










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkUniversidad y ciencia
versão impressa ISSN 0186-2979
Universidad y ciencia vol.27 no.1 Villahermosa Abr. 2011
Artículos
Crecimiento de alevines de Cichlasoma urophthalmus con dietas basadas en diferentes niveles de inclusión de proteína de soya y gluten de trigo
Growth of Cichlasoma urophthalmus fingerlings with diets based on different levels of wheat gluten and soybean protein
C Villarreal1, R Gelabert1, G Gaxiola2, G Cuzon3, LE Amador1, E Guevara1, R Brito1 *
1 DES Ciencias Naturales y Exactas, UNACAR Calle 56 No. 4 x Ave Concordia. Ciudad del Carmen 24118. Campeche, México. (CV)(RG)(LEA)(EG)(RB). * Correo electrónico: rbrito@pampano.unacar.mx
2 Unidad Multidisciplinaria de Docencia e Investigación. Facultad de Ciencias, UNAM. (GG)
3 COP-Tahití-IFREMER. (GC)
Artículo recibido: 10 de septiembre de 2009,
Aceptado: 07 de marzo de 2011
RESUMEN
Se determinó el efecto de seis dietas con diferente contenido de proteína de origen vegetal sobre el crecimiento de alevines de Cichlasoma urophthalmus. Se utilizaron dietas isoenergéticas e isolipídicas con contenido de proteína entre 10% y 60%, empleando soya y gluten de trigo como las principales fuentes proteicas. Los tratamientos no afectaron la supervivencia, la cual fue superior al 80% en todos los casos. Los índices de crecimiento variaron significativamente de acuerdo al contenido proteico de las dietas, siendo la tasa de crecimiento específica en las dietas con más de 30% de proteína comparable a las obtenidas por otros autores utilizando proteínas de origen animal.
Palabras clave: Proteína vegetal, tasa de crecimiento, Cichlasoma urophthalmus.
ABSTRACT
The effect of six diets with a different plant protein content on the growth of Cichlasoma urophthalmus fingerlings, was determined. Isoenergetic and isolipidic diets with a protein content of 10% to 60% were formulated using soybean and wheat gluten as the main protein sources. The treatments did not affect survival, which was above 80% in all cases. The growth indices varied significantly according to the protein content of the diets, and the specific growth rate with the diets with a protein content greater than 30% was comparable to that obtained by other authors using animal protein.
Key words: Plant protein, growth rate, Cichlasoma urophthalmus.
INTRODUCCIÓN
Cichlasoma urophthalmus (Günther, 1862), es un pez perteneciente a la familia Cichlidae, vive tanto en aguas dulces como salobres y se distribuye en México desde el Río Coatzacoalcos hasta Isla Mujeres y en Centroamérica se ha reportado su presencia en Belice, Guatemala, Honduras y Nicaragua (Chávez-López et al. 2005).
Al analizar la alimentación natural de esta especie Caso-Chávez et al. (1986) hallaron al detrito como la entidad más representada en los estómagos de los juveniles. Por otra parte, Martínez-Palacios & Ross (1988) describieron a la especie como principalmente carnívora, refiriendo que la estructura del tracto digestivo indica que estos organismos no digieren eficientemente el material vegetal. En contraste con estos autores, Chávez-López et al. (2005), consideraron a C. urophthalmus como principalmente herbívora, aunque su dieta varió estacionalmente de acuerdo a la disponibilidad de alimento, mientras Bergmann & Motta (2005) refieren que esta especie se alimenta fundamentalmente de peces y algas filamentosas, con algunos cambios en la dieta durante su ontogenia. Recientemente, Guevara et al. (2007) encontraron en el contenido estomacal de C. urophthalmus un predominio de materia vegetal y materia orgánica no identificada.
Estos resultados muestran una gran flexibilidad en la dieta de C. urophthalmus, que dependiendo de la disponibilidad de alimento y de su ontogenia puede ser considerada una especie carnívora, omnívora o herbívora.
C. urophthalmus se pesca de forma artesanal en varias regiones de México y tiene potencial para acuacultura en aguas dulces y salobres (Chávez-López et al. 2005), por lo que también se han estudiado aspectos de su nutrición en cautiverio.
Experimentos de digestibilidad in vivo han mostrado que el trigo puede ser usado como fuente principal de carbohidratos y que la harina de soya se puede usar como un sustituto parcial de proteína de pescado en dietas para la especie (Martínez-Palacios 1988). Chávez (1990), determinó los requerimientos de vitamina C, además se estimaron los requerimientos de lípidos, ácidos grasos y aminoácidos esenciales (Martínez-Palacios & Ross 1994). Estos autores también hicieron recomendaciones en los niveles de inclusión de carbohidratos y fibra en las dietas para C. urophthalmus. Martínez-Palacios et al. (1996a) determinaron el requerimiento de proteína de los juveniles de la especie utilizando harina de pescado como fuente proteica y Chávez-Sánchez et al. (2000), encontraron los niveles óptimos para el fósforo y calcio.
La harina de pescado es muy utilizada en la acuicultura debido a que es una fuente excelente de aminoácidos, ácidos grasos esenciales, vitaminas, minerales y además incrementa la palatabilidad de las dietas (Samocha et al. 2004). La producción de esta materia prima se ha mantenido prácticamente constante en los últimos años, mientras que su demanda ha ido en aumento, incrementándose la presión sobre las pesquerías y los precios (Saoud et al. 2008), por lo que es necesario encontrar alternativas al uso de la harina de pescado en dietas para la acuacultura.
Teniendo en cuenta la información sobre los hábitos alimentarios y estudios de nutrición en la especie, así como la necesidad de encontrar alternativas a la harina de pescado como fuente proteica en alimentos para la acuacultura, se consideró que es posible sustituir la harina de pescado por proteína de origen vegetal para cubrir los requerimientos proteicos de los alevines de esta especie, por lo que se elaboraron dietas con diferentes niveles de proteína de soya y gluten de trigo, con el objetivo de probar su efecto sobre el crecimiento de C. urophthalmus y compararlo con las tasas de crecimiento obtenidas en estudios donde se utilizó harina de pescado como fuente proteica.
MATERIALES Y MÉTODOS
Dietas experimentales
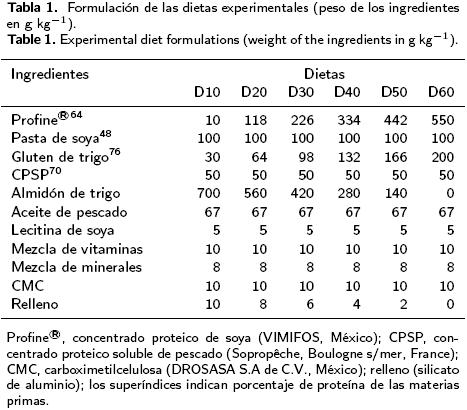
Se formularon y elaboraron seis dietas isolipídicas e isoenergéticas con un contenido teórico de 10%, 20%, 30%, 40%, 50% y 60% de proteína, utilizando como fuente proteica Profine® (concentrado proteico de soya,VIMIFOS, México), pasta de soya y gluten de trigo. Se incluyó 5% de CPSP70 (concentrado proteico soluble de pescado, Sopropêche, Boulogne s/mer, Francia) como atrayente. En la Tabla 1 aparece la formulación de las dietas experimentales.
Para la confección de las dietas se mezclaron completamente los ingredientes secos con los aceites y se agregó agua hasta que se obtuvo una masa firme. La masa se pasó a través de un molino de carne (Torrey modelo M32, E.U.) equipado con un dado de 2 mm y los pellets resultantes fueron secados en una estufa (Felisa modelo 291, México) con aire a 60 °C durante 12 h. Después de secos, los pellets se guardaron en bolsas herméticamente cerradas y etiquetadas a -4 °C hasta su uso.
La determinación de la composición proximal de las dietas se realizó utilizando los métodos de la AOAC (1990) y fueron realizadas por los laboratorios del Centro de Control Agroindustrial S.A. de C.V. (México). Los valores de contenido de energía de las dietas se determinaron mediante combustión en bomba calorimétrica modelo Parr previamente calibrada con ácido benzoico.
Para el cálculo de los porcentajes de los diferentes nutrientes en las dietas se utilizó el programa Excel diet formulation/nutrient calculation (Wilkinson 2008).
Origen de los animales experimentales
Los alevines se obtuvieron de reproductores mantenidos en el laboratorio. Se cosecharon dos desoves que ocurrieron simultáneamente, pasándose las larvas a un acuario de 200 L con aireación (manteniendo niveles por encima del 80% de saturación), posteriormente los alevines se colocaron en un tanque cilindro cónico de 1750 L acoplado a un sistema de recirculación con filtro de arena, filtro de cartucho de 100 μm (Pentair Aquatics® modelo VF12RS, E.U.), lámpara de luz UV (Pentair Aquatics® modelo QL40, E.U.), filtro biológico, eliminador de espuma y filtro semi-húmedo.
Durante los primeros siete días los animales se alimentaron con nauplios de Artemia (1 nauplio ml–1) y alimento artificial en polvo (iniciación #0 para trucha marca El Pedregal, con 52 % de proteína y tamaño menor a 600 μm). Durante los siguientes 8 d los alevines se alimentaron con alimento para trucha en polvo a razón del 15 % de la biomasa dividido en tres raciones (08:00, 14:00 y 20:00), posteriormente hasta los 21 d se alimentaron con una mezcla (50/50) de alimento iniciación #0 para trucha y alimento Tilapia MKT 30%, molido a tamaño de 500 μm a razón del 10 % de la biomasa dividido en tres raciones (08:00, 14:00 y 20:00).
Corrida experimental
Después de 21 d de cultivo, se colocaron 25 alevines (largo total inicial 1.68 ± 0.20 cm, peso húmedo inicial 88.1 ± 32.8 mg, peso seco inicial 21.1 ± 0.8 mg) en 18 acuarios de 40 L de capacidad conectados a un sistema de recirculación con filtro biológico, se asignó el tratamiento correspondiente a cada acuario de forma aleatoria considerándose tres réplicas por cada tratamiento (dieta).
Para la determinación del largo y peso inicial se sacrificaron 25 ejemplares (seleccionados al azar del lote inicial), a los cuales se les midió el largo total utilizando un vernier y tanto el peso húmedo como seco se determinó con una balanza analítica (Mettler Toledo modelo AL 104), con 0.0001 g de precisión. Para obtener el peso seco los peces fueron colocados en una estufa (Felisa modelo B131AD, México) a 60 °C durante 48 h, posteriormente se llevaron a temperatura ambiente en una desecadora (Corning Pyrex, E.U.) antes de pesarlos. El alimento confeccionado fue molido y tamizado para obtener migajas entre 500 μm y 1000 μm, consideradas adecuadas para el tamaño de los alevines. Los peces se alimentaron a saciedad tres veces por día (08:00, 14:00 y 20:00) durante el transcurso de todo el experimento.
Diariamente se realizó la limpieza de los acuarios mediante sifoneo y se renovó alrededor del 7% del agua del sistema con agua potable previamente declorada con aireación. Todos los días se registró el comportamiento del oxígeno disuelto con un oxímetro marca YSI modelo 75. La temperatura durante el experimento se registró con cuatro sensores de temperatura (Hobo® U22 Water Temp Pro v2), con una resolución de 0.02 °C, distribuidos en diferentes recipientes experimentales y programados para tomar registros cada una hora. Para las mediciones de amonio y nitritos se utilizó un kit Tetratest.
Después de 40 d de experimentación, se sacrificaron los peces bajándoles la temperatura y se les determinó el largo total, peso húmedo y peso seco a todos los organismos, siguiendo el método descrito anteriormente. Con los datos de peso obtenidos se determinó el crecimiento absoluto (CA) y relativo (CR) y las tasas de crecimiento absoluto (TCA) y relativo (TCR), según Busacker et al. (1990) y la tasa instantánea (o específica) de crecimiento (TCE) (Ricker 1979):
TCE = [(Ln Y2 - Ln Y1)/ (t2-t1)]·100
Donde: Ln Y1 y Ln Y2 son el logaritmo natural del peso al inicio y al final de la fase de crecimiento, t2-t1 la duración del experimento en días.
Análisis estadístico
Los índices de crecimiento se contrastaron con análisis de varianza (ANOVA) y posteriormente con la prueba Tukey donde se determinaron las diferencias entre tratamientos. Los datos de porcentaje de supervivencia se transformaron mediante arcoseno de raíz de x, para normalizarlos antes de realizar el ANOVA. En todos los casos se realizó una prueba de Kolmogorov-Smirnov para comprobar la normalidad y la prueba Levene para comprobar la homogeneidad de varianza previo al ANOVA y prueba Tukey (Zar 1984).
RESULTADOS
En la Tabla 2 se muestra la composición proximal de las dietas experimentales. La temperatura del agua durante el experimento fluctuó entre 26.70 °C y 30.40 °C con valor medio de 28.20 ± 0.67 °C. La concentración de oxígeno del agua se mantuvo entre 6.30 mg L–1 y 7.84 mg L–1 con valor promedio de 7.28 ± 0.85 mg L–1. Estos valores indican que la concentración de oxígeno estuvo por encima del 80% del valor de saturación durante todo el experimento. Los valores de amonio se mantuvieron por debajo del límite de detección del test utilizado, mientras que las concentraciones de nitrito permanecieron por debajo de 0.1mg L–1. Las dietas no afectaron la supervivencia (ANOVA; F=2.19, P=0.1229), la cual fue superior al 80 % en todos los tratamientos.

Los tratamientos tuvieron un efecto significativo sobre todos los índices de crecimiento calculados para los alevines de C. urophthalmus (Tabla 3). Al analizar los diferentes índices se observa que en todos los casos hay una tendencia a un incremento en sus valores según incrementa el porcentaje de proteínas en las dietas (de D10 a D60), con valores significativamente más bajos en el tratamiento D10 y más altos en el tratamiento D60.
En la Tabla 4 aparece la composición de las dietas experimentales según el análisis proximal y los cálculos realizados mediante el programa de Wilkinson (2008), así como los requerimientos o valores recomendados para diferentes nutrientes según Martínez-Palacios & Ross (1994) y Chávez-Sánchez et al. (2000). Como se puede observar, las dietas D30 a D60 cubrieron los requerimientos proteicos, mientras que las dietas de la D10 a la D40 tuvieron mayor porcentaje de carbohidratos que los recomendados. En cuanto a lípidos totales, todas las dietas excepto la D10, cumplieron con los requerimientos; sin embargo, en todos los casos hay menor proporción de ácidos grasos poliinsaturados de la serie n-6. Respecto a los aminoácidos esenciales, las dietas de la D30 a la D60 cubrieron en general los requerimientos. Según los cálculos realizados y los datos de Chávez-Sánchez et al. (2000), todas las dietas tuvieron deficiencia en los contenidos de fósforo y calcio.
DISCUSIÓN
La temperatura es uno de los factores ambientales que tiene mayor influencia en el metabolismo y el crecimiento de los peces. Martínez-Palacios et al. (1996b), encontraron que en C. urophthalmus se reduce la tasa de ingestión y en consecuencia la tasa de crecimiento y la eficiencia de conversión del alimento en temperaturas por debajo de 27 °C. En relación a la concentración de oxígeno Martínez-Palacios & Ross (1986) señalan que a 28 °C las condiciones de hipoxia en esta especie aparecen en concentraciones de oxígeno por debajo de 3.5 mg L–1, sin afectaciones en la tasa respiratoria en niveles superiores a este valor crítico. De acuerdo a esto, la temperatura y la concentración de oxígeno disuelto se mantuvieron en valores adecuados para la especie durante este experimento.
C. urophthalmus ha sido catalogada como una especie con un comportamiento agresivo y territorial, a lo que se atribuye la mortalidad registrada en diferentes estudios (Chávez, 1990; Martínez-Palacios et al., 1996b; Chávez-Sánchez et al., 2000), sin embargo en este trabajo se obtuvo una buena supervivencia en todos los tratamientos, posiblemente debido al pequeño tamaño de los peces utilizados.
El reemplazo de la harina de pescado por diferentes proporciones de proteínas de origen vegetal en dietas para peces teleósteos se ha llevado a cabo con éxito en numerosas especies desde hace años (Tacon 1993); sin embargo, sólo han aparecido unos pocos reportes de la utilización de proteína de origen vegetal como única fuente proteica en dietas para peces (Fontaínhas-Fernades et al. 1999). Kaushik et al. (2004) lograron sustituir hasta el 98% de la harina de pescado por proteína vegetal en dietas para Dicentrarchus labrax, sin encontrar efectos sobre la tasa de ingestión, la tasa de crecimiento, la eficiencia alimenticia ni la ganancia diaria en nitrógeno.
En este trabajo se utilizaron dietas basadas principalmente en proteínas de origen vegetal, la inclusión de PSPS70 (proteína soluble de pescado), se realizó en un nivel de 5% en todas las dietas con función de atrayente, no como fuente proteica, representando un aporte de entre el 10% y 5% del contenido de proteína total en las dietas D40 a la D60. Aunque no se cuantificó la tasa de ingestión, sí se pudo observar durante la corrida experimental que todas las dietas fueron bien aceptadas por los alevines de C. urophthalmus.
Papatryphon & Soares Jr. (2000), encontraron que el suplemento de 2% de proteínas solubles de pescado en dietas basadas en insumos de origen vegetal, incrementó significativamente la ingestión de alimento en Morone saxatilis. Sin embargo, en juveniles de Micropterus salmoides, la adición de atrayentes, como proteínas solubles de pescado, a dietas basadas en proteínas vegetales, no produjo ningún efecto en el peso final, tasa de ingestión, ganancia en peso o tasa de conversión del alimento (Oliveira & Cyrino, 2004).
Martínez-Palacios et al. (1990), obtuvieron una tasa específica de crecimiento de 1.88, utilizando una dieta de 40% de proteína en base a harina de pescado, en alevines de C. urohthalmus con pesos iniciales mayores (519 mg peso húmedo) que los utilizados en este trabajo. Las tasas de crecimiento de los organismos de los tratamientos D30 a D60 resultaron superiores a este valor, lo que sugiere que los animales tuvieron un crecimiento adecuado, respondiendo bien a dietas basadas en proteínas de origen vegetal. En otro trabajo Martínez-Palacios et al. (1996a), con animales con peso húmedo inicial de 150 mg, obtuvieron tasas de crecimiento entre 1.18 y 4, dependiendo de los porcentajes de proteína de dietas confeccionadas a partir de harina de macarela. Los mejores valores del índice obtenidos en este trabajo coinciden con los que reportan estos autores para las dietas que tenían entre 20% y 25% de proteína, valor cercano al 30% de proteína que señalan estos autores como requerimiento de la especie.
Utilizando dietas de 40 % de proteína basadas en caseína y con ejemplares de alrededor de 400 mg de peso inicial, Chávez-Sánchez et al. (2000) obtuvieron tasas específicas de crecimiento entre 1.2 y 2.74 dependiendo de los niveles de calcio y fósforo de las dietas, resultados comparables a los obtenidos en este trabajo (TEC de 1.2 a 2.9), confirmando que C. urophthalmus puede utilizar bien dietas donde la proteína proviene principalmente de fuentes vegetales. La información de la digestibilidad y palatabilidad de los ingredientes y la utilización e interferencia entre nutrientes constituyen un punto crucial en la evaluación de ingredientes para dietas de las especies en cultivo (Glencross et al. 2007). Martínez-Palacios (1988) estudió la digestibilidad de dietas elaboradas con diferentes ingredientes en C. urophthalmus, no encontrando diferencias en la digestibilidad de proteínas de diferentes orígenes (trigo, soya y harina de pescado). Este autor tampoco encontró ningún efecto negativo en la digestibilidad de proteínas debido a la presencia de factores anti-nutricionales que pueden estar presentes en las materias primas de origen vegetal.
La digestibilidad de carbohidratos también resultó alta en C. urophthalmus (Martínez-Palacios, 1988), siendo significativamente menor solamente en la dieta donde la fuente de carbohidratos fue la soya, que se caracteriza por su contenido de oligo-sacáridos (sacarosa, rafinosa y estaquiosa). La sacarosa generalmente es digerible para los organismos acuáticos, pero la rafinosa y la estaquiosa no son digestibles debido a la carencia de α-galactosidasas en peces y según Gatlin III et al. (2007), esto puede constituir uno de los mayores retos en el uso de la soya en dietas para peces, ya que puede afectar negativamente la utilización de nutrientes y reducir la eficiencia alimentaria. Entre los carbohidratos de la soya además se encuentran polisacáridos no amiláceos (mayormente hemicelulosa y pectina), pero no almidón, estos polisacáridos pudieran actuar como antinutrientes tanto en peces carnívoros como herbívoros (Krogdhal et al. 2005).
La mayoría de las dietas utilizadas en este trabajo (Tabla 4), tuvieron más carbohidratos que los recomendados por Martínez-Palacios & Ross (1994) y una parte de estos carbohidratos provenían de la pasta de soya, esto pudo haber afectado el crecimiento de los organismos, a pesar de que las dietas cubrieran los requerimientos de proteína. No obstante, según los resultados de este estudio, el crecimiento de los alevines de C. urophthalmus estuvo directamente relacionado con el nivel de proteína de las dietas e inversamente relacionado con el nivel de inclusión de carbohidratos, independientemente de su origen (pasta de soya o almidón de trigo). En C. urophthalmus la digestibilidad de las dietas disminuyó cuando se incrementó el porcentaje de fibra en las formulaciones (Martínez-Palacios 1988), por lo que este autor recomienda mantener el nivel de fibra por debajo del 5 % al respecto, ninguna de las dietas utilizadas en este trabajo sobrepasó este valor.
En la literatura hay pocos estudios sobre los requerimientos de C. urophthalmus, específicamente cuando se trata de aminoácidos. Sólo se cuenta con los estimados por Martínez-Palacios & Ross (1994), basándose en la composición del cuerpo y en los requerimientos de tilapia. Al comparar los resultados aquí obtenidos con estos reportes (Tabla 4), se observa que las dietas que tuvieron porcentajes de proteína por encima de 30% (D30 a D60), cubren en general con los requerimientos de aminoácidos de la especie. Sin embargo, todas las dietas tuvieron deficiencias en calcio y fósforo según Chávez-Sánchez et al. (2000), estos autores refieren que C. urophthalmus tiene altos requerimientos de estos nutrientes si se comparan con otras especies. Las tasas de crecimiento obtenidas en este trabajo son comparables a las referidas por Chávez-Sánchez et al. (2000) y los valores de requerimiento determinados en C. urophthalmus son el doble de los determinados para otros peces, por ejemplo: Oreochromis aureus (Robinson et al. 1984, 1987), Morone saxatilis (Dougall et al. 1996), Chanos chanos (Borlogan & Sato 2001), Melanogrammus aeglefinus (Roy & La l l 2003). Teniendo en cuenta lo anterior y que el método utilizado por Chávez-Sánchez et al. (2000) para calcular el requerimiento (línea quebrada), se ha discutido como inadecuado (Shearer 2000), es posible que los requerimientos de estos nutrientes hayan sido sobrestimados y que las dietas utilizadas en este trabajo aportaran calcio y fósforo en las cantidades requeridas por los alevines de C. urophthalmus.
Otra deficiencia de las dietas utilizadas en este trabajo de acuerdo a lo recomendado por Martínez-Palacios & Ross (1994), estuvo relacionada con las cantidades de ácidos grasos insaturados de la serie n-6, debido a que las materias primas de origen vegetal utilizadas aportaron pocos lípidos, lo que indica que sería necesario incrementar el nivel de inclusión de aceites vegetales en las dietas. Como los requerimientos de ácidos grasos no han sido establecidos para C. urophthalmus, Martínez-Palacios & Ross (1994) basaron su recomendación en los requerimientos de Tilapia y otros cíclidos con proporción de 1 % de n-6 y 1 % de n-3. Según Dabrowski & Portella (2006), existe discrepancia en cuanto a que los ácidos grasos de ambas series sean esenciales para los cíclidos ya que se ha demostrado que con un suplemento de altos niveles de n-3 se obtienen los mejores crecimientos en tilapia.
Esta investigación demostró que los alevines de C. urophthlamus alimentados con dietas con más de 30% de proteína elaboradas con la combinación de la pasta de soya, el concentrado proteico de soya y el gluten de trigo, presentaron supervivencias y tasas de crecimiento comparables a estudios en los que se ha empleado proteína de origen animal.
LITERATURA CITADA
AOAC (1990) Official Methods of Analysis of the Association of Official Analytical Chemists, 15th edition, AOAC: Washington, DC. 1141 pp. [ Links ]
Bergmann GT, Motta PJ (2005) Diet and morphology through ontogeny of the nonindigenous Mayan cichlid 'Cichlasoma (Nandopsis)' urophthalmus (Günther 1862) in southern Florida. Environmental Biology Fisheries 72: 205-211. [ Links ]
Borlongan IG, Satoh S (2001) Dietary phosphorus requirement of juvenile milkfish, Chanos chanos (Forsskal). Aquaculture Research 32: 26-32. [ Links ]
Busacker PG, Adelman RI, Goollish ME (1990) Growth. 363-387. En: Schreck BC, Moyle BP (Ed). Methods for Fish Biology. American Fisheries Series 13. Great Britain. 684 pp. [ Links ]
Caso-Chávez M, Yañez-Arancibia A, Lara-Domínguez AL (1986) Biología, ecología y dinámica de poblaciones de Cichlasoma urophthalmus (Günther) (Pices: Cichlidae) en hábitat de Thalassia testudinum y Rhizophora mangle, Laguna de Términos, sur del Golfo de México. Biótica 11(2): 79-111. [ Links ]
Chávez MC (1990) Vitamin C requirement of the Mexican native cichlid Cichlasoma urophthalmus (Günther). Aquaculture 86: 409-416. [ Links ]
Chávez-López R, Peterson MS, Brown-Peterson NJ, Morales-Gómez AA, Franco-López J (2005) Ecology of the mayan cichlid, Cichlasoma urophthalmus Günther, in the Alvarado lagoonal system, Veracruz, Mexico. Gulf Caribbean Research. 17: 123-131. [ Links ]
Chávez-Sánchez C, Martínez-Palacios CA, Martínez-Pérez G, Ross LG (2000) Phosphorus and calcium requirements in the diet of the American cichlid Cichlasoma urophthalmus (Günther). Aquaculture Nutrition. 6: 1-9. [ Links ]
Dabrowski K, Portella MC (2006) Feeding Plasticity and Nutritional Physiology in Tropical Fishes En: Val AL, Almeida-Val VMF & Randall DJ (eds) The Physiology of Tropical Fishes, Vol. 21. Elsevier, London. 634 pp. [ Links ]
Dougall DS, Woods III LC, Douglass LW, Soares JH (1996) Dietary phosphorus requirement of juvenile striped bass Morone saxatilis. Journal of the World Aquaculture Society 27: 82-91. [ Links ]
Fontaínhas-Fernades A, Gomes E, Reis-Henriques MA, Coimbra J (1999) Replacement of fish meal by plant proteins in the diet of Nile tilapia: digestibility and growth performance. Aquaculture International. 7: 57-67. [ Links ]
Gatlin III DM, Barrows FT, Brown P, Dabrowski K, Gaylord TG, Hardy RW, Herman E, Hu G, Krogdahl Å, Nelson R, Overturf K, Rust M, Sealey W, Skonberg D, Souza EJ, Stone D, Wilson R, Wurtele E (2007) Expanding the utilization of sustainable plant products in aquafeeds: a review. Aquaculture Research. 38: 551-579. [ Links ]
Glencross BD, Booth M, Allan GL (2007) A feed is only as good as its ingredients - a review of ingredient evaluation strategies for aquaculture feeds: Aquaculture Nutrition. 13: 17-34. [ Links ]
Guevara E, Sánchez AJ, Rosas C, Mascaró M, Brito R (2007) Asociación trófica de peces distribuidos en vegetación acuática sumergida en Laguna de Términos, sur del Golfo de México, Univ. Cienc. 23 (2): 151-166. [ Links ]
Kaushik SJ, Covès D, Dutto G, Blanc D (2004) Almost total replacement of fish meal by plant protein sources in the diet of a marine teleost, the European seabass, Dicentrarchus labrax. Aquaculture 230: 391-404. [ Links ]
Krogdhal Å, Hemre GI, Mommsen TP (2005) Carbohydrates in fish nutrition: digestion and absorption in postlarval stages. Aquaculture Nutrition. 11: 103-22. [ Links ]
Martínez-Palacios CA (1988). Digestibility studies in juveniles of the Mexican cichlid, Cichlasoma urophthalmus (Günther). Aquaculture and Fisheries Management 19: 347-354. [ Links ]
Martínez-Palacios CA, Chávez-Sánchez C, Ross LG (1996b). The effects of water temperature on food intake, growth and body composition of Cichlasoma urophthalmus (Günther) juveniles. Aquaculture Research. 27: 455-461. [ Links ]
Martínez-Palacios CA, Harfush-Meléndez M, Chávez-Sánchez C, Ross LG (1996a) The optimum dietary protein level for the Mexican cichlid Cichlasoma urophthalmus (Günther): A comparison of estimates derived from experiments using fixed-rate feeding and ad-libitum feeding. Aquaculture Nutrition. 2: 11-20. [ Links ]
Martínez-Palacios CA, Ross LG (1986) The effects of temperature, body weight and hypoxia on the respiratory rate of the Mexican mojarra (Cichlasoma urophthalmus). Aquaculture and Fisheries Management 17: 243-248. [ Links ]
Martínez-Palacios CA, Ross LG (1988) The feeding ecology of the Central American cichlid Cichlasoma urophthalmus (Günther). Journal of Fisheries and Biology. 33(5): 665-670. [ Links ]
Martínez-Palacios CA, Ross LG (1994) Biología y Cultivo de la mojarra Latino-Americana Cichlasoma urophthalmus. CONACyT, México. 250 pp. [ Links ]
Martínez-Palacios CA, Ross LG, Rosado M (1990) The effects of salinity on the survival and growth of juvenile Cichlasoma urophthalmus. Aquaculture 91: 65-75. [ Links ]
Oliveira AMBMS, Cyrino JEP (2004) Attractants in plant protein-based diets for the carnivorous largemouth bass Micropterus salmoides. Science Agriculture. (Piracicaba, Braz.) 61(3): 326-331. [ Links ]
Papatryphon E, Soares Jr JH (2000) Identification of feeding stimulants for striped bass, Morone saxatilis. Aquaculture 185: 339-352. [ Links ]
Ricker W (1979) Growth rates and models. 677-743 pp In: Hoar, W., Randall, D., Brett, J. (eds.). Fish Physiology. Volume VIII; Bioenergetics and Growth. Academic Press, New York, USA. [ Links ]
Robinson EH, LaBomascus D, Brown PB, Linton TL (1987) Dietary calcium and phosphorus requirements of Oreochromis aureus reared in calcium-free water. Aquaculture 64: 267-276. [ Links ]
Robinson EH, Rawles SD, Yette HE, Greene LW (1984) An estimate of the dietary calcium requirement of fingerling Tilapia aurea reared in calcium-free water. Aquaculture 41: 389-393. [ Links ]
Roy PK, La l l, S.P (2003) Dietary phosphorus requirement of juvenile haddock (Melanogrammus aeglefinus L.). Aquaculture 221: 451-468. [ Links ]
Samocha TM, Davis DA, Saoud IP, DeBault K (2004) Substitution of fish meal by co-extruded soybean poul-try by-product meal in practical diets for the Pacific white shrimp, Litopenaeus vannamei. Aquaculture 231: 197-203. [ Links ]
Saoud IP, Rodgers LJ, Davis DA, Rouse DB (2008) Replacement of fish meal with poultry by-product meal in practical diets for redclaw crayfish (Cherax quadricarinatus). Aquaculture Nutrition. 14: 139-142. [ Links ]
Shearer KD (2000) Experimental design, statistical analysis and modeling of dietary nutrient requirement studies for fish: a critical review. Aquaculture Nutrition 6: 91-102. [ Links ]
Tacon AGJ (1993) Feed ingredients for warmwater fish: fish meal and other processed feedstuffs. FAO Fisheries Circular, no. 856, FAO, Rome. 64 pp. [ Links ]
Wilkinson S (2008) Diet formulator program (Excel). CSIRO Marine & Atmospheric Research. In: http://www.enaca.org/modules/wfdownloads/singlefile.php?cid=111&lid=952&utm [ Links ]
Zar JH (1984) Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, NY, USA, 718 pp. [ Links ]

