











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Una vez que ocurre la fecundación del óvulo por el espermatozoide, se desencadena una secuencia de eventos que continúan con la implantación, el desarrollo de la placenta y el embrión y que finalizan, idealmente, con el nacimiento alrededor de las 38-40 semanas de gestación; esta serie de acontecimientos se denominan embarazo1. Al término del embarazo ocurre el trabajo de parto, el cual está caracterizado, entre otros acontecimientos, por la aparición de contracciones uterinas regulares que aumentan su frecuencia (aumento de su ocurrencia en un intervalo de tiempo) e intensidad2. Cuando el nacimiento ocurre antes del periodo normal establecido, se considera un parto pretérmino, el cual se define clínicamente como aquel que comienza antes de las 37 semanas de gestación3. Los nacimientos prematuros siguen siendo la principal causa de muerte y discapacidad neonatal en el mundo4,5. Por otro lado, la disminución en la capacidad contráctil del miometrio (atonía uterina) está fuertemente asociada a la hemorragia posparto, una de las principales causas de mortalidad materna6. Por lo tanto, resulta muy importante conocer los mecanismos fisiológicos, celulares y moleculares involucrados en el establecimiento del trabajo de parto para determinar cuáles son los factores que podrían provocar las patologías asociadas al parto.
Uno de los tejidos más importantes para que se lleve a cabo el trabajo de parto es el miometrio, el cual es la capa muscular del útero. En comparación con otras células musculares, las células del miometrio (las cuales se conocen como miocitos uterinos) tienen un tamaño relativamente grande y pueden almacenar grandes cantidades de calcio (Ca2+), lo que les permite llevar a cabo contracciones de gran intensidad7. Al término del embarazo, los miocitos experimentan una serie de cambios fisiológicos, bioquímicos y moleculares, lo que permite que el tejido pase de un estado o fenotipo de quiescencia a un fenotipo contráctil que favorecerá el inicio del trabajo de parto8. En parte, esto es provocado por la acción de las hormonas sexuales progesterona (P4), estradiol (E2) y la hormona peptídica9-11. Estas hormonas ejercen su acción mediante sus receptores intracelulares y membranales específicos12-14.
Hasta el momento, no se conocen por completo los mecanismos moleculares implicados en la activación del miometrio durante el trabajo de parto. En particular, se tiene poca información acerca de cuál es el efecto de las hormonas sexuales en la actividad miometrial. En el presente trabajo se realizó una revisión bibliográfica de trabajos en los que se han estudiado los mecanismos moleculares involucrados en la regulación de la actividad de las células miometriales por las hormonas P4, E2 y OXT, y sus receptores correspondientes.
Eventos y moléculas que desencadenan las fases del trabajo de parto
El trabajo de parto es un proceso fisiológico único que ocurre durante el término de la gestación, el cual requiere la rotación del feto hacia la posición de nacimiento, lo que va acompañado de una cascada de cambios endocrinos que termina con un parto exitoso15. El miometrio se encuentra relativamente quiescente durante el 95% del embarazo, lo cual corresponde a la fase 0 del parto. La activación del miometrio corresponde a la fase 1 y se efectúa predominantemente por el estiramiento mecánico del útero, y por medio de la acción de los estrógenos (principalmente el E2). La estimulación corresponde a la fase 2, cuando las uterotoninas endógenas, incluidas las prostaglandinas (PG) y la OXT, actúan sobre el miometrio activado. Esta serie de eventos concluye con la involución posparto, que corresponde a la fase 316.
A continuación se describen las fases del trabajo de parto que abarcan desde el embarazo hasta el término de este, junto con las moléculas involucradas en cada etapa.
Embarazo. Fase 0 del parto
Durante el desarrollo del embarazo, el útero se mantiene en un estado de reposo o quiescencia principalmente por la acción de la P4 y otros factores como la prostaciclina, la relaxina, el péptido relacionado con la hormona paratiroidea, el péptido relacionado con el gen de la calcitonina, el péptido intestinal vasoactivo y el óxido nítrico17. Todos estos agentes actúan en los miocitos uterinos como mediadores de un aumento de las concentraciones intracelulares de monofosfato de adenosina cíclico (AMPc) o monofosfato de guanosina cíclico, los cuales inhiben la liberación de Ca2+ intracelular para evitar así la contracción del miometrio11,17.
Activación del miometrio. Fase 1 del parto
La activación del miometrio está asociada a un aumento de E2 y de la hormona liberadora de corticotropina, junto con el estiramiento mecánico de este tejido que es provocado por el crecimiento fetal18. Dichos eventos conducen a una regulación positiva de la expresión de un grupo de genes necesarios para las contracciones, los cuales codifican a las proteínas asociadas a la contracción (PAC). Dentro de las PAC, se incluye a GJA1, también conocida como conexina 43, la cual forma uniones comunicantes o tipo gap entre los miocitos uterinos, permitiendo la comunicación entre cada una de las células que forman este tejido para llevar a cabo contracciones coordinadas19. Otras PAC corresponden al receptor de oxitocina (OXTR) y a la ciclooxigenasa 2 (PTGS2), que estimulan el inicio del trabajo de parto2,17.
Estimulación del miometrio. Fase 2 del parto
La fase de activación prepara al miometrio para responder a los estimulantes producidos que provocan la contractilidad miometrial. La contracción es estimulada por uterotoninas como la OXT y citocinas proinflamatorias como la interleucina (IL) 1β, la IL-6, la IL-8, el factor de necrosis tumoral alfa y la proteína quimiotáctica de monocitos (MCP-1) las cuales promueven el trabajo de parto11,20.
Involución del útero. Fase 3 del parto
La tercera etapa del trabajo de parto se define como el tiempo que transcurre desde el nacimiento del bebé, la expulsión de la placenta y la reparación del tejido que forma al útero16. Esta etapa incluye la involución uterina y se ha atribuido principalmente a los efectos de la OXT21. Los cambios moderados en las concentraciones de OXT en el plasma materno al final de la gestación se compensan con un aumento sustancial en la expresión del OXTR en el miometrio, lo que causa un aumento dramático en la sensibilidad uterina a la estimulación con esta hormona2,11,17.
Características del miometrio
El miometrio es la pared más gruesa del útero, la cual está conformada por dos tipos de capas musculares: circular y longitudinal22. Durante el embarazo, el miometrio sufre hipertrofia (incremento de tamaño) e hiperplasia (aumento del número de células). Durante el trabajo de parto, el miometrio experimenta contracciones rítmicas que ayudan al feto a atravesar el cuello uterino que se ablanda y dilata al término del embarazo8.
En un estudio reciente, se demostró mediante la técnica de secuenciación masiva de transcriptoma de célula única (single-cell RNA-seq), que el miometrio humano está compuesto principalmente por tres tipos de células de músculo liso, monocitos/macrófagos, células estromales, células endoteliales, entre otros tipos celulares, los cuales se comunican y participan en procesos asociados al parto. En particular, se identificaron tres tipos de células de músculo liso que se denominaron SMC-1, SMC-2, SMC-3 (por su nombre en inglés smooth muscle cells). Durante el trabajo de parto, el perfil transcripcional de estas células se asoció a los siguientes procesos biológicos: SMC-1 a las contracciones musculares, SMC-2 a la expresión de IL-1β e IL-6 (moléculas proinflamatorias indispensables durante el trabajo de parto) y SMC-3 a la respuesta al estímulo inflamatorio del interferón gamma. Con este estudio se confirmó la naturaleza contráctil de las SMC uterinas y se amplió el conocimiento acerca de la participación de estas células en el proceso inflamatorio que tiene lugar en el parto23.
Las células del músculo liso del miometrio están caracterizadas por tener forma de huso (forma alargada y elipsoide), núcleo central, un elevado número de mitocondrias, retículo sarcoplásmico (RS) y mioproteínas, como la actina y miosina24. Lo que distingue a los miocitos uterinos de otras células musculares es su tamaño relativamente grande, de hasta 0.5 mm de largo, que permite la entrada de grandes cantidades de Ca2+ y llevar a cabo poderosas contracciones7. La membrana plasmática delimita el miocito miometrial y alberga los canales iónicos, bombas e intercambiadores necesarios para la función uterina7.
Una característica universal de la contracción en las células musculares es el aumento en la concentración de Ca2+ intracelular. En el músculo liso, el Ca2+ tiene una función dual que consiste en activar los potenciales de acción y formar el complejo Ca2+-calmodulina. Este complejo activa a la cinasa de cadena ligera de miosina, que a su vez fosforila a la cadena ligera de la miosina (MLC) para que la miosina interactúe con la actina y finalmente se lleve a cabo la contracción7.
Los miocitos uterinos se encuentran conectados por uniones comunicantes o tipo gap. Estas conexiones están formadas por la conexina 43, la cual es una proteína transmembranal que oligomeriza formando un canal de seis subunidades que se conectan con el canal correspondiente a la célula adyacente, permitiendo de esta manera el acoplamiento eléctrico entre las células25,26. Se ha demostrado que las uniones entre los miocitos uterinos y su correspondiente sincronización eléctrica juegan un papel fundamental en el desarrollo gradual de la contractilidad uterina durante el trabajo de parto27-29. Durante el parto, el miometrio uterino pasa de ser un tejido con una conectividad relativamente baja entre los miocitos individuales a un tejido con amplias conexiones físicas mediadas por los canales de conexina 4330. Las conexiones entre los miocitos durante el parto también se forman por la liberación paracrina de prostaglandina F2α y la liberación local de Ca2+. Esta amplia conectividad física y bioquímica permite que la despolarización en los miocitos individuales se transmita a las células vecinas y así se forman extensas ondas de despolarización y contracción en grandes áreas del útero. Esto provoca un aumento de la presión intrauterina y una distensión progresiva del cuello uterino, lo que conduce a la expulsión del feto8.
Participación de los estrógenos, progesterona y oxitocina en el trabajo de parto
La función de las células miometriales durante el embarazo está controlada por una interacción compleja de señales endocrinas, paracrinas y autocrinas de los sistemas materno y fetal que inducen dos estados principales: 1) inactividad para el mantenimiento del embarazo, y 2) trabajo de parto, que termina el embarazo10.
En los mamíferos, esta transición es provocada por la alteración de los niveles y/o respuesta a las hormonas esteroides sexuales E2 y P4 en torno al término de la gestación. En la mayoría de los mamíferos, los niveles de P4 disminuyen y los niveles E2 aumentan o se mantienen constantes, dependiendo la especie31. En el humano, los niveles de E2 y P4 se mantienen elevados al término del embarazo, por lo que su participación en esta etapa del embarazo depende de la respuesta de las células miometriales a dichas hormonas31 (Fig. 1). Durante el término del embarazo, también se presenta un aumento en los niveles de OXT durante el parto (Fig. 1), que si bien no es una hormona esteroide, es importante para que este proceso y la involución del útero se lleven a cabo de una manera adecuada32. Para una mejor comprensión de los mecanismos de acción de las hormonas E2, P4 y OXT durante el trabajo de parto, nos centraremos en los efectos que desencadenan estas hormonas al unirse a sus receptores específicos de manera general y específica para cada hormona.
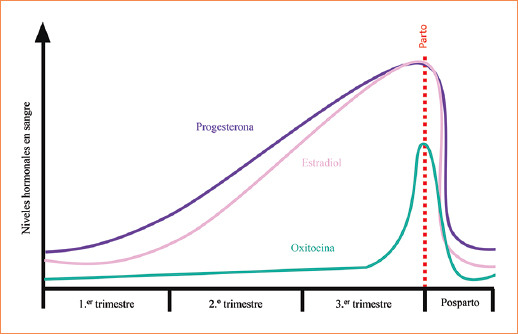
Figura 1 Niveles relativos de las hormonas progesterona, estradiol y oxitocina en sangre a lo largo del desarrollo del embarazo. Las escalas de los niveles hormonales en sangre son aproximadas.
Receptores de hormonas implicadas en el trabajo de parto
La acción de las hormonas en el miometrio ocurre por medio de sus receptores hormonales, de los cuales hay dos tipos: los receptores intracelulares y los receptores membranales (Fig. 2).
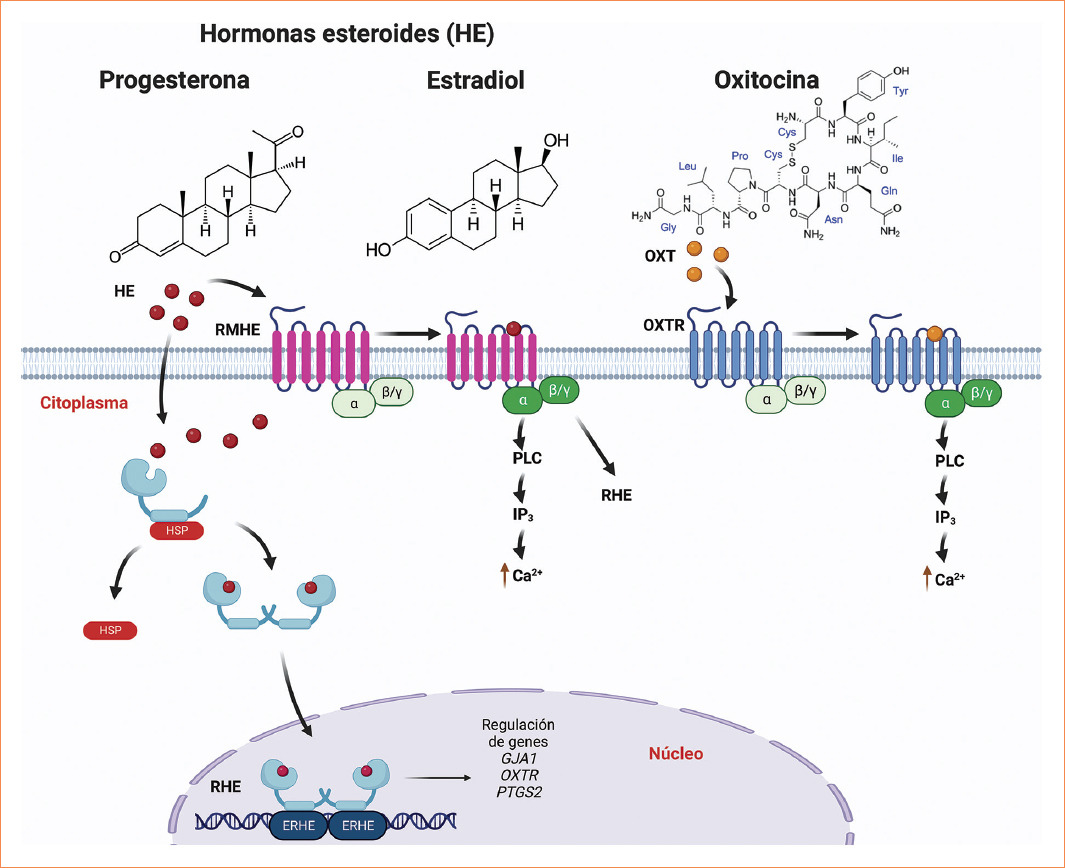
Figura 2 Mecanismo de acción de las hormonas esteroides (E2 y P4) y la OXT en el miometrio humano.E2: estradiol; ERHE: elementos de respuesta a hormonas esteroides; HSP: complejo de chaperonas multiproteicas; IP3: inositol trifosfato; OXT: oxitocina; OXTR: receptor membranal de OXT; P4: progesterona; PLC: fosfolipasa C; RMHE: receptor membranal de hormonas esteroides.
Los receptores intracelulares generalmente actúan como factores de transcripción, que son proteínas que interaccionan con sus genes blanco para inducir o inhibir su expresión. Estos receptores cuentan con un dominio de unión específico para una hormona (generalmente de tipo esteroide), un dominio de unión a ADN y varios dominios de unión a otras proteínas que regulan su actividad transcripcional33. En la mayoría de los casos, los receptores intracelulares se localizan en el citoplasma, unidos a chaperonas (generalmente proteínas de choque térmico) que inhiben su activación. Una vez que las hormonas entran a la célula por medio de difusión simple a través de la membrana plasmática, se forma el complejo hormona-receptor, las chaperonas se separan del receptor y este se asocia en forma de dímeros que son objeto de una serie de modificaciones postraduccionales para finalizar su activación. El complejo hormona-receptor se transloca al núcleo y se une a dominios de unión al ADN conocidos como elementos de respuesta a hormonas, que son secuencias específicas localizadas en la región promotora de sus genes blanco a la cual se reclutan cofactores para regular su expresión34.
Los receptores membranales de hormonas generalmente se encuentran acoplados a proteínas G, que al activarse conducen a la formación de moléculas que actúan como segundos mensajeros, que a su vez inducen la activación de una cascada de señalización en la célula35.
Oxitocina y su papel en la regulación del inicio del trabajo de parto
Durante el parto, los niveles de OXT incrementan de manera local en los tejidos placentarios y uterinos36. Se ha reportado que los niveles plasmáticos de OXT se mantienen relativamente constantes durante el inicio del trabajo de parto, sin embargo, los niveles de esta hormona incrementan durante el parto y disminuyen de manera gradual durante el posparto37,38. La OXT únicamente actúa mediante su receptor membranal (OXTR), cuya expresión en el útero aumenta hasta 200 veces al término del embarazo39. El aumento de la sensibilidad a la OXT favorece la aparición de las contracciones uterinas10.
El OXTR pertenece a la clase A de la superfamilia de receptores membranales acoplados a la proteína G. Cuando la OXT se une a su receptor, este sufre cambios conformacionales que inducen la disociación de las subunidades b y g de la proteína G, y por lo tanto, la activación de la subunidad a. Esto induce la fosforilación y activación de la fosfolipasa C, la cual cataliza la hidrólisis de fosfatidilinositol 4,5-bifosfato a inositol-trifosfato (IP3) y diacilglicerol40,41. El IP3 funciona como un segundo mensajero que propaga la señal de la OXT, activando la liberación de los iones Ca2+ del interior del RS hacia el citoplasma. El RS es un orgánulo vesicular presente en las células de músculo liso formado por una bicapa lipídica, que permite el paso unidireccional de los iones, permitiendo su almacenamiento hasta el estímulo de liberación provocado por el IP342.
Como se describió anteriormente, el Ca2+ liberado estimula las contracciones mediante su acción por medio de la calmodulina14. Además, hay una redistribución del Ca2+ mediante la formación de canales de comunicación intercelular por medio de la proteína GJA1, lo que favorece la propagación de las contracciones en todo el tejido miometrial.
Progesterona y su efecto durante el embarazo y el trabajo de parto
La P4 actúa principalmente mediante su receptor intracelular, codificado por el gen PGR. Este gen cuenta con dos sitios de inicio de la transcripción y da como producto a las isoformas PGR-A y PGR-B, las cuales son idénticas excepto en la región amino-terminal de la isoforma B, que es más larga por 164 aminoácidos43. Ambas isoformas funcionan como factores de transcripción al regular la expresión de sus genes blanco34.
En el caso del ser humano, no hay una disminución en los niveles plasmáticos de P4 durante el término del embarazo, sino que la inducción del parto es desencadenada por un cambio en la proporción de la expresión de las dos isoformas del PGR44. A lo largo del embarazo predomina la isoforma PGR-B, que inhibe la expresión de genes proinflamatorios y del receptor de estrógenos alfa (ESR1). Esto mantiene al útero en un estado de quiescencia durante el embarazo43. Al inicio del trabajo de parto, cambian las proporciones de PGR-A respecto a PGR-B, aumentando la expresión de la isoforma A, que induce la expresión de genes proinflamatorios como IL-6, IL-1B y PTGS245. Además, PGR-A promueve la expresión del gen GJA1 que codifica para la conexina 43, proteína esencial para la propagación de las contracciones durante el parto46.
Hasta el momento, aun no se conoce por completo cuál es el mecanismo asociado al cambio en la proporción de las isoformas del PGR durante el inicio del trabajo de parto, aunque se ha propuesto que mecanismos epigenéticos pudieran estar involucrados47. Adicionalmente, se ha propuesto que durante el inicio del trabajo de parto los niveles intracelulares de P4 en los miocitos uterinos disminuyen debido al aumento de la expresión de la 20-α hidroxiesteroide deshidrogenasa, la cual es una enzima que metaboliza a la P4, sugiriendo que la actividad de la isoforma PGR-A es independiente de ligando48.
Existe una amplia variedad de receptores membranales de P4 que se dividen en dos tipos principales. El primer tipo de estos receptores se compone de los isotipos de la familia de los receptores membranales de P4 y adiponectina Q (PAQR) acoplados a proteína G y los principales son PAQR5, PAQR7 y PAQR849. A lo largo del embarazo, la activación de los receptores PAQR5 y PAQR7 potencia la actividad de PGR-B para el mantenimiento de la quiescencia uterina durante el embarazo. Por otro lado, ambos receptores pueden inducir la fosforilación de la MLC para activar la contracción del miometrio durante el trabajo de parto50.
Adicionalmente, se encuentra la familia de los componentes de membrana del receptor de P4 (PGRMC), de los cuales se encuentran el PGRMC-1 y PGRMC-251. Se ha propuesto que estos receptores membranales participan en la falta de la respuesta a la P4 que se presenta en el miometrio durante el trabajo de parto52, sin embargo se requieren de más estudios para confirmarlo.
Estradiol y su participación en la inducción del trabajo de parto
El E2 es el estrógeno más potente en el cuerpo humano53. El E2 puede actuar mediante receptores intracelulares y membranales. Existen dos receptores intracelulares del receptor de estrógenos, codificados en dos genes distintos, ESR1 y ESR2. Ambos genes se expresan a lo largo del embarazo en las células del miometrio, pero ESR1 es el único que, durante el parto, cambia su expresión de forma significativa54.
Durante el trabajo de parto, se inhibe la represión de la expresión del gen ESR1 mediada por PGR-B, lo que aumenta la sensibilidad del miometrio a los estrógenos. A su vez, el ESR1 unido al E2 incrementa la transcripción del OXTR, que al activarse, libera iones de Ca2+, necesarios para las contracciones uterinas55.
El ESR1 cuenta con varias isoformas, entre ellas las de mayor interés en el miometrio durante el trabajo de parto son: ER66 que contiene todos los exones8 codificados en el gen y ERΔ7, que omite el séptimo exón. A lo largo del embarazo, ERΔ7 es la isoforma predominante y se encarga de inhibir la expresión de ER66. La expresión de la isoforma ERΔ7 es favorecida por el factor de splicing RBMX o proteína con motivo de unión a ARN asociada al cromosoma X. Cuando aumentan los niveles de E2 al término del embarazo, se inhibe la expresión de RBMX, evitando la síntesis de la isoforma ERΔ7 y por lo tanto, permitiendo la síntesis completa de ER66, que induce la expresión de GJA1 al término del embarazo56.
El receptor membranal de estrógenos (GPER1) regula la expresión de diversos genes de manera indirecta. El GPER1 actúa estimulando la producción de segundos mensajeros, como AMPc, que a su vez estimulan la activación de proteínas como la proteína cinasa C y la proteína cinasa A activada por AMPc57. En particular, se ha demostrado que la activación selectiva de GPER1 aumenta la respuesta contráctil a OXT en las células de miometrio58, por lo que contribuye a la propagación e inicio de las contracciones durante el parto.
Conclusiones y perspectivas
El trabajo de parto es un evento fisiológico que ha sido objeto de estudio desde hace más de un siglo, sin embargo aun no se conocen por completo los mecanismos celulares y moleculares implicados en su inicio. El miometrio es la capa muscular del útero que se encarga de llevar a cabo las contracciones durante el trabajo de parto. Se ha demostrado que existen múltiples factores que inducen la actividad contráctil del miometrio, tales como la inflamación, estímulos mecánicos, el estrés oxidante, el cortisol y las hormonas sexuales, entre otros. En este trabajo, nos centramos en el papel de las hormonas, E2, P4 y OXT en la actividad de las células y/o el tejido miometrial. En general, la P4 mantiene la quiescencia del miometrio durante el embarazo al regular la expresión de moléculas proinflamatorias y proteínas asociadas a la contracción, mientras que, al término del embarazo, el E2 induce la expresión de dichas moléculas. Por su parte, la OXT induce un aumento en la concentración de Ca2+ intracelular para llevar a cabo las contracciones de los miocitos uterinos.
Hasta el momento, no se han dilucidado por completo los mecanismos moleculares por medio de los cuales actúan estas hormonas en el miometrio humano, así como la relación que guardan entre sí al inicio del trabajo de parto. Las técnicas actuales de edición genómica serán de gran utilidad para establecer cuál es la participación de los receptores y moléculas señalizadoras que responden a estas hormonas59. Asimismo, el estudio detallado de otros procesos celulares y moleculares, como el estrés del retículo endoplásmico60 y mecanismos epigenéticos61 y su contribución a la activación del miometrio permitirá en un futuro identificar la compleja red de procesos que inducen el trabajo de parto, tanto fisiológico como patológico. En este sentido, el entendimiento de estos procesos será fundamental para la búsqueda de blancos terapéuticos y estrategias para la prevención de patologías muy frecuentes que ponen en riesgo la vida y calidad de vida de los neonatos y sus madres, tales como el parto pretérmino y la hemorragia posparto6.
Finalmente, tomando en cuenta que gracias a los estudios de célula única se conoce el atlas de las células que componen al miometrio23, los cultivos tridimensionales62 serán de gran utilidad para comprender los procesos que ocurren en este tejido y la participación de las hormonas sexuales de una manera más integral y cercana al tejido in vivo.

