










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkUniversidad y ciencia
versão impressa ISSN 0186-2979
Universidad y ciencia vol.26 no.1 Villahermosa Abr. 2010
Artículos
Estructura y diversidad de la vegetación arbórea del Parque Estatal Agua Blanca, Macuspana, Tabasco
Structure and diversity of arboreal vegetation in the Parque Estatal Agua Blanca, Macuspana, Tabasco
VM Zarco–Espinosa1*, Jl Valdez–Hernández1, G Ángeles–Pérez1, O Castillo–Acosta2
1 Programa Forestal Colegio de Postgraduados km 36.5 Carretera México – Texcoco. Montecillo 56230 Edo. de México, México. (VMZE)(JIVH)(GAP). *Correo electrónico: vmzarcoe@colpos.mx
2 División Académica de Ciencias Biológicas, UJAT. (OCA)
Artículo recibido: 20 de junio de 2007
Aceptado: 5 de marzo de 2010
RESUMEN
Se analizó la estructura y diversidad de especies arbóreas en cuatro unidades de muestreo (UM) de 50 x 50 m (0.25 ha) cada una establecidas al azar en la porción centro–oeste del Parque Estatal Agua Blanca (PEAB), Macuspana, Tabasco. Se registraron variables dasométricas de todos los individuos con diámetro a la altura del pecho > 1 cm, determinando su estratificación vertical y distribución horizontal, así como el cálculo de índices de importancia estructural y de diversidad. La vegetación presentó dos estratos bien definidos: inferior (< 9 m) y superior (> 9 m), mientras que la distribución horizontal de los individuos fue según el índice de Morisita (Ιδ), generalmente aleatoria. Se identificaron 71 especies pertenecientes a 57 géneros y 40 familias; la familia Meliaceae fue la más rica en especies (14), seguida de Fabaceae (8) y Moraceae (7). El género Chamaedorea (Arecaceae), al igual que las especies Rinorea guatemalensis (Violaceae) y Astrocaryum mexicanum (Arecaceae), tuvieron los valores más altos de importancia estructural. De acuerdo con los índices de Margalef (Dα) y Shannon (Η'), la riqueza y diversidad de especies fue significativamente (p < 0.05) menor en la UM4 que en las UM1, UM2 y UM3, las cuales fueron estadísticamente iguales entre sí. En comparación con otras selvas de México, el PEAB tuvo valores intermedios en número de árboles por hectárea, bajos en riqueza de especies e intermedios en diversidad α de Fisher.
Palabras clave: Estratificación vertical, distribución horizontal, índices estructurales, riqueza de especies.
ABSTRACT
The diversity and structure were analysed for the species of trees in four 50 x 50 m (0.25 ha) sampling units (SU) randomly established in the central–western area of the "Parque Estatal Agua Blanca" (PEAB), Macuspana, Tabasco. Dasometric variables of all individuals with a diameter at breast height > 1 cm were registered, including the vertical stratification and horizontal distribution. The structural importance and diversity indices were calculated. The vegetation presented two well defined strata: lower (< 9 m) and higher (> 9 m), whereas the horizontal distribution of the specimens was generally random according to the Morisita index (Ισ). A total of 71 species were identified, belonging to 57 genera and 40 families. The Meliaceae family was the richest in species (14), followed by Fabaceae (8) and Moraceae (7). The genus Chamaedorea (Arecaceae) and the species Rinorea guatemalensis (Violaceae) and Astrocaryum mexicanum (Arecaceae) had the greatest values of structural importance. In accordance with the Margalef (Dα) and Shannon (H') indices, the richness and diversity of species were significantly (p < 0.05) lower in SU4 than in SU1, SU2 and SU3, which were statistically similar. Compared with other tropical rainforests in Mexico, the PEAB had intermediate values with respect to the number of trees per hectare, low values for species richness and intermediate values for Fisher's α diversity.
Key words: Vertical stratification, horizontal distribution, structural indices, species richness.
INTRODUCCIÓN
Latinoamérica es el área con mayor riqueza de especies en el mundo con cerca de 120 mil especies de plantas con flores (Toledo 1988) pero la que presenta a su vez el mayor grado de destrucción de sus ecosistemas naturales: 60–65% de la deforestación total mundial (Anónimo 2006a; Anónimo 2007). De manera similar, México se encuentra junto con Brasil, Colombia e Indonesia en los primeros lugares de riqueza de especies en el planeta (Anónimo 1998; Ceballos et al. 2005), la cual existe principalmente en sus bosques tropicales (Cruz–Lara et al. 2004; Martínez–Ramos & García–Orth 2007); sin embargo, entre 1976 y 2000 la tasa de pérdida de estos bosques fue 0.76% por año (Mas et al. 2002).
En el estado de Tabasco se han reportado tasas de disminución anual en cobertura de selvas de 6.1 % (selva baja) a 34.9% (selva mediana) en Pantanos de Centla (Guerra–Martínez & Ochoa–Gaona 2006) y de 4.1 a 11.6% (selva alta) en Sierra Madrigal (Salazar–Conde et al. 2004); lo cual ha provocado que para el período 2002–2005 se reporte solamente 3.4 % de cubierta de selvas del total de la superficie estatal (Anónimo 2008). No obstante este pequeño porcentaje, los remanentes de selvas en Tabasco albergan una alta biodiversidad: alrededor de 2500 especies de plantas (Pérez et al. 2005) y representan una fuente importante de bienes (p.ej. madera, medicinas) y servicios ambientales (p.ej. recarga hidrológica).
En reconocimiento a estos valores y con la finalidad de contribuir a su conservación, se han decretado áreas naturales protegidas (ANP) de carácter federal (Reserva de la Biosfera Pantanos de Centla; Anónimo 2006b) y estatal (p.ej. Parque Estatal de la Sierra) de las cuales forma parte el Parque Estatal Agua Blanca (Vargas–Márquez 2002).
En este Parque se han llevado a cabo trabajos relacionados con el medio físico, particularmente geomorfología y edafología (Castillo & Zavala 1996), propuestas de uso (Cruz 1999) y ordenamiento (Zequeiros 2006); así como con el entorno biológico, principalmente composición florística (Angel 2002), inventarios de murciélagos (Castro–Luna 1999) y anfibios (Barragán 2005). Sin embargo, faltan investigaciones que evalúen la distribución, diversidad y dominancia relativa de árboles. La obtención de esta información básica contribuirá a un mejor entendimiento de los patrones espaciales y de abundancia de la vegetación.
Por lo anterior, el presente estudio se desarrolló en el Parque Estatal Agua Blanca, municipio de Macuspana, estado de Tabasco, con la finalidad de describir la estructura vertical y horizontal de la vegetación arbórea, estimar valores de importancia estructural, así como determinar la composición florística, riqueza y diversidad de especies arbóreas.
Las hipótesis planteadas fueron: a) los individuos exhiben una distribución horizontal aleatoria en las unidades de muestreo y b) no existen diferencias en riqueza y diversidad de especies entre unidades de muestreo.
MATERIALES Y MÉTODOS
Área de estudio
El Parque Estatal Agua Blanca (PEAB) fue decretado el 19 de diciembre de 1987 en el Periódico Oficial del Gobierno del estado de Tabasco (Vargas–Márquez 2002). Se ubica en la porción Oeste y Noreste de la región Sierra de Tabasco, en el municipio de Macuspana (Figura 1). El Parque cuenta con una extensión territorial de 2,025 ha y se localiza entre los paralelos 17° 35" y 17° 38" de latitud Norte y 92° 25" y 92° 29" de longitud Oeste, entre los 100 y 200 m s. n. m. (Anónimo 2003).
Este Parque forma parte de la provincia fisiográfica Sierra Norte de Chiapas, donde predomina la roca caliza del Cretácico Superior, de naturaleza cárstica, caracterizada por la disolución de esta roca que origina un sistema de ríos subterráneos y cavernas (p. ej. gruta del cerro) formando el arroyo Agua Blanca. Los suelos, desarrollados in situ, son derivados de la desintegración de rocas calizas y se clasifican como rendzinas; son delgados y con presencia de un horizonte "A" de color oscuro, rico en nutrientes, el cual está localizado arriba de un horizonte "C" o roca madre (Castillo & Zavala 1996).
El clima es cálido húmedo con lluvias todo el año Af(m) w" (i)g, la temperatura promedio anual fluctúa entre los 23 y 26 °C, mientras la precipitación pluvial anual promedio va de 2100 a 3200 mm (García 1988). La vegetación corresponde a 2 000 ha de selva mediana perennifolia (Castillo & Zavala 1996).
Procedimiento de muestreo y medición de variables
Se establecieron cuatro unidades de muestreo (UM) de 50 x 50 m (0.25 ha) cada una. Estas se trazaron con ayuda de una brújula y la inclinación de las pendientes se corrigió con un clisímetro. Cada UM fue dividida en 25 cuadros de 10 x 10 m (100 m2) cada uno, que fueron señalados con estacas de madera. En cada cuadro se localizaron mediante coordenadas cartesianas (x, y) todos los individuos de especies arbóreas con diámetro a la altura del pecho (DAP) > 1 cm, de acuerdo con lo sugerido por Hubbell (1979), Hubbell & Foster (1983) y Dallmeier (1992) para bosques tropicales. A cada individuo se le asignó un número y fueron marcados con etiquetas de aluminio, las cuales fueron sujetadas evitando dañar los árboles.
Se censaron todos los árboles enraizados dentro de las UM y que presentaron un tronco definido hasta una altura de 1.30 m. Para cada individuo se registró su identidad taxonómica, diámetro a la altura del pecho con cinta diamétrica; la altura total y de fuste limpio (hasta la primera ramificación) se obtuvieron con estadal extendible (individuos < 8 m) y clisímetro (árboles > 8 m). También se midió el diámetro máximo y mínimo de copa en cada individuo, considerando las proyecciones de la misma sobre el suelo (Dawkins 1958).
En forma simultánea al censo, se realizó la recolecta por triplicado de ejemplares botánicos de cada especie, los cuales fueron procesados, herborizados (Lot & Chiang 1986) e identificados taxonómicamente en los herbarios CHAPA del Colegio de Postgraduados campus Tabasco y UJAT de la División Académica de Ciencias Biológicas de la Universidad Juárez Autónoma de Tabasco, quedando depositados en éste último herbario.
Cálculo de atributos dasométricos e índices estructurales
Con objeto de describir la estructura vertical de las UM se elaboraron histogramas de frecuencia por categoría de altura, mientras que para evaluar la estructura horizontal se aplicó el índice de Morisita (Morisita 1959):

Donde:
Ιδ = Índice de distribución espacial, q = Número de cuadros, ni = Número de individuos en el i–ésimo cuadro, N = Número total de individuos en todos los q cuadros.
Valores de Ιδ menores de uno, iguales a uno y mayores a uno indican una distribución regular o uniforme, distribución aleatoria y distribución agregada, respectivamente (Morisita 1959).
La información del DAP de los individuos censados se organizó por categoría diamétrica de 10 cm de amplitud: 1.0 a 11.0, 11.1 a 21.0, y así sucesivamente. Este análisis se realizó conjuntando los valores de las cuatro UM (1 ha).
Se utilizaron dos índices de valoración estructural: a) índice de valor de importancia (IVI) y b) índice de valor forestal (IVF). Estos índices fueron determinados para cada UM (0.25 ha) de manera independiente, así como para las cuatro UM en conjunto (1 ha).
Indice de Valor de Importancia (IVI). Fue desarrollado por Curtis & McIntosh (1951) y aplicado por Pool etal. (1977), Cox(1981), Cintrón & Schaeffer–Novelli (1983) y Corella et al. (2001). Es un índice sintético estructural, desarrollado principalmente para jerarquizar la dominancia de cada especie en rodales mezclados y se calculó de la siguiente manera:

La dominancia (estimador de biomasa: área basal, cobertura) relativa se obtuvo de la siguiente manera:

Donde:

El área basal (AB) de los árboles se obtuvo con la fórmula siguiente:
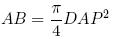
La densidad relativa se calculó de la siguiente manera:

Donde:

La frecuencia relativa se calculó de la siguiente manera:

Donde:

Indice de Valor Forestal (IVF). Se aplicó con el propósito de evaluar la estructura bidimensional de la vegetación arbórea en zonas cálido–húmedas de México considerando tres medidas: la primera al nivel del estrato inferior en el plano horizontal (diámetro a la altura del pecho), la segunda que incluye los estratos inferior y superior en el plano vertical (altura), y la tercera al nivel del estrato superior en el plano horizontal (cobertura); se calculó como sigue (Corella et al. 2001):

El diámetro relativo se obtuvo mediante la fórmula:

Donde:

La altura relativa se obtuvo mediante la fórmula:

Donde:
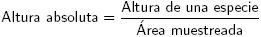
La cobertura relativa se obtuvo mediante la fórmula:

Donde:

La cobertura se estimó mediante la fórmula para el área de una elipse:

Donde:
a = semieje mayor
b = semieje menor
Medidas de la diversidad de especies
A partir de la información obtenida se elaboraron curvas de especies–área para cada UM, las cuales consistieron en graficar el número de especies vegetales encontradas para una superficie de muestreo determinada (Greig–Smith 1983). La riqueza de especies (Dα) se calculó mediante el Índice de Margalef (1977):

Donde:
S = número de especies
N = número total de individuos
A mayor valor de D mayor riqueza de especies.
Con el propósito de conocer que tan homogéneas o heterogéneas fueron las UM, se calcularon los siguientes índices de diversidad (Magurran 1988; Krebs 1989):
Shannon–Wiener (Η'). Mide el grado promedio de incertidumbre para predecir la especie a la que pertenece un individuo tomado al azar dentro de las UM.

Donde:
S = número de especies
Pi = proporción de individuos de la especie i
A mayor valor de Η' mayor diversidad de especies.
Simpson (S). Mide la probabilidad de que dos individuos seleccionados al azar en las UM sean de la misma especie.

Donde:
ni = número de individuos en la iésima especie
N = número total de individuos
A mayor valor de S menor dominancia de una (o de un grupo) de especie(s).
Equidad (E). La equidad se calculó con la siguiente ecuación:

Donde:
Η' = índice de Shannon–Wiener
S = número total de especies
Valores cercanos a 1 representan condiciones hacia especies igualmente abundantes y aquellos cercanos a 0 la dominancia de una sola especie. Para determinar la existencia de diferencias significativas (Tukey, p < 0.05) en la diversidad de especies arbóreas entre las UM, se utilizó el paquete estadístico SAS (Schlotzhauer & Littell 2000).
Diversidad α de Fisher está dado por (Fisher et al. 1943):

Donde:
S = número de especies registradas en la muestra
N = total de individuos en la muestra α = índice de diversidad
Este índice se empleó para comparar cuantitativamente este estudio con otros, ya que sólo considera el número de especies (S) y el total de individuos (N) en las muestras estudiadas (Leigh 1999; Berry 2002); además, no depende del tamaño del área muestreada como los índices de Shannon y Simpson. El cálculo de estos tres índices se realizó usando el programa EstimateS (Colwell 2005).
Con la finalidad de conocer la semejanza florística entre las UM, se utilizó el coeficiente de Sorensen (IS) que se basa en la relación presencia–ausencia de especies entre dos sistemas (Stiling 1999):

Donde:
A = número de especies encontradas en la comunidad A
B = número de especies encontradas en la comunidad B
C = número de especies comunes en ambas comunidades
Este coeficiente presenta valores que van de 0 a 1, significando la menor y mayor semejanza, respectivamente.
RESULTADOS
Estratificación vertical
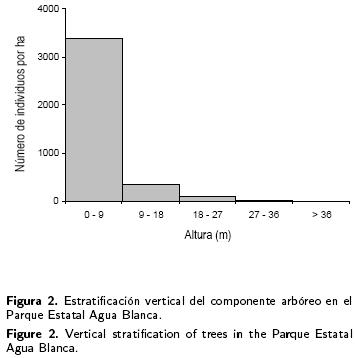
La altura del componente arbóreo en el PEAB alcanzó los 27 m, con algunos individuos mayores a 35 m; se lograron diferenciar dos estratos: inferior y superior (Figura 2). El estrato inferior estuvo constituido por categorías de altura menores a 9 m (86.5% del total de individuos), mientras que el estrato superior comprendió categorías de altura mayores a 9 m (13.5 % del total).
Distribución horizontal
Los valores en DAP variaron de 1 cm a 2.32 m, con una media de 5.7 cm; de 3789 individuos censados, 3722 pertenecieron a las primeras cuatro categorías diamétricas, con el 76 % incluido en la categoría de 1 a 11 cm DAP (Figura 3).

De acuerdo con el Índice de Morisita (Ιδ), en las unidades de muestreo se observó en general, una distribución aleatoria (Ιδ cercanos a 1) de los individuos (Figura 4) mostrando un patrón agregado en el estrato inferior de la UM1 (Figura 4a, Ιδ = 1.27; Figura 4b, Ιδ = 1.36) y en el estrato superior de la UM4 (Figura 4c, Ιδ= 1.28).
Índices estructurales
Las especies con altos IVI presentes en las cuatro UM fueron Rinorea guatemalensis, Astrocaryum mexicanum, Quararibea funebris y Chamaedorea pinnatifrons; los valores de frecuencia y densidad relativas determinaron el IVI de R. guatemalensis y los de área basal el de Q. funebris (Tabla 1).
Las UM 2, 3 y 4 comparten Pseudolmedia oxyphyllaria y Psychotria chiapensis con IVI elevados, mientras que Parmentiera aculeata también con alto IVI se comparte en las UM 1, 2 y 4 (Tabla 1). Otras especies de importancia estructural fueron Chamaedorea tepejilote, Bursera simaruba y Louteridium mexicanum (Tabla 1); Guarea glabra y Poulsenia armata (UM2), Dialium guianense y Guatteria anomala (UM3), así como Ceiba pentandra (UM4).
Tomando en cuenta las diez especies con mayor IVF en las cuatro UM, se confirmó la importancia estructural de Rinorea guatemalensis, Astrocaryum mexicanum, Quararibea funebris y Chamaedorea pinnatifrons (Tabla 2).
En contraste, especies con IVF elevados que no tuvieron altos IVI fueron Trophis racemosa en la UM1, Dialium guianense en la UM2, Cryosophila argentea en la UM3 y Guarea chichon en la UM4 (Tabla 2). En este estudio se registraron 71 especies de árboles con DAP > 1 cm pertenecientes a 40 familias.
Considerando las cuatro UM (1 ha), la familia Meliaceae con 14 especies fue la más rica sobresaliendo el género Guarea con tres especies determinadas y tres morfoespecies, de ellas Guarea glabra aún cuando presentó bajas densidades (26 ind ha–1) su dominancia por área basal fue alta (28.5 m2 ha–1). La familia Fabaceae fue la segunda en importancia con ocho especies, sobresaliendo Dialium guianense (25 ind ha–1, 29.4 m2 ha–1), y la familia Moraceae con siete especies fue la tercera en importancia, destacando Poulsenia armata (27 ind ha–1 , 16.1 m2 ha–1). Los géneros Astrocaryum y Chamaedorea (Arecaceae) tuvieron al igual que Rinorea guatemalensis (Violaceae) una alta importancia estructural.
Diversidad de especies
De acuerdo con los índices de Margalef y Shannon la riqueza y diversidad de especies fue significativamente (p < 0.05) menor en la UM4 que en las otras tres UM, las cuales fueron estadísticamente iguales entre sí (Tabla 3); en contraste, los índices de Simpson y α de Fisher no mostraron diferencias significativas entre las UM (Tabla 3).

Semejanza florística
Los valores más altos se obtuvieron entre la UM1 y UM2: 55 a 71 % de semejanza y 29 a 45 % de especies distintas (Tabla 4). En general, los valores más bajos se presentaron entre la UM4 con relación a las otras UM, lo cual se corrobora al observar las curvas especies–área (Figura 5).

DISCUSIÓN
Estratificación vertical
La distribución de los individuos por su altura en el área muestreada permitió definir claramente dos estratos verticales: inferior (< 9 m) y superior (> 9 m); sin embargo, para esta misma área otros autores han establecido tres estratos a intervalos de 10 m cada uno (Castillo & Zavala 1996; Ángel 2002), posiblemente debido a que en estos trabajos el limite inferior de DAP fue de 2.5 cm, mientras que en el presente estudio fue de 1.0 cm, lo que incrementó el número de árboles en categorías inferiores y permitió identificar una clara discontinuidad en la distribución de alturas: 87% de individuos en el estrato inferior y 13 % en el superior. Esto coincide con lo obtenido por Godínez–Ibarra & López–Mata (2002) en una selva mediana subperennifolia de Veracruz, donde la distribución de individuos por su altura mostró valores cercanos a 88% en el estrato inferior y a 12 % en los superiores.
El análisis de distribución realizado por categoría diamétrica en las cuatro UM (1 ha) mostró al 76 % de los individuos en la categoría de 1 a 11 cm DAP; lo que corresponde al patrón de distribución diamétrica tipo I señalado por Bongers et al. (1988), donde la gran mayoría de los individuos se encuentran en categorías pequeñas y existe una continua incorporación hacia las siguientes categorías.
Los resultados obtenidos en la distribución de alturas y diámetros, así como de composición florística, muestran que el 80% de árboles muestreados en la selva mediana perennifolia del PEAB pertenecieron al estrato inferior, donde Rinorea guatemalensis junto con la palma Astrocaryum mexicanum constituyeron parte importante, similar a lo mencionado para el estrato inferior de otras selvas medianas y altas perennifolias de México (Meave del Castillo 1990).
Especies tolerantes a la sombra como A. mexicanum, Dialium guianense, Guarea glabra y Pseudolmedia oxyphyllaria, distribuidas en lugares con escasa perturbación, estuvieron bien representadas en el área de estudio, superando en valor de importancia a Heliocarpus donnell–smithii, Bursera simaruba y Dendropanax arboreus que, de acuerdo con Pennington & Sarukhán (1998) son especies de vegetación secundaria como Quararibea funebris la cual se presentó en ambos estratos (inferior y superior).
Distribución horizontal
Se rechaza la hipótesis de distribución aleatoria de individuos en las unidades de muestreo, puesto que en dos de ellas (UM1 y UM4) se encontró un patrón agregado. Dos especies influyeron principalmente en dicho resultado: R. guatemalensis (Ιδ= 2.24) para el estrato inferior de la UM1 y Q. funebris (Ιδ= 1.13) para el estrato superior de la UM4, ambas con altos valores de importancia estructural y una distribución agregada.
Esta distribución agregada puede ser atribuida a condiciones físicas en las UM1 y UM4, particularmente diferencias notables en altura del microre–lieve (5–10 m) que generaron una superficie heterogénea para el establecimiento de individuos; así como a características biológicas de R. guatemalensis y Q. funebris, clasificada la primera como una especie no secundaria que existe en bosque primario pero demandante de luz solar durante su primera etapa de crecimiento (Arroyo–Rodríguez & Mandujano 2006), en tanto que la segunda es reconocida como una especie exclusiva de vegetación madura (Levy–Tacher et al. 2002) cuyas semillas son dispersadas por animales (Benítez–Malvido & Lemus–Albor 2005).
Importancia estructural
La vegetación del PEAB mostró similitudes con lo encontrado por Bongers et al. (1988) en Los Tuxtlas, Veracruz, donde A. mexicanum y P. oxyphyllaria tuvieron valores altos de dominancia relativa, y con lo reportado por Meave del Castillo (1990) en Bonampak, Chiapas, donde Psychotria chiapensis tuvo densidades importantes. En cambio, para la selva alta de Yaxchilán, Chiapas, Valle (2000) cita entre las especies más importantes a Chamaedorea tepejilote, Protium copal, Brosimum alicastrum y Ceiba pentandra.
El valor del índice de diversidad de Shannon obtenido en el presente trabajo fue de 2.5, el cual se puede considerar bajo al compararse con los de otras selvas en México; 4.7 en Chamela, Jalisco (Lott et al. 1987), 4.6 en Los Tuxtlas, Veracruz (Bongers et al. 1988), 3.3 en Santa Gertrudis, Veracruz (Godínez–Ibarra & López–Mata 2002), 3.1 en Calakmul, Campeche (Díaz–Gallegos et al. 2001), 3.5 en San Miguel, Veracruz (Villavicencio–Enríquez & Valdez–Hernández 2003). Esto sugiere, que la vegetación en el área del presente estudio puede encontrarse en diferentes etapas de sucesión ecológica. Al confrontar los valores del índice de diversidad α de Fisher para otras selvas de México, se observa que el obtenido en el PEAB fue intermedio (Tabla 5).
Diversidad de especies
Se rechaza la hipótesis de igual riqueza y diversidad de especies arbóreas entre unidades de muestreo, debido a que una de ellas (UM4) fue significativamente diferente de las otras. Este resultado se confirma al observar tanto las curvas especies–área (Figura 5), donde la UM4 se ubica por debajo de las otras UM, como al comparar los valores del coeficiente de Sorensen (Tabla 4), el cual indica menor semejanza florística (i.e. menor cantidad de especies compartidas) de la UM4 con relación a las otras UM.
Esta menor diversidad en especies puede ser debida, según Clark (2002), a las propiedades del terreno. Al respecto, la UM4 tuvo una mayor inclinación de la pendiente (36–70%), exposición sur y menor fertilidad del suelo, en comparación con las otras UM que presentaron menor inclinación de la pendiente (0–57 %), exposiciones norte y noroeste, así como mayor fertilidad del suelo. Asimismo, y de acuerdo con García–Montiel (2002), debe tomarse en cuenta el efecto de las perturbaciones humanas en la composición y estructura de los bosques tropicales. En el área de estudio dichas perturbaciones se refieren principalmente a la realización de actividades con fines agropecuarios (p.ej. uso del fuego) y extracción ilegal de madera (p.ej. árboles grandes de especies valiosas).
La UM4 mostró características que indican un mayor nivel de perturbación: mayor densidad de individuos (4420 ha–1 ) y menores valores promedio tanto en altura de árboles (5.2 m) como en cobertura de sus copas (4.9 m2). En comparación, las otras UM tuvieron menor densidad de individuos (24804184 ha–1) y mayores valores promedio tanto en altura de árboles (5.7–6.6 m) como en cobertura de sus copas (8.9–11.9 m2). Además, la UM4 tuvo especies consideradas de vegetación secundaria como Parmentiera aculeata que alcanzó mayores valores de importancia y forestal en comparación con las otras UM.
Sin embargo, y tomando en cuenta los hallazgos de Arroyo–Rodríguez & Mandujano (2006) así como lo expresado por Ochoa–Gaona et al. (2007), las selvas en etapas de sucesión temprana pueden actuar como reservas de especies propias de etapas más tardías. Esto se aprecia en la UM4 donde P. oxyphyllaria, Q. funebris, A. mexicanum y D. guianense, reconocidas como elementos de vegetación madura (Levy–Tacher et al. 2002; Benítez–Malvido & Lemus–Albor 2005) o de bosque primario (Arroyo–Rodríguez & Mandujano 2006; Ochoa–Gaona et al. 2007), estuvieron entre las primeras nueve especies con mayores valores de importancia y forestal.
AGRADECIMIENTOS
El presente estudio se llevó a cabo gracias al financiamiento del fondo sectorial SEMARNAT–CONACyT a través del proyecto 2002–001–0761: "Diagnostico de la selva alta perennifolia del estado de Tabasco". El primer autor agradece al CONACyT la beca otorgada para el desarrollo de sus estudios de Maestría en el Colegio de Postgraduados (COL–POS), campus Montecillo. Al Dr. Joel Zavala Cruz, Profesor Investigador del COLPOS campus Tabasco, por el apoyo brindado en el desarrollo de este trabajo. Un agradecimiento especial a la entusiasta colaboración del grupo de estudiantes de la M.C. Ofelia Castillo Acosta de la División Académica de Ciencias Biológicas (DACB) de la Universidad Juárez Autónoma de Tabasco (UJAT).
LITERATURA CITADA
Ángel ARL (2002) Estructura y composición florística del Parque Estatal Agua Blanca, Macuspana, Tabasco, México. Tesis Licenciatura UJAT DACB. 82 p. [ Links ]
Anónimo (1998) La diversidad biológica de México: Estudio de País, 1998. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. 341 p. [ Links ]
Anónimo (2003) Carta Geográfica Estatal 2003. Secretaria de Comunicaciones, Asentamientos y Obras Públicas de Tabasco. [ Links ]
Anónimo (2006a) Tendencias y perspectivas del sector forestal en América Latina y el Caribe. Estudio FAO: Montes 148. Roma. 178 p. [ Links ]
Anónimo (2006b) VI Aniversario CONANP. Comisión Nacional de Areas Naturales Protegidas. SEMARNAT. México. [ Links ]
Anónimo (2007) Situación de los bosques del mundo. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Roma. 143 p. [ Links ]
Anónimo (2008) Anuario estadístico de Tabasco, 2008. Gobierno del estado de Tabasco. Instituto Nacional de Estadística, Geografía e Informática. México. [ Links ]
Arroyo–Rodríguez V, Mandujano S (2006) The importance of tropical rain forest fragments to the conservation of plant species diversity in Los Tuxtlas, México. Biodiversity and Conservation 15: 4159–4179. [ Links ]
Barragán R (2005) Anfibios del Parque Estatal de Agua Blanca. Tesis de Maestría. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco. [ Links ]
Benítez–Malvido J, Lemus–Albor A (2005) The seedling community of tropical rainforest edges and its interaction with herbivores and pathogens. Biotropica 37(2): 301–313. [ Links ]
Berry PE (2002) Diversidad y endemismo en los bosques neotropicales de bajura. pp. 83–96. En: Guariguata MR, Kattan GH (eds) Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional (EULAC–GTZ). Cartago, Costa Rica. [ Links ]
Bongers F, Popma J, Meave del Castillo J, Carabias J (1988) Structure and floristic composition of the lowland rain forest of Los Tuxtlas, Mexico. Vegetatio 74: 55–80. [ Links ]
Castillo AO, Zavala CJ (1996) Fisiografía, recursos vegetales y alternativas de manejo en el Parque Estatal Agua Blanca, Tabasco, Universidad y Ciencia 12(23): 63–70. [ Links ]
Castro–Luna A (1999) Composición y abundancia de la comunidad de murciélagos (Mammalia: Chiroptera) del Parque Estatal Agua Blanca, Macuspana, Tabasco. Universidad y Ciencia 15 (29): 57–68. [ Links ]
Ceballos G, Arroyo–Cabrales J, Medellín R.A., Medrano–González L, Oliva G (2005) Diversidad y conservación de los mamíferos de México. pp. 21–66. En: Ceballos G, Oliva G (coords) Los Mamíferos Silvestres de México. CONABIO. FCE. México. [ Links ]
Cintrón G, Schaeffer–Novelli Y (1983) Introducción a la ecología del manglar UNESCO. Montevideo, Uruguay 109 p. [ Links ]
Clark DB (2002) Los factores edáficos y la distribución de las plantas. pp. 193–221. En: Guariguata MR, Kattan GH (eds) Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional (EULAC–GTZ). Cartago, Costa Rica. [ Links ]
Colwell RK (2005) EstimateS: Statistical estimation of species richness and shared species from samples: Versión 7.5. Persistent URL <Purl.oclc.org/estimates> [ Links ].
Corella JF, Valdez HJI, Cetina AVM, González CFV, Trinidad SA, Aguirre RJR (2001) Estructura forestal de un bosque de mangles en el noreste del estado de Tabasco, México. Ciencia Forestal en México 26(90): 73–102. [ Links ]
Cox WG (1981) Laboratory manual of general ecology. William C. Brown Co. Publishers. Iowa, USA. 230 p. [ Links ]
Cruz HP (1999) Ordenamiento ecológico del Parque Estatal Agua Blanca, Tabasco. Tesis de Licenciatura. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco. [ Links ]
Cruz–Lara LE, Lorenzo C, Soto L, Naranjo E, Ramírez–Marcial N (2004) Diversidad de mamíferos en cafetales y selva mediana de las cañadas de la Selva Lacandona, Chiapas, México. Acta Zoológica Mexicana (n.s.) 20(1): 63–81. [ Links ]
Curtis JT, McIntosh RP (1951) An upland forest continuum in the pariré–forest border region of Wisconsin. Ecology 32: 476–496. [ Links ]
Dallmeier F (1992) Long–term monitoring of biological diversity in tropical forest areas: methods for establishment inventory of permanent plot. MAB Digest 11 UNESCO– Pris. 72 p. [ Links ]
Dawkins HC (1958). The management of natural tropical high–forest with special reference to Uganda. Institute Paper No 34. Imperial Forestry Institute, Oxford. 155 p. [ Links ]
Díaz–Gallegos JR, García–Gil G, Castillo O, March EI (2001) Uso del suelo y transformación de selvas en un ejido de la Reserva de la Biosfera Calakmul, Campeche, México. Investigaciones Geográficas 44: 39–53. [ Links ]
Fisher RA, Cobet AS, Williams CB (1943) The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology 12: 42–58. [ Links ]
García E (1988) Modificaciones al sistema de clasificación climática de Kôppen (para adaptarlo a las condiciones de la República Mexicana) Offset Larios, México, D.F. 165 p. [ Links ]
García–Montiel DC (2002) El legado de la actividad humana en los bosques neotropicales contemporáneos. pp. 97–116. En: Guariguata MR, Kattan GH (eds) Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional (EULAC–GTZ). Cartago, Costa Rica. [ Links ]
Godínez–Ibarra O, López–Mata L (2002) Estructura, composición, riqueza y diversidad de árboles en tres muestras de selva mediana subperennifolia. Anales del Instituto de Biología. Universidad Nacional Autónoma de México. Serie Botánica 73(2): 283–314. [ Links ]
Greig–Smith P (1983) Quantitative plant ecology. 3er Edition. University of California Press. Berkeley, CA. 347 p. [ Links ]
Guerra–Martínez V, Ochoa–Gaona S (2006) Evaluación espacio–temporal de la vegetación y uso del suelo en la Reserva de la Biosfera Pantanos de Centla, Tabasco (1990–2000). Investigaciones Geográficas 59:7–25. [ Links ]
Hubbell SP (1979) Tree dispersion, abundance, and diversity in a tropical dry forest. Science 203:1299–1309. [ Links ]
Hubbell SP, Foster RB (1983) Diversity of canopy trees in a neotropical forest and implications for conservation. pp. 25–41. En: Sutton SL, Whitmore TC, Chadwick AC (eds) Tropical Rain Forest: Ecology and Management. Blackwell Scientific, Oxford. [ Links ]
Krebs CJ (1989) Species diversity measures. pp. 328–370. En: Krebs CJ (ed) Ecological Methodology. Uharper Collins Publishers, Inc. [ Links ]
Leigh GE Jr (1999) Tropical diversity. pp. 179– 210 En: Leigh GE Jr (ed) Tropical Forest Ecology: a view from Barro Colorado Island. Oxford University Press. New York. [ Links ]
Levy–Tacher SI, Aguirre–Rivera JR, Martínez–Romero MM, Durán–Fernández A (2002) Caracterización del uso tradicional de la flora espontánea en la comunidad lacandona de Lacanhá, Chiapas, México. Interciencia 27(10): 512–520. [ Links ]
Lot A, Chiang F (1986) Manual de Herbario. Administración y manejo de colecciones, técnicas de recolección y preparación de ejemplares botánicos. Consejo Nacional de Ciencia y Tecnología. México. 142 p. [ Links ]
Lott EJ, Bullock SH, Sanchez–Magallanes JA (1987) Floristic diversity and structure of upland and arroyo forests of coastal Jalisco. Biotropica 19: 228–235. [ Links ]
Magurran AE (1988) Ecological Diversity and its Measurement. Princeton University Press, Princeton. N. J. 179 p. [ Links ]
Margalef R (1977) Ecología. Ediciones Omega. Barcelona 951 p. [ Links ]
Martínez–Ramos M, García–Orth X (2007) Sucesión ecológica y restauración de las selvas húmedas. Boletín de la Sociedad Botánica de México 80: 69–84. [ Links ]
Mas JF, Velázquez A, Díaz JR, Mayorga R, Alcántara C, Castro R, Fernández T (2002) Monitoreo de los cambios de cobertura en México. Memorias del II Seminario Latinoamericano de Geografía Física (Disco Compacto). 24–27 julio 2002. Maracaibo, Venezuela. [ Links ]
Meave del Castillo J (1990) Estructura y composición de la selva alta perennifolia de los alrededores de Bonampak. Colección Científica, serie Arqueología, Instituto Nacional de Antropología e Historia, México, D.F. 147 p. [ Links ]
Morisita M (1959) Measuring the dispersion and the analysis of distribution patterns. Memoires of the Faculty of Science, Kyushu University, Series E. Biology 2: 215–235. [ Links ]
Ochoa–Gaona S, Hernández–Vázquez F, De Jong BHJ, Gurri–García FD (2007) Pérdida de diversidad florística ante un gradiente de intensificación del sistema agrícola de roza–tumba–quema: un estudio de caso en la Selva Lacandona, Chiapas, México. Boletín de la Sociedad Botánica de México 81: 65–80. [ Links ]
Pennington TD, Sarukhán J. (1998) Árboles tropicales de México Fondo de Cultura Económica ONU–FAO UNAM. México 521 p. [ Links ]
Pérez LA, Sousa SM, Hanan AM, Chiang F, Tenorio P (2005) Vegetación terrestre. pp. 65–110. En: Bueno J, Alvarez F, Santiago S (eds) Biodiversidad del estado de Tabasco. Instituto de Biología, UNAM–CONABIO. México. [ Links ]
Pool DJ, Snedaker SC, Lugo AE (1977) Structure of mangrove forest in Florida, Puerto Rico, México and Costa Rica. Biotropica 9(3): 195–212. [ Links ]
Salazar–Conde EC, Zavala–Cruz J, Castillo–Acosta O, Cámara–Artigas R (2004) Evaluación espacial y temporal de la vegetación de la Sierra Madrigal, Tabasco, México (1973–2003). Investigaciones Geográficas 54:7–23. [ Links ]
Schlotzhauer DS, Littell CR (2000) SAS System for elementary statistical analysis. Second edition. University of Florida 123 p. [ Links ]
Stiling P (1999) Ecology; theories and applications. 3rd edition. Prentice Hall. New Jersey, USA. 840 p. [ Links ]
Toledo VM (1988) La diversidad biológica de Latinoamérica: un patrimonio amenazado. Ambiente y Desarrollo 4 (3): 13–24. [ Links ]
Valle JA (2000) Análisis estructural de una hectárea de selva alta perennifolia en el monumento natural Yaxchilán (Chiapas), México. Tesis. Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F. 99 p. [ Links ]
Vargas–Márquez F (2002) Areas naturales protegidas de México con decretos estatales. Volumen 2. SEMARNAT. Instituto Nacional de Ecología. CONANP. México. 1014 p. [ Links ]
Villavicencio–Enríquez I, Valdez–Hernández JI (2003) Análisis de la estructura arbórea del sistema agroforestal rusticano de café en San Miguel, Veracruz, México. Agrociencia 37(4): 413–423. [ Links ]
Zequeiros C (2006) Plan Maestro de ordenamiento ecológico del Parque Estatal de Agua Blanca. Tesis de Maestría. División Académica de Ciencias Económico Administrativas, Universidad Juárez Autónoma de Tabasco. [ Links ]

