F6,144= 17.06, P < 0.001). La mayor velocidad de infiltración media (VIM) fue de 31.30 ± error estándar (e.e.) 4.02 mL cm–2 min–1 (intervalo 0.15–1.11 mL cm–2 min–1) y se registró en el bosque de pino con mantillo y difirió en forma significativa de las tasas obtenidas en la selva baja (7.74 ± 1.70 mL cm–2 min–1; 1.86–40.17 mL cm–2 min–1) y en el matorral xerófilo (con Larrea, 6.01 ± 0.80 mL cm–2 min–1, 0.68–13.34 mL cm–2 min–1; con Acacia,2.61 ± 0.73 mL cm–2 min–1, 0.72–16.62 mL cm–2 min–1) (Figura 1). Asimismo, la VIM del suelo del bosque de pino sin mantillo (6.01 ± 0.80 mL cm–2 min–1, 0.72–16.62 mL cm–2 min–1) fue significativamente más baja que la del suelo del mismo ecosistema con mantillo. La menor VIM se registró en el matorral xerófilo con Acacia y fue más baja que en bosque de pino con mantillo, bosque de encino (21.02 ± 3.20 mL cm–2 min–1, 6.18–66.60 mL cm2 min–1), pastizal (20.55 ± 2.25 mL cm–2 min–1, 6.00–40.80 mL cm–2 min–1) y selva baja (7.74 ± 1.70 mL cm–2 min–1, 1.86–40.17 mL cm–2 min–1).
Por otro lado, las VIM en bosque de encino y selva baja fueron más altas que en el bosque de pino sin mantillo (Figura 1).

Aunque la prueba de Tukey sugiere que no hay diferencias significativas de la VIM entre los dos sitios de matorral xerófilo, la prueba de t de Student para muestras independientes indica que esta variable es más alta en el sitio dominado por Larrea (t = 3.01, g.l. = 40, P = 0.0045).
Los resultados del presente trabajo muestran que la presencia de una capa de mantillo favorece la infiltración del agua, incrementando el volumen que es captado por el suelo. De manera clara, la VIM en el bosque de pino con el mantillo intacto fue cinco veces más alta que en los sitios del mismo ecosistema a los que se les removió esta capa protectora (Figura 1). Asimismo, los sitios asentados en bosques, con una capa de mantillo > 4 cm, tuvieron mayor VIM que las zonas de matorrales, cuyos suelos carecían de éste. Los resultados apoyan la hipótesis de Tamhane et al. (1978) en la que sostienen que la materia orgánica (MO) del suelo juega un papel central para facilitar el ingreso del agua, ya que funciona como una esponja capaz de absorber el agua de la lluvia reduciendo la escorrentía superficial. Por su parte, Lado et al. (2004) encontraron que los suelos de pastizales que tienen escasa MO tienen bajas tasas de infiltración. Estos autores discuten que si hay bajo contenido de MO, la estabilidad de los agregados disminuye y se incrementa la dispersión de las fracciones texturales, lo cual también favorece la disminución de la porosidad, disminuyendo la infiltración. Kunst et al. (2003) encontraron que los factores más importantes que afectan la velocidad de infiltración media son la presencia y el tipo de mantillo vegetal. Maass et al. (1983) observaron que en sitios donde se ha removido la vegetación y se introduce maíz existe una gran pérdida de nutrimentos por lixiviación; sin embargo, si se aplica una capa de mantillo encima del suelo en este tipo de sitios, la pérdida de nutrimentos por lixiviación se reduce entre un 80 y un 96%, lo cual sugiere una reducción en la escorrentía. Según Kunst et al. (2003), la velocidad de infiltración registrada bajo mantillo de plantas leñosas (con hojarasca y ramillas) superó casi en un 50% a la registrada en suelo desnudo y suelo con mantillo de gramíneas, lo cual explica también las mayores tasas de infiltración en los bosques, comparadas con las de los ecosistemas de suelo desnudo (matorrales semidesérticos), o bien, pastizales dominados por gramíneas.
En el presente estudio el pastizal, con un mantillo pobre y con compactación evidente, no siguió el patrón discutido en el párrafo anterior, ya que presentó una VIM de 20.55 ± 2.25 mL cm–2 min–1 (Figura 1), que es un valor muy similar al registrado en el suelo del bosque de encino, en el que se observó la presencia de una textura franca–arenosa, con una capa de mantillo de alrededor de 4 cm de profundidad. Los suelos de los pastizales están sometidos al efecto negativo del pastoreo del ganado sobre la VIM, ya que los suelos de estos ecosistemas se compactan por pisoteo del ganado y esto provoca que la distancia entre los agregados del suelo se reduzca lo que, a su vez, provoca que la aireación disminuya y se incremente la susceptibilidad a la erosión (Flamand–Rodríguez, 1995; Trimble y Mendel, 1995; Drewry et al., 2001). Sin embargo, el suelo del pastizal estudiado, aunque carecía de mantillo, registró altos valores de VIM. Se sugiere determinar el contenido de MO, la compactación y la textura de este suelo para explicar este fenómeno. La cercanía de este sitio a un abrevadero sugiere que el pastizal mantiene un alto contenido de MO debido a la deposición de heces del ganado caballar, lo cual favorecería la absorción de agua. Existen evidencias de que la velocidad de infiltración se incrementa de forma proporcional con la edad de un ecosistema boscoso (Sarukhán y Maass, 1990), mientras que en los pastizales sucede lo contrario (León–Arteta, 1991; Deuchars, 1999). En pastizales de Estados Unidos se ha registrado una tasa de infiltración de 1.2 × 10–5 mL cm–2 min–1 (Lado et al., 2004), la cual es más baja que la registrada en este atípico pastizal queretano.
En el matorral xerófilo, la zona que registró la mayor VIM fue la que estaba dominada por L. tridentata (Figura 1). Los resultados del presente trabajo sugieren que la diferencia de velocidades de infiltración entre zonas de matorral xerófilo se debe a la lejanía del sitio dominado por L. tridentata al río temporal, lo cual supone que este sitio es más seco que el dominado por A. constricta. Los suelos secos absorben mayor cantidad de agua que los que ya están humedecidos y se encuentran alrededor de la capacidad de campo (Tamhane et al., 1978; Porta et al., 1994). Otro factor que pudo influir en este resultado, es la diferencia de topografía que se observó entre sitios. El sitio con presencia de L. tridentata se ubicó en los dos tercios superiores del lomerío, mientras que el sitio dominado por A. constricta se ubicó en la cañada del río de temporal. Los suelos localizados en sitios con pendientes pronunciadas tienen una alta permeabilidad, ya que su textura es más arenosa, dado que las partículas finas se depositan en el fondo de las cañadas, lo cual es un factor que reduce la permeabilidad de los suelos en estos sitios (Tamhane et al., 1978; Daubenmire, 1979; Cobertera, 1993). Algunos datos de Sarukhán y Maass (1990) indican que los bosques tropicales son los ecosistemas de mayor edad y, por esta razón, han acumulado una mayor cantidad de MO en el suelo a lo largo del tiempo. Aunque en el presente estudio no se logró evaluar con precisión el efecto de la edad de los ecosistemas sobre la VIM, los datos señalan al bosque de encino como el ecosistema con el segundo valor más importante de VIM (Figura 1). En México este tipo de bioma representa una zona de transición muy importante entre los bosques de pino y los bosques mesófilos (bosques de niebla) (Rzedowski, 1988). Ambos son considerados como refugios de flora pleistocénica y dado que el dosel de los árboles del género Quercus spp. aportan una gran cantidad de MO al suelo, se les califica como especies clave en la restauración de ecosistemas cuyo suelo ha experimentado algún tipo de disturbio antropogénico. Por su parte, el bosque de pino con su capa de mantillo íntegra tiene una edad de 30 años, se trata de un ecosistema artificial en el que se introdujo Pinus patula a fin de lograr la rehabilitación de la vegetación en Pinal de Amoles. Este sitio de edad relativamente joven registró la mayor VIM debido a que las especies que dominan el estrato arbóreo del sitio (Pinus patula, 98%; Cupressus sp., 2%) se caracterizan por ser especies que pierden sus acículas y que se acumulan de manera importante en el suelo de estos ambientes (Torres–Espinosa et al., 2006).
Los resultados del presente trabajo indican que la VIM es una característica que varía dependiendo de las características mecánicas del suelo (compactación) en cada ecosistema; sin embargo, muchas suposiciones están basadas en la profundidad del mantillo observada en las localidades, por lo que se sugiere la implementación de un estudio cuantitativo que brinde más datos sobre el papel que juega la profundidad del mantillo sobre la tasa de infiltración. De igual forma, es necesario llevar a cabo estudios detallados sobre las características del suelo que afectan a la VIM, como son: la estructura de los agregados, la textura, la densidad aparente y real, la porosidad y el porcentaje de MO.
CONCLUSIONES
Los resultados sugieren que los factores que determinan la velocidad de infiltración media (VIM) son la presencia y la profundidad del mantillo. Los ecosistemas que tuvieron altos valores de VIM fueron el bosque de pino, el bosque de encino y el pastizal, y el que tuvo el menor valor de este parámetro fue el matorral xerófilo. Otros factores que son determinantes del valor de la VIM son: el tipo de roca que originó al suelo de cada ecosistema, la textura edáfica, la cercanía a los cuerpos de agua y la edad del ecosistema.
AGRADECIMIENTOS
Al grupo 5175 de Ecología I (semestre 2005–2) por su ayuda en la toma de datos. A la Unidad de Enseñanza de Biología de la Facultad de Ciencias de la Universidad Nacional Autónoma de México por su apoyo. A los Bióls. Marco A. Romero Romero y Miguel Hernández Alva por su apoyo técnico. Finalmente, al editor técnico, así como a dos revisores anónimos cuyas observaciones ayudaron a mejorar sustancialmente la elaboración de la versión final de este artículo.
LITERATURA CITADA
Arshad, M. and S. Martin. 2002. Identifying critical limits for soil quality indicators in agro–ecosystems. Agric. Ecosyst. Environ. 88: 153–160. [ Links ]
Buol, S. W., F. D. Hole y R. J. McCracken. 1981. Génesis y clasificación de los suelos. Trillas. México, D. F. [ Links ]
Cerdá, A. 1996. Seasonal variability of infiltration rates under contrasting slope conditions in Southeast Spain. Geoderma 69: 217–232. [ Links ]
Cobertera, E. 1993. Edafología aplicada. Cátedra. Madrid, España. [ Links ]
Daubenmire, R. F. 1979. Ecología vegetal, tratado de autoecología de plantas. 3a ed. Limusa. México, D. F. [ Links ]
Deuchars, S. 1999. Changes in soil structure and hydraulic properties in regenerating rain forest. Soil Use Manage. 15:183–187. [ Links ]
Drewry, J. J., K. C. Cameron, and G. D. Buchan. 2001. Effect of simulated dairy cow treading on soil physical properties and ryegrass pasture yield. N. Z. J. Agric. Res. 44: 181–190. [ Links ]
Flamand–Rodríguez, C. L. 1995. Introducción a la mecánica de suelos. 2a ed. Universidad Autónoma Chapingo. Chapingo, México. [ Links ]
Franzluebbers, A. J. 2001. Water infiltration and soil structure relates to organic matter and its stratification with depth. Soil Tillage Res. 66: 197–205. [ Links ]
Gregory, P. J., J. V. Lake, and D. A. Rose. 1987. Root development and function: effects of the physical environment. Cambridge University Press. Cambridge, UK. [ Links ]
Gurovich, L. 1985. Fundamentos y diseño de sistema de riego. Instituto Interamericano de Cooperación para la Agricultura (IICA). San José, Costa Rica. [ Links ]
INE (Instituto Nacional de Ecología). 1999. Programa de manejo de la Reserva de la Biosfera Sierra Gorda, México. INE–Secretaría de Medio Ambiente, Recursos Naturales y Pesca. México, D. F. [ Links ]
Kunst, C., R. Ledesma, N. Basan, G. Angela, D. Prieto y J. Godoy. 2003. Rolado de "fachinales" e infiltración de agua en suelo en El Chaco Occidental (Argentina). Revista de Investigaciones Agropecuarias 32: 105–126. [ Links ]
Lado, M., A. Paz, and M. Ben–Hur 2004. Organic matter and aggregate size interactions in infiltration, seal formation and soil loss. Soil Sci. Soc. Am. J. 68: 935–942. [ Links ]
León–Arteta, R. 1991. Nueva Edafología, regiones tropicales y áreas templadas de México. 2a ed. Distribuciones Fontamara. México, D. F. [ Links ]
Maass, J. M., C. F. Jordan, and J. Sarukhan. 1983. Soil erosion and nutrient losses in seasonal tropical agroecosystems under various management techniques. J. Appl. Ecol. 25: 343–363. [ Links ]
Návar, J. and T. J. Synnott. 2000. Soil infiltration and land use in Linares, N. L., Mexico. Terra 18: 255–262. [ Links ]
Osuna C., E. S. y J. S. Padilla R. 1998. Estimación de la sorbilidad e infiltración usando datos de simulación de lluvia para tres tipos de suelo de la zona semiárida de México. Terra 16: 293–302. [ Links ]
Philip, J. R. 1957. The theory of infiltration: 1. The infiltration equation and its solution. Soil Sci. 83: 345–357. [ Links ]
Philip, J. R. 1969. The theory of infiltration. Adv. Hydrosci. 5: 215–296. [ Links ]
Porta C., J., M. López–Acevedo y C. Roquero. 1994. Edafología para la agricultura y el medio ambiente. Mundi–Prensa. Madrid, España. [ Links ]
Rzedowski, J. 1988. Vegetación de México. Limusa. México, D. F. [ Links ]
Sarukhán, J. y J. M. Maass. 1990. Bases ecológicas para un manejo sostenido de los ecosistemas: el sistema de cuencas hidrológicas. pp. 81–114. In: E. Leff (ed.). Medio ambiente y desarrollo en México. Vol. I. Grupo Porrúa. México, D. F. [ Links ]
Tamhane, R., D. Motiramani y Y. Bali. 1978. Suelos: su química y fertilidad en zonas tropicales. Diana. México, D. F. [ Links ]
Torres–Espinosa, L. M., J. A. Sánchez–Salas y J. Jiménez–Pérez. 2006. Análisis estructural de un ecosistema forestal de Pinus–Quercus en la Sierra Madre Oriental. Ciencia Forestal en México 31: 7–30. [ Links ]
Trimble, S. W. and A. C. Mendel. 1995. The cow as a geomorphic agent: a critical review. Geomorphology 13: 233–253. [ Links ]
Weixelman, D., D. Zamudio, K. Zamudio, and R. Tausch. 1996. Classifying ecological types and evaluating site degradation. J. Range Manage. 50: 315–321. [ Links ]
Zar, J. H. 1999. Biostatistical analysis. Prentice Hall. Upper Saddle River, NJ, USA. [ Links ]
* Publicado como nota de investigación en Terra Latinoamericana 26: 21–27.

