










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkUniversidad y ciencia
versão impressa ISSN 0186-2979
Universidad y ciencia vol.29 no.1 Villahermosa Abr. 2013
Artículos
Condición corporal de la cerceta ala azul (Anas discors) obtenida por actividad cinegética en el Estado de México
Body condition of the blue-winged teal (Anas discors) hunted in the state of Mexico
Nany Stivalis Cárdenas-García1, Arturo Cortes-Cuevas2, José Antonio Quintana-López3, Gary García-Espinosa4
1 Dpto. Medicina y Zootecnia de Aves. Facultad de Medicina Veterinaria y Zootecnia. UNAM.
2 Centro de Enseñanza, Investigación y Extensión en Producción Avícola. Facultad de Medicina Veterinaria y Zootecnia.
3 Dpto. Producción Animal: Aves. Facultad de Medicina Veterinaria y Zootecnia. UNAM.
4 Dpto. Producción Animal: Aves. Facultad de Medicina Veterinaria y Zootecnia. UNAM. Av. Universidad 3000. UNAM,CU. Coyoacán, México. D.F. CP 04510. Correo electrónico: gary@unam.mx
Artículo recibido: 14 de abril de 2009
Aceptado: 14 de enero de 2013
RESUMEN
La condición corporal de la cerceta ala azul (Anas discors) fue evaluada por dos años consecutivos durante su estancia en la Laguna de Chiconahuapan, un humedal fragmentado de las Ciénegas de Lerma en el Estado de México. El estudio incluyó la evaluación del peso corporal, grasa subcutánea, y ancho y grosor de músculos pectorales en ejemplares de cerceta de ala azul. En el primer año de estudio se observó que el peso corporal y la grasa subcutánea decrecieron progresivamente de noviembre a febrero; en los músculos pectorales, decreció el ancho e incrementó el grosor. En el segundo año, el peso y la grasa decrecieron progresivamente de noviembre a enero; en cuanto a los músculos pectorales no se observaron variaciones significativas. El presente estudio mostró que la cerceta de ala azul, durante su estancia en el humedal fragmentado del Estado de México, mantiene una condición corporal similar a la de otras especies de aves acuáticas.
Palabras clave: Anas discors, peso corporal, músculos pectorales, grasa subcutánea, humedal fragmentado.
ABSTRACT
The body condition of the Blue-winged Teal (Anas discors) was evaluated for two consecutive years during the bird's stay at Chiconahuapan lagoon, a fragmented wetland in the Ciénegas de Lerma, state of Mexico. The study included the evaluation of body weight, subcutaneous fat, and width and depth of pectoral muscles in specimens of Blue-winged Teal. During the first year of the study, body weight and subcutaneous fat decreased progressively from November to February; the width of the pectoral muscles decreased while the depth increased. During the second year, body weight and subcutaneous fat decreased progressively from November to January; no significant differences were observed in the pectoral muscles. The study showed that the Blue-winged Teal maintains a body condition similar to that of other waterfowl species during its stay in the fragmented wetland of the state of Mexico.
Key words: Anas discors, body weight, pectoral muscle, subcutaneous fat, fragmented wetland.
INTRODUCCIÓN
Se han realizado varios estudios en anátidos migratorios, donde se ha demostrado que tienden a presentar variaciones invernales en su condición corporal (Bellrose & Hawkins 1947, Ankney 1982, Reinecke & Stone 1982, Whyte & Bolen 1984, Baldassarre et al. 1986, Miller 1986, Whyte et al. 1986, Thompson & Baldassarre 1990, Loesch et al. 1992). No se sabe con exactitud lo que da origen a dichas variaciones, sin embargo se menciona la participación de factores endógenos (Reinecke & Stone 1982, Baldassarre et al. 1986, Perry et al. 1986, Loesch et al. 1992) y factores exógenos (Joyner et al. 1984, Paulus 1984, Baldassarre et al. 1986).
En esos estudios no se reportan las condiciones ecológicas en que se encuentran los humedales donde se refugian las aves. Año con año, México alberga diversas especies de aves durante el invierno, incluyendo 22 especies de anátidos migratorios (Sibley 2001).
Estas aves arriban a diferentes humedales, uno de ellos, son las Ciénegas de Lerma que se localizan en el Estado de México y que se caracterizan por ser hábitats fragmentados por actividades agrícolas y asentamientos urbanos (CONABIO 2007a, 2007b), lo cual podría generar un impacto negativo sobre la condición corporal de las aves.
La mayoría de los estudios respecto al comportamiento de la condición corporal de anátidos migratorios se ha realizado en regiones neárticas y solo un estudio en la península de Yucatán, México (Thompson & Baldassarre 1990). Sin embargo, se desconoce cuál es el comportamiento de la condición corporal de anátidos migratorios que habitan en humedales fragmentados cercanos al geotrópico como son las Ciénegas de Lerma en el Estado de México (CONABIO 2007a, 2007b), por lo que, el objetivo del presente estudio fue evaluar la condición corporal en la cerceta ala azul (Anas discors) a través del peso corporal, distribución de la grasa subcutánea, ancho y grosor de músculos pectorales durante dos temporadas otoño-invierno. La cerceta ala azul fue elegida para el estudio, por ser una especie que anida en el sur de Canadá, norte y centro de Estados Unidos de América, y cuyo comportamiento migratorio la hace recorrer largas distancias hacia el sur del continente para alojarse en México, Centroamérica y la parte norte de Sudamérica durante la temporada otoño-invierno (Price et al. 1995, Howard & Moore 1998, Sibley 2001).
MATERIAL Y MÉTODOS
Área de estudio
El estudio se realizó en la Laguna de Chiconahuapan, que es una de las tres lagunas que conforman las Ciénegas de Lerma y es la que se encuentra en las mejores condiciones ecológicas. Está ubicada en la cuenca alta del Río Lerma en el Estado de México (Vázquez 2004, CND 2007). Dicha laguna comprende gran parte del municipio de Almoloya del Río y una pequeña parte del municipio de Texcalyacac, ambos pertenecientes al Estado de México. Se localiza en las coordenadas 19°06'29" de Latitud Norte y 19°30'12" de Longitud Oeste.
Tiene une cobertura de 596 Ha y una altitud de 2600 m.s.n.m. (CND 2007) (Figura 1). De acuerdo con la clasificación de Köpen, el clima es templado subhúmedo, semifrío y frío con lluvias en verano y parte del otoño. Temperatura media anual 10-14 °C. La precipitación pluvial anual es de 7001200 mm, con una evaporación media anual de 2439 mm, (CONABIO 2007a). El 75 % de la vegetación corresponde a productos agrícolas (maíz), del 25 % restante, el 70 % son tulares y el 30 % son claros y otras plantas acuáticas (Vázquez 2004). Las Ciénegas de Lerma, albergan 146 especies de aves de los siguientes órdenes: Podicipediformes, Ciconiformes, Gruiformes, Charadrüformes, Anseriformes, Falconiformes, Columbiformes, Piciformes, Apodici-formes y Passeriformes (Vázquez 2004, CONABIO 2007a, 2007b), donde la cerceta ala azul (Anas discors) es la especie de anátido más abundante con base a observaciones previas de campo en el lugar de estudio. Además de culebras, ajolotes, ranas, ratones, tlacuaches y peces (Vázquez 2004, CONABIO 2007a).
Permisos
El presente trabajo se realizó con el permiso de colecta científica de la SEMARNAT SGPA/DGVS/05688/06 y bajo el convenio de colaboración SEMARNATUNAM 17419146025X05.
Obtención de ejemplares
Las cercetas fueron producto de la actividad cinegética legal del lugar durante la temporada otoño-invierno (noviembre-febrero). Para el primer año, se realizaron 5 muéstreos, obteniéndose 152 ejemplares de cerceta de ala azul en total; para el segundo año se colectaron 92 ejemplares obtenidos durante 4 muéstreos. Lo anterior debido a la migración temprana de las aves y al consecuente cierre de la temporada cinegética (Tabla 1).

Peso corporal
Se pesaron los ejemplares en el borde de la laguna, con una báscula de reloj (PERALPAMR) con rango de medición de 25 g a 10 kg. Se analizaron los datos considerando 2 grupos diferentes de individuos: 1) machos y 2) hembras.
Músculos pectorales
La pérdida de tejido muscular en los músculos pectorales se caracteriza por una disminución en su grosor con base a la quilla, que es una evaluación cualitativa (Harrison & Ritchie 1994). En este estudio se realizó una medición cuantitativa del ancho y grosor de los músculos pectorales con un flexómetro de un metro de longitud y un vernier, respectivamente. El ancho de ambos músculos pectorales superficiales se midió desde la quilla hasta el borde externo del esternón. La medición del grosor del músculo consistió en hacer una incisión perpendicular en la porción craneal de ambos músculos pectorales sobre el eje longitudinal de la quilla con el bisturí a 3 mm. del extremo craneal de la quilla. El vernier se introdujo hasta alcanzar el hueso coracoides y se registró la lectura hasta dos decimales.
Grasa subcutánea
Para evaluar la distribución de la grasa subcutánea en todos los ejemplares se realizó la disección de la piel, retirándola completamente, excluyendo la piel de los tarsos-metatarsos, y la de los carpos, metacarpos y falanges (Gill 2001). Para la evaluación cualitativa de la deposición de la grasa en todos los ejemplares, se elaboraron esquemas individuales, ejemplificando la cantidad y distribución de la grasa sobre las diferentes regiones corporales del ave. Todas las observaciones y los esquemas fueron realizados por la misma persona. Con base a los esquemas, la grasa se clasificó como: escasa (+), regular (++). moderada (+++) y abundante (++++).
Análisis estadístico
Los datos obtenidos en la temporada otoño-invierno en los dos periodos evaluados (2006-2007 y 2007-2008), fueron sometidos a un análisis de varianza con arreglo factorial 2X2; donde un factor fueron los dos periodos antes señalados durante la temporada otoño-invierno y el otro factor fue el sexo (macho y hembra) (Steel & Torrie 1980). Este análisis se utilizó para comparar el efecto entre los dos periodos evaluados, así como el efecto del sexo sobre las variables: peso, ancho y grosor del músculo pectoral de los patos. Posteriormente, se hizo un análisis de correlación múltiple de Pearson de dos colas para saber cual fue la correlación que existió entre el sexo, peso, grosor y ancho de los músculos pectorales de los patos con respecto a los dos periodos (Steel & Torrie 1980). Además, en cada temporada otoño-invierno del año se realizó un análisis de regresión lineal y cuadrática para indicar si existió efecto del tiempo sobre el sexo, peso, ancho y grosor de músculo pectoral. A los resultados obtenidos de cada análisis con diferencia estadística (P < 0.05), se les hizo una comparación de medias mediante la prueba de Tukey de acuerdo a lo señalado por Steel y Torrie (1980). Los análisis estadísticos se realizaron con el paquete estadístico SPSS 10.0 para Windows.
RESULTADOS
Se analizaron los datos obtenidos de 244 ejemplares en total, 152 ejemplares en la primera temporada (44 hembras y 108 machos) y de 92 ejemplares en la segunda (40 hembras y 52 machos). Los resultados promedio obtenidos para el peso, ancho y grosor de músculos pectorales para cada sexo y para los dos periodos, se pueden observar en la Tabla 2. Para las variables peso y ancho de músculo pectoral no existió diferencia entre los dos años evaluados, sin embargo entre sexos hubo diferencia estadística, con mayor peso en los machos que en las hembras. Para el grosor del músculo pectoral no existió diferencia estadística entre sexos. Por otra parte, los datos de grosor del músculo pectoral indicaron que fueron mayores en el periodo 2006-2007 respecto al 2007-2008. El efecto del tiempo para cada uno de los periodos es explicado por separado para cada sexo; en algunos casos se explica a través de una regresión lineal y en otros a través de un efecto cuadrático. En la Tabla 3 se muestran las ecuaciones de regresión. El peso corporal en machos durante el primer periodo, tuvo un efecto lineal negativo (P < 0.05), es decir las aves perdieron 15.4 g por cada mes transcurrido. Sin embrago en el segundo año, el peso de los machos tuvo un efecto cuadrático lo que indica que las aves perdieron 67.8 g mensuales y a partir del cuarto mes ganaron 11.97 g (Figura 2). Al comparar los dos periodos evaluados, en el primero las aves tuvieron una pérdida de peso del 10.25 % y para el segundo periodo del 12.52 %, con una mayor pérdida de peso en el segundo año del 2.27 % respecto al primer periodo.

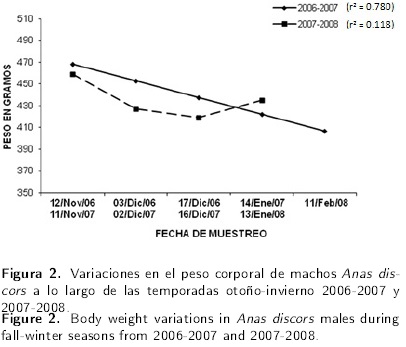
En cuanto al peso corporal de las embras perdieron mayor peso (8 g) a partir del quinto mes, no así en el siguiente año, donde se observó un efecto lineal negativo (P < 0.01), con una pérdida de peso mensual de 23.8 g. En el primer año, las hembras presentaron una pérdida de peso del 10.09 % y del 17.54 % en el segundo año, lo que indica que en el segundo periodo perdieron 7.45 % más peso con respecto al primero (Figura 3). Los datos del ancho muscular pectoral en los machos, durante el primer periodo indicaron un efecto lineal cuadrático (P < 0.05), donde estos perdieron 0.37 cm de ancho muscular los primeros 4 meses y para el quinto mes ganaron 0.45 cm. Sin embargo, para el año 2007-2008 los datos solo tuvieron un efecto lineal negativo (P = 0.059), donde perdieron 0.6 cm en cada mes transcurrido (Figura 4).

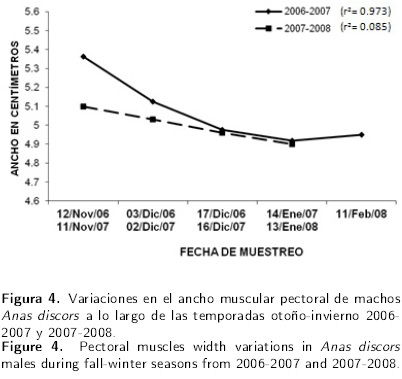
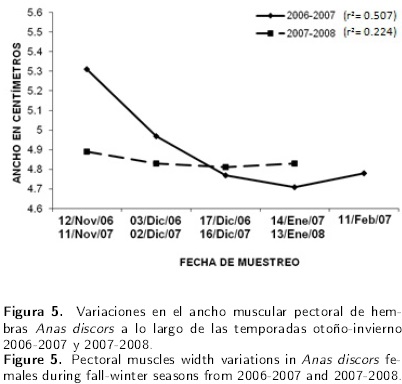
En el caso de las hembras, se puede apreciar que en el ciclo 2006-2007 existió un efecto lineal cuadrático (P < 0.01), con una disminución de 0.55 cm en los primeros cuatro meses y un aumento de 0.07 cm en el quinto mes. Por otro lado, para el ciclo 2007-2008, no existió efecto lineal cuadrático disminuyó 0.12 cm en los primeros tres meses y 0.02 cm en el cuarto mes (Figura 5). En lo que se refiere al grosor de músculos pectorales, los datos de esta variable en los machos durante el periodo 2006-2007 tuvieron un efecto lineal positivo (P > 0.05), lo que indica que en cada mes transcurrido las aves aumentaron 0.04 cm en el grosor.
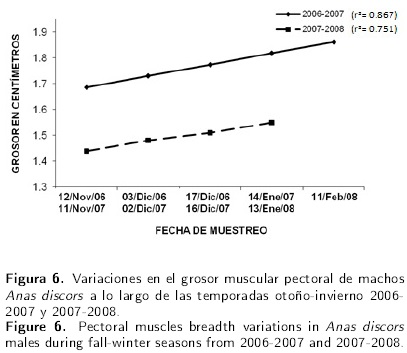
Sin embargo, para el periodo 2007-2008, no existió efecto lineal signficativo (P = 0.13) solo con una tendencia a ganar 0.034 cm por mes (Figura 6). Los datos obtenidos para el grosor de músculos pectorales en las hembras en el primer año, mostraron efecto lineal cuadrático (P < 0.05) que explica el incremento del grosor muscular de 0.056 cm en los primeros 4 meses y para el quinto mes aumentó 0.014 cm. No así para el segundo periodo, donde no existió efecto lineal cuadrático significativo (P = 0.174) (Figura 7).

En los resultados de la prueba de correlación entre las variables dependientes e independientes, se puede apreciar que existió correlación estadística entre las variables temporada-sexo (P = 0.021) y temporada-grosor (P=0.000). Por otra parte, también existió correlación entre sexo-peso (P = 0.000), sexo-ancho (P = 0.000) y sexo-grosor (P = 0.041). En cuanto a la variable mes, existió correlación entre mes-peso (P = 0.000), mes-ancho (P = 0.000) y entre én existió correlación entre peso-ancho (P = 0.000) y peso-grosor (P = 0.037). Además se encontró la existencia de una correlación entre la temporada y ancho (P = 0.059) que no fue estadísticamente significativa.
Se agruparon los esquemas elaborados según la cantidad de grasa y la distribución aproximadas (Figura 8 y 9). En la Tabla 4 se muestra el porcentaje y la proporción de aves que fueron clasificadas en cada una de las categorías, por muestreo y por temporada. Se puede apreciar un aumento casi progresivo en el porcentaje de aves con grasa escasa (+), registrándose el máximo porcentaje en el mes de febrero para la temporada 2006-2007, y en el mes de enero para 2007-2008. Además, es posible observar que el porcentaje de aves con grasa moderada (+++) fue disminuyendo a lo largo de ambas temporadas, al igual que el porcentaje de aves con (++++) en los menores porcentajes. El porcentaje de aves con grasa regular (++) también incrementó progresivamente.
DISCUSIÓN
Peso corporal
Es común que los anátidos silvestres, al igual que muchas otras especies, presenten variaciones estacionales en el peso corporal (Thompson & Baldassarre 1990); como se pudo apreciar en los resultados del presente estudio, las cercetas de ala azul presentaron, estadísticamente, un decremento gradual en el peso a lo largo de los meses de la temporada otoño-invierno (noviembre-febrero) en ambos periodos. Varios autores mencionan un patrón similar para algunas especies de aves como: patos de collar (Anas platyrhynchos) en Texas y Mississippi, EUA (Whyte & Bolen 1984, Whyte et al. 1986, Loesch et al. 1992); patos golondrinos (Anas acuta) en California, Illinois, EUA y Yucatán, México (Bellrose & Hawkins 1947, Miller 1986, Thompson & Baldassarre 1990); patos negros (Anas rubrípes) en Maine, EUA (Reinecke KJ, Stone 1982); cerceta ala verde (Anas crecca) en Texas (Baldassarre et al. 1986); cerceta ala azul (Anas discors) en Yucatán (Thompson & Baldassarre 1990); gansos nevados (Anser caerulescens) en Texas y Louisiana, EUA (Ankney 1982). En la primera temporada, la pérdida de peso para esta especie, fue del 10.1 % (43.89 g) en las hembras y 10.25 % (46.69 g) en los machos. Para la segunda temporada, la pérdida de peso total fue del 16.47 % (26.04 g) en las hembras y del 5.66 % (75 g) en los machos, registrándose, para este caso, una pérdida mayor en las hembras. La diferencia en la pérdida de peso entre sexos fue mayor en la temporada 2007-2008 en comparación con la temporada 2006-2007. Al analizar los datos disponibles y las gráficas presentadas en el reporte de Thompson y Baldassarre (1990) para la cerceta ala azul en Yucatán, se detectaron pérdidas de 1.8 % (hembras) y 2 % (machos). Los datos de White et al. (1981), mostraron pérdidas del 7 % (hembras) y 8.6 % (machos). Con lo anterior, se hace evidente que las cercetas perdieron peso en el estado de México. Esto podría deberse a: 1) la diferencia de latitudes, 2) las diferencias climáticas entre las diferentes regiones, 3) la fragmentación del humedal mexiquense, que influye sobre la calidad y cantidad del alimento disponible, 4) el incremento en el gasto de energía por aumento en la frecuencia de vuelo para buscar alimento y refugio, y 5) la competencia por el alimento entre las distintas especies de anátidos migratorios y residentes.
A pesar de que las cercetas pierden más peso en el estado de México, el peso promedio total fue mayor que el de las aves de los estudios anteriores. Por ejemplo, la cerceta de ala azul en el estado de México, pesó en promedio, 17.8 % (64.1 g) más que las cercetas de ala azul en Yucatán (Thompson & Baldassarre 1990), y 8.76 % (34.15 g) más que las de Missouri y Texas (White et al. 1981). Algunos estudios sustentan que existen variaciones entre las diferentes latitudes (Ryan 1972, Ankney 1982, Whyte et al. 1986, Thompson & Baldassarre 1990, Rave & Baldassarre 1991) y que las aves que invernan al sur tienden a ser más pesadas que las que se encuentran hacia el norte (Ryan 1972, Whyte et al. 1986; Thompson & Baldassarre 1990). En este caso, las aves del estado de México sí fueron más pesadas que las de Missouri y Texas, pero las cercetas del estado de Yucatán (Thompson & Baldassarre 1990), que se encuentran, más cerca del ecuador, resultaron ser menos pesadas que las de Missouri, Texas y el Estado de México. Por lo tanto hay discrepancia con lo reportado en la literatura.
Los datos del presente estudio en la periodo 2006-2007, muestran un ligero incremento en el peso corporal de 3.75 % (16.34 g) entre noviembre y diciembre. Estos mismos datos, pero particularmente de hembras y machos por separado, muestran que el peso aumenta (entre noviembre y diciembre) 3.76 % (17.13 g) para los machos y 3.1 % (13.39 g) para las hembras; posteriormente se observó un declive progresivo del peso hasta enero, y en febrero se apreció nuevamente un ligero incremento de 2.88 % (10.97 g), únicamente para las hembras. A pesar de que estas variaciones no fueron estadísticamente significativas, algunos autores mencionan que sus objetos de estudio presentaron un incremento en fechas similares (Bellrose & Hawkins 1947, Ankney 1982, Reinecke & Stone 1982, Whyte & Bolen 1984, Baldassarre 1986, Miller 1986, Whyte et al. 1986, Thompson & Baldassarre 1990, Loesch et al. 1992). En la periodo 2007-2008, se observa un decremento gradual del peso desde el mes de noviembre hasta enero, excepto en los machos, donde se encontró que en el mes de enero hay un ligero incremento de 2.56 % (10.83 g) comparado con el mes de diciembre. En este caso, aparentemente no hay recuperación del peso corporal en el segundo mes de la temporada como se refiere en la literatura y como se observó en la temporada anterior.
Varios autores informan que existe un aumento en el peso corporal en la temporada pre-migratoria de primavera (Ankney 1982, Whyte & Bolen 1984, Hobaugh 1985, Baldassarre et al. 1986, Whyte et al. 1986, Thompson & Baldassarre 1990); pero en este estudio no se pudo observar esta característica debido a que las aves migraron a principios de febrero.
Las variaciones de peso observadas a lo largo de la estancia de las aves en las ciénegas de Lerma se pueden asociar a la baja de peso durante el invierno por aumento del metabolismo del ave que algunos autores argumentan como una forma de adaptación para reducir el gasto energético causado por transportar su propio cuerpo, mientras más ligeras sean, menor será el gasto de energía (Reinecke & Stone 1982, Perry et al. 1986, Loesch et al. 1992).
Otros autores refieren que la pérdida de peso se debe a las inclemencias del clima como: bajas temperaturas, vientos fríos, fuertes corrientes de aire, disminución en el nivel del agua del humedal; las cuales afectan, principalmente, la disponibilidad de alimento como son las semillas, plantas acuáticas y pequeños invertebrados (Joyner et al. 1984, Paulus 1984, Tory & Chalif 1989). También puede influir la competencia por el alimento entre las distintas especies de patos migratorios de superficie en las ciénegas de Lerma como son la presencia de la cerceta ala verde (Anas crecca), pato cucharón (Anas clypeata), pato mexicano (Anas diazi), pato golondrino (Anas acuta), entre otras (SEMARNAT, 2008).
No hay pruebas científicas que expliquen como se controla esta pérdida de peso invernal, pero se especula que el metabolismo determina cuando ocurrirán las variaciones, y los factores ambientales determinan la amplitud de dichos cambios (Baldassarre et al. 1986).
Músculos pectorales
Se observó una leve disminución progresiva en el ancho muscular, pero se encontró que el grosor de los músculos incrementó desde el inicio hasta el final de la temporada. Esto coincide con las de Schwilch et al. (2002), quienes mencionaron que las aves que realizan vuelos migratorios largos, comienzan a catabolizar proteínas cuando las reservas de grasa disminuyen hasta llegar a niveles críticos, reflejándose principalmente en los músculos pectorales y algunos órganos digestivos. Varios estudios citados por este autor, mencionan que justo antes de iniciar la migración, se ha observado incremento en la masa muscular pectoral de dichas aves, que coincide con los resultados obtenidos en el grosor de los músculos pectorales en los ejemplares de cerceta de ala azul.
Flickinger y Bolen (1979), observaron atrofia muscular pectoral en gansos machos examinados en el mes de marzo a lo largo de cuatro años (19731976). Miller (1986) reportó que, en patos golondrinos (Anas acuta) de California, el peso de los músculos pectorales fue mucho menor en verano que invierno y lo relacionó a la utilización del tejido muscular durante el vuelo de migración o a la poca actividad de vuelo durante el verano.
La dinámica del músculo observada en este estudio podría deberse a un aumento de tamaño de las fibras musculares previo a una migración de larga distancia o la cantidad de ejercicio que realizarían para localizar refugio y alimento en humedales fragmentados como las Ciénegas de Lerma. A pesar de que no hay datos publicados en patos que expliquen esta observación, es posible que posean fibras musculares oxidativas, las cuales son características de las personas y animales que recorren largas distancias (Randall et al. 2001; Ganong 2006). La medición del ancho y grosor de los músculos pectorales en este estudio permite tener mayor precisión en la evaluación de la pérdida muscular con respecto a la apreciación cualitativa, y una alternativa cuando se desea evaluar la pérdida de músculo en comparación a la remoción del músculo pectoral de la quilla para evaluar el peso en gramos.
Depósitos de grasa
Este estudio mostró que la grasa subcutánea empezó a disminuir a partir del mes de enero, donde incrementó el porcentaje de ejemplares con grasa escasa (+); por el contrario, el porcentaje de aves con grasa moderada (+++) disminuyó conforme al transcurso del tiempo. Existen reportes que indican que la cantidad de grasa de la cerceta de ala azul (Anas discors), pato golondrino (Anas acuta) en Yucatán (Thomposon & Baldassarre 1990), pato de collar (Anas platyrhynchos) en Texas (Whyte & Bolen 1984, Whyte et al. 1986), y cerceta de ala verde (Anas crecca) en Texas y Louisiana (Baldassarre et al. 1986, Rave & Baldassarre 1991); se incrementa desde el arribo hasta mediados o finales de diciembre, donde se registra una disminución, la cual continúa hasta los meses de enero e incluso hasta febrero, para incrementar nuevamente justo antes de la migración de primavera.
La declinación en las reservas de grasa en este estudio, parece estar relacionado con la pérdida de peso observada en los dos periodos, y posiblemente se deba al alto gasto de energía requerido durante el invierno para mantener su termorregulación (Whyte et al. 1986, Smith & Prince 1973), y la frecuencia de vuelo para buscar refugio, alimento, y huir de los depredadores y cazadores en los humedales fragmentados de las Ciénegas de Lerma que implica un aumento del 20 % en el gasto de energía (Wooley & Owen 1977). El uso de esquemas sobre la deposición de grasa subcutánea en este estudio permite la evaluación práctica en varios ejemplares en el campo o laboratorio en comparación a la remoción y cuantificación de la grasa subcutánea por métodos químicos y físicos en el laboratorio.
La correlación negativa entre el tiempo y la condición corporal de la cerceta ala azul durante su estancia en un hábitat fragmentado como es la laguna de Chiconahuapan que pertenece a las Ciénegas de Lerma, evidencia que estas aves tienen una tendencia a mantener un comportamiento corporal similar a otras especies de anátidos migratorios en zonas neárticas y en la península de Yucatán (Thompson & Baldassarre 1990). Sin embargo, la falta de deposición de grasa en la cerceta ala azul en los meses previos a la migración en la zona de Chiconahuapan, indica que las aves no logran alcanzar su reserva energética antes de la migración, por lo que este estudio pone de manifiesto que la fragmentación de las Ciénegas de Lerma en los periodos estudiados, puede comprometer la salud y conservación de la cerceta ala azul y posiblemente de otros anátidos migratorios y residentes.
AGRADECIMIENTOS
Se agradece al Biólogo Pedro Esteban Díaz Díaz y a la Dirección General de Vida Silvestre de la Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT) del Distrito Federal y del Estado de México por el apoyo brindado durante el desarrollo del proyecto; así como la cooperación de los compañeros y colegas Sofía González, Edgar Arturo Cuevas, Ernesto Godinez, José Manuel Carranza, Sarahí López, Mariemma Escorcia, Eduardo Aguilar, Rodrigo Barrón, Elia Armas, Alejandra Domínguez, Alma Cruz, Karla Irazema Moreno, Francisco Javier Barrón y Adelfo Juárez, durante la recolección y el procesamiento de las muestras.
LITERATURA CITADA
Ankney CD (1982) Annual Cycle of Body Weight in Lesser Snow Geese. Wildlife Society Bulletin 10: 60-64. [ Links ]
CONABIO (2007a) Regionalización: Listado de Regiones Hidrológicas Prioritarias, 65. Cabecera del Río Lerma. CONABIO. www.conabio.gob.mx (consultado el lunes 14 de marzo de 2007 [ Links ]
CONABIO (2007b) Regionalización. AICAS: Ciénegas de Lerma. CONABIO. http:www.conabio.gob.mx (consultado el lunes 14 de marzo de 2007). [ Links ]
CND (2007c) Situación Actual de la Cuenca del Río Lerma, Conocimiento y Derechos de las Comunidades Originarias. CND. www.cnd.or.mx (consultado el lunes 14 de marzo de 2007). [ Links ]
Baldassarre GA, Whyte RJ, Bolen EG (1986) Body Weight and Carcass Composition of Nonbreeding Green-winged Teal on the Southern High Plains of Texas. Journal of Wildlife Management 50: 420-426. [ Links ]
Bellrose FC, Hawkins AS (1947) Duck Weights in Illinois. Auk 64: 422-430. [ Links ]
Flickinger EL, Bolen EG (1979) Weights of lesser snow geese taken on their winter range. Journal of Wildlife Management 43: 531-533. [ Links ]
Ganong WF (2006) Fisiología Médica. Vigésima edición. El Manual Moderno. México D.F. 745 pp. [ Links ]
Gill FB (2001) Ornithology. 2nd edition. WH Freeman and Company. USA. 766 pp. [ Links ]
Harrison GJ and Ritchie BW (1994) Making distinctions of physical examination. In: Avian Medicine: Principles and application. (Ed). Ritchie BW, Harrison GJ, Harrison. 1st edition. Wingers Publishings. LR Florida, USA. 1384 pp. [ Links ]
Hobaugh WC (1985) Body Condition and Nutrition of Snow Geese Wintering in Southeastern Texas. Journal of Wildlife Management 49: 1028-1037. [ Links ]
Howard R, Moore A (1998) A Complete Check List of the Birds of the World. 2nd edition. Academic Press. New York, USA. 640 p. [ Links ]
Joyner DE, Arthur RD, Jacobson BN (1991) Winter Weight Dinamics, Grain Consuption and Reproductive Potential in Canada Geese. Condor 86: 275-280. [ Links ]
Loesch CR, Kaminski RM, Richardson DM (1992) Endogenous Loss of Body Mass by Mallards in Winter. Journal of Wildlife Management 56: 735-739. [ Links ]
Miller MR (1986) Northern Pintail Body Condition During Wet and Dry Winters in the Sacramento Valley, California. Journal of Wildlife Management 50: 189-198. [ Links ]
Paulus SL (1984) Activity Budgets of Nonbreeding Gadwalls in Louisiana. Journal of Wildlife Management 48: 371-380. [ Links ]
Perry MC, Kuenzel WJ, Williams BK, Serafín JA (1986) Influence of Nutrients on Feed Intake and Condition of Captive Canvasback in Winter. Journal of Wildlife Management 50: 427-434. [ Links ]
Price J, Droege S, Price A (1995) The Summer Atlas of North American Birds. Academic Press. New York, USA. 364 pp. [ Links ]
Randall D, Warren B, French K (2001) Animal Physiology: Mechanisms and Adaptation. 5th edition. W.H. Freeman and Company. New York, USA. 752 pp. [ Links ]
Rave DP, Baldassarre GA (1991) Carcass Mass and Composition of Green-winged Teal Wintering in Louisiana and Texas. Journal of Wildlife Management 55: 457-461. [ Links ]
Reinecke KJ, Stone TL (1982) Seasonal Carcass Composition and Energy Balance of Female Black Ducks in Maine. Condor 84: 420-426. [ Links ]
Ryan RA (1972) Body Weight and Weight Changes of Wintering Diving Ducks. Journal of Wildlife Management 36: 159-165. [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (2008) Estrategia para la conservación, manejo y aprovechamiento de las aves acuáticas y su hábitat en México. 1ra edición. SEPRIM. Distrito Federal, México. 2008. [ Links ]
Schwilch R, Grattarola A, Spina F, Jenni L (2002) Protein loss during long-distance migratory flight in passerine birds: adaptation and constraint. The Journal of Experimental Biology 205: 687-695. [ Links ]
Sibley AG (2001) The Sibley Guide to Birds. 1st edition. Alfred A. Knopf. New York, USA. 544 pp. [ Links ]
Smith KG, Prince HH (1973) The Fasting Metabolism of Sub adults Mallards Acclimatized to Low Ambient Temperatures. Condor 75: 330-335. [ Links ]
Steel RCD, Torrie JH (1980) Principles and Procedures of Statistics. 2nd edition. McGraw Hill. New York, USA. 633 pp. [ Links ]
Thompson JD, Baldassarre GA (1990) Carcass Composition of Nonbreeding Blue-winged Teal and Northern Pintails in Yucatan, Mexico. Condor 92: 1057-1065. [ Links ]
Tory PR and Chalif E (1989) Aves de México. 1ra edición. Diana. México. 473 pp. [ Links ]
Vázquez RH (2004) Preferencias de Hábitat por la avifauna presente en la Laguna de Chimaliapan, Ciénegas de Lerma, Lerma, Estado de México [tesis de maestría]. Estado de México, México: Universidad Nacional Autónoma de México. 131 pp. [ Links ]
White DH, King KA, Mitcheli CA, Krynitsky AJ (1981) Body Lipids and Pesticide Burdens of Migrant Blue-Winged Teal. Journal Field Ornithology 52: 23-28. [ Links ]
Whyte RJ, Bolen EG (1984) Variation in Winter Fat Depots and Condition Indices of Mallards. Journal of Wildlife Management 48: 1370-1373. [ Links ]
Whyte RJ, Baldassarre GA, Bolen EG (1986) Winter Condition of Mallards on the Southern High Plains of Texas. Journal of Wildlife Management 50: 52-57. [ Links ]
Wooley Jr JB, Owen Jr RB (1977) Metabolic rates and heart rate-metabolism relationships in the Black Duck (Anas rubripes). Comparative Biochemistry and Physiology A 57: 363-367 [ Links ]

