










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkUniversidad y ciencia
versión impresa ISSN 0186-2979
Universidad y ciencia vol.24 no.1 Villahermosa abr. 2008
Artículos
Efecto de nauplios de Artemia franciscana enriquecidos sobre el crecimiento, supervivencia y resistencia al estrés de postlarvas (PL5-PL40) de Litopenaeus vannamei (BOONE, 1931).
Effect of enriched Artemia franciscana nauplii on growth, survival and stress resistance of Litopenaeus vannamei (Boone, 1931) postlarvae (PL5-PL40).
R Gelabert1, R Brito1, MG Gaxiola2, T Castro3, C Rosas2
1 DES Ciencias Naturales y Exactas, UNACAR. Av. 56 No 4 por. Av. Concordia, Ciudad de Carmen CP. 24180, Campeche, México. rgelabert@pampano.unacar.mx.
2 Unidad Multidisciplinaria de Docencia e Investigación, Facultad de Ciencias, UNAM.
3 Departamento el Hombre y su Ambiente. UAM Xochimilco.
Artículo recibido: 1 de noviembre de 2007
Aceptado: 12 de marzo de 2008
Resumen
Se estimó el crecimiento (incremento en peso seco), la supervivencia y la resistencia a un estrés salino en postlarvas de Litopenaeus vannamei alimentadas con nauplios enriquecidos de Artemia franciscana (Kellog, 1906). Se estudiaron los estadios de PL5 a PL10, de PL10 a PL30, y de PL20 a PL40. Las dietas consistieron en: 1) nauplios de A. franciscana enriquecida con una emulsión de lecitina de soya y aceite de hígado de bacalao (NA+L+AHB), 2) nauplios de A. franciscana enriquecida con una emulsión comercial (NA+IKA W3), y 3) nauplios de A. franciscana no enriquecida como control (NANE). Ninguna de las dietas influyó en el crecimiento de las postlarvas mayores (PL20-40) (p = 0.351). En las postlarvas menores (PL5_10) y de edad intermedia (PL10-30) se observó un crecimiento significativamente mayor (p < 0.05) cuando fueron alimentadas con ambas dietas de A. franciscana enriquecida. El incremento en peso seco de PL5-10 y PL10-30 no presentó diferencias estadísticamente significativas cuando se emplearon las dos dietas de nauplios de Artemia franciscana enriquecidos (p > 0.05). Ninguna dieta afectó la supervivencia ni la resistencia al estrés en los tres grupos de postlarvas analizados (p > 0.05). Se concluye que es más importante garantizar una apropiada alimentación de las postlarvas menores que reforzar la alimentación en las postlarvas más avanzadas.
Palabras clave: Artemia franciscana, bioencapsulación, Litopenaeus vannamei, postlarva, estrés, alimento vivo.
Abstract
Growth (dry weight increase), survival and resistance to saline stress were estimated for Litopenaeus vannamei postlarvae fed enriched Artemia franciscana (Kellog, 1906) nauplii. Stages PL5 to PL10, PL10 to PL30, and PL20 to PL40 were studied. Diets consisted of: 1) A. franciscana nauplii enriched with an emulsion of soybean lecithin and cod liver oil (NA+L+AHB), 2) A. franciscana nauplii enriched with a commercial emulsion (NA+IKA W3), and 3) non-enriched A. franciscana nauplii as control (NANE). No diet affected the growth of the bigger postlarvae (PL20-40) (p = 0.351), however significantly greater growths (p < 0.05) were observed in the younger (PL5-10) and intermediate age (PL10-30) postlarvae when fed the two diets with enriched A. franciscana nauplii. The dry weight increase of PL5-10 and PL10-30 was not statistically different when using the two diets with enriched A. franciscana nauplii (p > 0.05). No diet affected survival or the resistance to saline stress in the three age groups (p > 0.05). It is concluded that it is more important to insure a proper feeding of the early postlarvae than to reinforce the feeding of the more advanced stages.
Key words: Artemia franciscana, bioencapsulation, Litopenaeus vannamei, postlarva, stress, live feed.
INTRODUCCIÓN
Uno de los aspectos al que los acuacultores han prestado una considerable atención es el relacionado con la adecuada nutrición de los organismos cultivados. Diversos autores han estudiado el efecto o los requerimientos de los lípidos en la nutrición del camarón blanco Litopenaeus vannamei (Boone, 1931) y resaltan la importancia de los mismos como estimuladores del crecimiento, fuente de energía, constituyentes estructurales de las membranas celulares que favorecen los procesos de osmorregulación y la permeabilidad de las branquias, agente de transporte de sustancias liposolubles y precursores metabólicos, entre otros (Glencross & Smith 2001a; Glencross et al. 2001; Palacios et al. 2002; Palacios et al. 2004; Cheng & Hardy 2004; Martins et al. 2006).
La elaboración de dietas artificiales es otro de los aspectos que se ha abordado intensamente con vistas a reducir los costos de producción y garantizar una adecuada nutrición de los organismos. Una variante es la modificación nutricional del alimento vivo mediante las técnicas de bioencapsulación, que permiten la incorporación de elementos esenciales en la dieta, y en algunos casos, estimulan respuestas como el crecimiento, supervivencia, desarrollo y resistencia a condiciones estresantes comúnmente presentadas en las actividades de cultivo (Sakamoto et al. 1982; Clawson & Lovell 1992; Fernández-Reiriz et al. 1993; Rodríguez et al. 1996; Sorgeloos et al. 1998; Gara et al. 1998; Narciso et al. 1999; Han et al. 2000; Gómez-Gil et al. 2001). Algunos investigadores han estudiado el enriquecimiento del alimento vivo con emulsiones lipídicas cuyos componentes principales son aceites ricos en ácidos grasos polinsaturados de cadena larga extraídos de peces como el bacalao (Southgate & Lou 1995; Czesny et al. 1999; Smith et al. 2002) y lo recomiendan como alternativa para incorporar en el alimento vivo nutrientes que no son tradicionalmente encontrados en su composición (Smith et al. 2002).
Entre los indicadores empleados por investigadores y acuacultores para evaluar la producción larvaria en los sistemas experimentales y productivos se encuentran la supervivencia, el crecimiento en largo o peso y la calidad del producto determinado por la respuesta ante pruebas de estrés. Entre estas últimas, una de las más empleadas por la rapidez con que se realiza y que no demanda de equipo especializado es la prueba de resistencia al estrés salino (Palacios & Racotta 2007).
Si se tiene en cuenta que la historia nutricional de los organismos cultivados influye notablemente en las características de sus descendencias (Hagiwara & Hino 1990; Molony 1993), es de suponer que una adecuada alimentación de los primeros estadios favorecerá el crecimiento, la supervivencia y la resistencia al estrés de los organismos cultivados, lo que explica los resultados obtenidos por Castille et al. (1993), quienes indican que las diferencias observadas en el crecimiento de las postlarvas de L. vannamei de dos poblaciones diferentes se continúan expresando en el crecimiento de los juveniles.
Al tener en cuenta los diferentes requerimientos nutricionales que presentan los camarones en su desarrollo (Cuzon et al. 2004), se consideró importante determinar el efecto de nauplios de Artemia franciscana (Kellogg, 1906) enriquecida con dos emulsiones lipídicas sobre el crecimiento, supervivencia y resistencia al estrés osmótico en tres grupos de postlarvas de L. vannamei de diferentes edades (PL5-10, PL10-30, PL20-40).
MATERIALES Y MÉTODOS
Suministro de larvas
Las larvas de L. vannamei fueron obtenidas del laboratorio de camaronicultura comercial de Industrias Pecis S.A. de C.V. (Sisal , Mérida, Yucatán, México) y transferidas en estadio de nauplio II al Laboratorio de Ecología y Biología Marina Experimental de la UNAM en Ciudad del Carmen, Campeche, México. Las larvas fueron criadas hasta los estadios requeridos de postlarva en un recipiente de fibra de vidrio de 400 l donde se empleó un esquema de alimentación a base de diatomeas ( Chaetoceros gracilis), flagelados (Tetraselmis chuii) y nauplios de A. franciscana recién eclosionados (Aquatic EcoSystems Inc.) (Tabla 1).
Preparación de las emulsiones
Para el enriquecimiento de A. franciscana se prepararon dos tipos de emulsiones, la primera fue elaborada con lecitina de soya / aceite de hígado de bacalao / agua destilada en la siguiente proporción: 0.5 g / 4 g / 100 ml. Para evitar la pérdida excesiva de los ingredientes debido al contacto con las paredes del recipiente, los mismos fueron vertidos en un vaso de precipitado de 150 ml colocado sobre una balanza analítica en el siguiente orden: agua destilada, aceite y luego la lecitina.
La otra emulsión fue preparada con el producto comercial IKA W3©(aceite enriquecido de calamar) en la proporción de 4 g de producto en 100 ml de agua destilada. Este producto fue adquirido en "Sistemas de Equipo Especializado para Acuicultura (AQUASIS)" en México.
En ambos casos los ingredientes se emulsificaron en una licuadora doméstica a 812 rpm durante tres lapsos de 10 s con intervalos de receso entre ellos de 10 s. El contenido de la licuadora se colocó en un embudo separador que permitió desechar la espuma formada. Las emulsiones fueron caracterizadas mediante la observación a través de un microscopio biológico lo que permitió establecer las densidades de glóbulos ml-i para el posterior suministro como agente bioencapsulante en la preparación del medio enriquecedor.
Enriquecimiento de Artemia
En los ensayos se emplearon quistes de A. franciscana (Argentemia certified brine shrimp eggs, grade II). La eclosión se realizó en agua de mar a 33 ups. Para mantener los quistes en suspensión se suministró aireación y se mantuvo una iluminación constante de aproximadamente 1000 lux hasta la eclosión de los mismos.
Los nauplios cosechados se colocaron en agua de mar filtrada con T. chuii como alimento, a una densidad de 200 000 cel ml-i durante un período de 24 h. Los empleados en el tratamiento control (NANE) se mantuvieron otras dos horas en estas condiciones, se enjuagaron con agua de mar filtrada y se suministraron directamente a las postlarvas. Los empleados en los demás tratamientos se transfirieron a un recipiente donde recibieron el correspondiente tratamiento de enriquecimiento. La densidad de organismos en dichos recipientes se fijó en 250 nauplios ml-i . La concentración de los glóbulos de emulsión en el medio enriquecedor fue ajustada a 400 000 glóbulos ml-i en ambos tratamientos. Los nauplios de A. franciscana se mantuvieron bajo esas condiciones durante un período de 2 h (Gelabert 2003), después del cual los organismos fueron cosechados, enjuagados con agua de mar filtrada y suministrados a las postlarvas de acuerdo con el tratamiento correspondiente.
Ensayos con postlarvas de Litopenaeus vannamei
Tres ensayos se realizaron con postlarvas (PL) de Litopenaeus vannamei de tres grupos de edades (PL5-10, PL10-20, PL20-40), las cuales se determinaron de acuerdo al número de días transcurridos después de la metamorfosis de mysis III a PL. Las postlarvas fueron alimentadas con nauplios de A. franciscana enriquecida con lecitina de soya y aceite de hígado de bacalao (NA+L+AHB), nauplios de A. franciscana enriquecida con aceite enriquecido de calamar (NA+IKA W3) y A. franciscana no enriquecida (NANE), como dieta control. Cada tratamiento se realizó por triplicado.
Los animales se cultivaron en recipientes plásticos de 40 l de color blanco con 20 l de agua de mar filtrada y aireación constante, mantenidos a 27.5 ± 0.5 0C, salinidad de 38 ups, pH de 8.1 ± 0.1 y concentración de oxígeno mayor a 5.5 mg O2 l-1.
Para el primer ensayo (cinco días de experimentación) se utilizó una densidad de 20 PL5 l-1 (0.122 ± 0.034 mg de peso seco, n = 31), en el segundo ensayo (20 días de experimentación) la densidad fue de 1.5 PL10 l-1 (0.438 ± 0.117 mg de peso seco, n = 197), mientras que para el tercer ensayo (20 días de experimentación) la densidad inicial fue de 0.75 PL20 l-1 (5.06 ± 1.34 mg de pesos seco, n = 270).
El 75 % del medio de cultivo se recambió diariamente, lo cual incluyó la extracción de los sedimentos y remanentes de alimento. El volumen de agua se restableció con agua de mar filtrada (5 /xm) y esterilizada con luz ultravioleta. Una vez restablecido el volumen, se añadió la A. franciscana correspondiente a cada tratamiento y su densidad se ajustó entre 5 y 7.5 nauplios ml-1 .
El efecto de las dietas en las postlarvas de L. vannamei se evaluó mediante el crecimiento, supervivencia y resistencia al estrés.
Crecimiento
El crecimiento de los individuos fue determinado por el incremento en peso seco alcanzado durante el período experimental. El peso seco fue obtenido de muestras de cada unidad experimental. Las postlarvas fueron colocadas en una estufa a 60 0C por un período de 24 h y pesadas en una microbalanza CAHN modelo C-33 con una precisión de 0.001 mg. Como mínimo se pesaron diez postlarvas por unidad experimental. Los resultados fueron comparados mediante pruebas estadísticas paramétricas (ANDEVA) con un nivel de significación de a = 0.05 después de comprobar la normalidad y homocedasticidad de los datos (p > 0.05). En los casos en que se encontraron diferencias entre los tratamientos se aplicó la prueba Duncan como prueba de comparación múltiple de medias (Zar 1984).
Supervivencia
La supervivencia fue establecida por la proporción de individuos cosechados al final del experimento con respecto a la cantidad sembrada al inicio. Los resultados fueron comparados mediante una prueba no paramétrica de Chi cuadrada con una significación de p < 0.05.
Resistencia al estrés
Al finalizar cada ensayo se realizó una prueba de resistencia al estrés cuyo indicador fue obtenido al someter a todos los individuos a un estrés salino al reducir bruscamente la salinidad de 38 a 5 ups. Transcurrida una hora, se registró el número de individuos vivos y muertos. Con esta información se establecieron los porcentajes de supervivencia y los resultados se compararon con una prueba no paramétrica de Chi cuadrada con p < 0.05.
RESULTADOS
Ensayos con postlarvas PL5-10
El peso seco de las postlarvas alimentadas con NA+L+AHB y NA+IKA W3 resultó significativamente mayor que las alimentadas con la dieta control (NANE) (ANDEVA F = 6.56; p = 0.001) (Figura 1). Los incrementos en peso seco alcanzados por las postlarvas en los tratamientos NA+L+AHB y NA+IKA W3 no fueron significativamente diferentes (Duncan; p = 0.707).
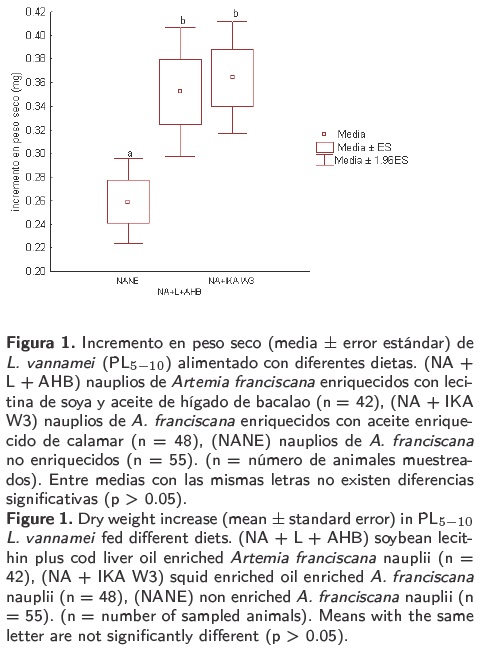
Los valores de supervivencia variaron entre 66 y 75 % en los diferentes tratamientos (Tabla 2). Estos resultados no difirieron significativamente entre sí (Chi cuadrada = 2.289; p = 0.318).
En las pruebas de resistencia al estrés los valores de supervivencia fluctuaron entre 82 y 83% para los organismos provenientes de los tres tratamientos (Tabla 2). Ninguna de las dietas utilizadas tuvo un efecto significativo sobre la resistencia de las postlarvas al estrés salino (Chi cuadrada = 0.460; p = 0.977).
Ensayo con postlarvas de PL10-30
En los tratamientos NA+L+AHB y NA+IKA W3 el crecimiento fue significativamente superior (ANDEVA F = 8.40; p = 0.000) al obtenido cuando se empleó la dieta NANE (Figura 2). Estos resultados concordaron con los obtenidos con las postlarvas de menor edad PL5-10).

Los valores de supervivencia oscilaron entre 88 y 90 % (Tabla 2). En este caso tampoco se observó un efecto significativo de las dietas sobre la supervivencia (Chi cuadrada = 0.204; p = 0.902).
Los valores obtenidos en la prueba de estrés se ubicaron entre 97 y 98 % de supervivencia en todos los tratamientos (Tabla 2). Estos resultados no difirieron estadísticamente (Chi cuadrada = 0.257; p = 0.879).
En general, el crecimiento, la supervivencia y la resistencia al estrés obtenidos en los tratamientos con PL10-30 concordó con el obtenido en PL5-10.
Ensayo con postlarvas de PL20-40
Los valores medios del crecimiento en peso seco obtenido fluctuaron entre 68 y 74 mg (Figura 3). En este caso, ninguno de los alimentos empleados tuvo un efecto significativo sobre el crecimiento (ANDEVA F = 1.05; p = 0.351).

La supervivencia para los tres tratamientos fue estadísticamente similar (Tabla 2, Chi cuadrada = 2.277; p = 0.320). Los valores se encontraron entre 95.4 y 98.8%, lo que reflejó que las condiciones de cultivo fueron favorables para las postlarvas de esta edad.
La prueba de estrés mostró resultados análogos para los tres tratamientos (Tabla 2). Todos los animales sobrevivieron al cambio brusco de salinidad, lo que se manifestó por la adecuada motilidad de los mismos en el momento de la culminación de la prueba.
DISCUSIÓN
La importancia de los ácidos grasos para el crecimiento de los peneidos ha recibido gran atención desde los trabajos iniciales realizados por Kanazawa et al. (1977). En Penaeus monodon ha sido demostrado el efecto promotor de diferentes ácidos grasos insaturados así como el balance entre ellos para lograr un crecimiento adecuado (Glencross & Smith 2001a y b; Glencross et al. 2002). El crecimiento significativamente superior obtenido con NA+L+AHB y NA+IKA W3 en los experimentos con PL5_10 y PL10_30, muestra que estas dietas cubrieron mejor los requerimientos de estas postlarvas que la dieta de NANE. Similar efecto del uso de nauplios de Artemia enriquecidos se ha registrado en estudios de alimentación de larvas de camarones (Kontara & Nurdjana 1992; Rees et al. 1994; Wouters et al. 1997; Immanuel et al. 2007), mientras que Martins et al. (2006) detectaron este efecto sólo en PL10 y no en PL1 de Farfantepenaeus paulensis.
Los pesos finales alcanzados por las postlarvas en estos experimentos con PL20_40 fueron considerablemente superiores, para la misma especie e intervalo de edad que los obtenidos por Coutteau et al. (1996). Estas diferencias se pueden justificar por las diferentes condiciones experimentales en que se desarrollaron los ensayos y por la variabilidad genética de los organismos empleados. Si se tiene en cuenta que el crecimiento refleja el efecto neto de las condiciones ambientales, incluido el alimento (Beamish et al. 1975), los resultados de este experimento pudieran ser considerados como indicadores de las condiciones adecuadas en que fueron desarrollados los experimentos.
Las condiciones de cultivo influyen notablemente en el crecimiento de las postlarvas de L. vannamei (Ogle 1992). En el presente trabajo, las condiciones de cultivo fueron similares para los tres experimentos y solamente varió la edad de los individuos. Por lo tanto, las diferentes respuestas registradas pueden ser atribuidas a la desigualdad en los requerimientos lipídicos de las postlarvas de diferente edad, que a su vez están relacionados con los cambios en la ontogenia del sistema digestivo por las que transitan para alcanzar esta etapa de desarrollo, (Lovett & Felder 1990a y b) y al cambio de hábitat y hábitos alimenticios en la etapa de postlarva. Una evidencia de ello fue presentada por Camara et al. (1997), quienes encontraron que el requerimiento de fosfatidilcolina decreció con la edad de las postlarvas de Marsopenaeus japonicus.
En ensayos realizados con postlarvas de peneidos, Tackaert et al. (1991) registraron que la presencia de fosfatidilcolina en la dieta de los organismos jóvenes mejoró los resultados de sobrevivencia y crecimiento, sin manifestarse diferencia en los resultados de la prueba de resistencia al estrés. En otra fase de cultivo, donde se trabajó con organismos de mayor talla, se detectó que los mejores crecimientos y resistencia al estrés se alcanzaron cuando los animales eran alimentados con dietas deficientes en fosfatidilcolina. Esto indica que los requerimientos nutricionales varían en dependencia de los estadios que se analicen.
De acuerdo con Watanabe et al. (1978) y Leger et al. (1986), la presencia de ácidos grasos altamente insaturados de la serie n3 determinan el valor nutrimental de nauplios de Artemia para larvas de peces marinos y crustáceos. Investigaciones sobre ácidos grasos esenciales han demostrado que para un normal crecimiento, las especies marinas requieren ácidos grasos altamente insaturados del tipo n3 tales como el eicosapentaenoico (20:5n3) y el docosahexaenico (22:6n3), así como el araquidónico (20:4n6) (Sargent et al. 1999). Estos son parte constituyente de las membranas celulares y juegan un papel importante en la formación de las branquias para la función de intercambio iónico que en las mismas se realiza (Palacios et al. 2004). En el presente trabajo no se registró la información de los perfiles de lípidos en las dietas probadas, sin embargo, el uso de dos dietas nutricionalmente reforzadas desde el punto de vista de su composición lipídica tuvo un efecto positivo en el crecimiento de los organismos en los primeros estadios de desarrollo postlarval. Cuando las mismas dietas fueron usadas en postlarvas de mayor edad no se manifestó el mismo efecto. Wouters et al. (2001) mencionaron que una excesiva cantidad de lípidos totales en la dieta puede tener un efecto adverso en la maduración gonadal en L. vannamei, posiblemente debido a la manifestación de una sensación de saciedad en los organismos y a la reducción del consumo de alimento derivada de la misma. Es probable que este efecto se pudiera estar presentando en las postlarvas mayores estudiadas en el presente trabajo. Las diferentes respuestas a los tratamientos también pueden ser explicadas según Kanazawa (1993) por la incapacidad de las postlarvas más jóvenes de sintetizar fosfolípidos a una tasa suficientemente elevada como para cubrir los requerimientos nutricionales en este periodo que presenta un crecimiento acelerado. En Marsupenaeus japonicus se demostró que el efecto positivo de la incorporación de fosfolípidos en la dieta se debió a que propiciaron una mejor movilización de los lípidos desde la glándula digestiva o el intestino hacia la hemolinfa, resultando en una mayor deposición de los lípidos en los tejidos y un incremento de la energía disponible para el crecimiento (Kontara et al. 1997). En el presente trabajo, la lecitina de soya se empleó como agente emulsificante del aceite de hígado de bacalao, generalmente los fosfolípidos de ésta son una mezcla de fosfatidil colina, fosfatidil etanolamina, fosfatidil inositol y ácido fosfatídico (Gong et al. 2000). Entre la lecitina y el aceite de hígado de bacalao empleados en el tratamiento NA+L+AHB se aportaron ácidos grasos insaturados y polinsaturados como el oleico, linolenico, eicosapentaenoico y docosahexaenoico. La emulsión IKA W3, compuesta por aceite de calamar enriquecido, también es una fuente rica en ácidos grasos polinsaturados que contribuyen al transporte de lípidos de forma más eficiente, mejoran la fluidez y permeabilidad de las membranas así como cubren requerimientos no satisfechos en las postlarvas menores por los nauplios de Artemia franciscana no enriquecidos.
Para los acuacultores e investigadores, la supervivencia es un importante indicador para la toma de decisiones relacionadas con la modificación de las condiciones de cultivo y además refleja las condiciones en las que los organismos son mantenidos. Altas supervivencias están directamente relacionadas con condiciones apropiadas de mantenimiento, sin embargo, Kuban et al. (1985), analizaron el efecto de seis combinaciones de alimentos diferentes en larvas de cuatro especies de peneidos y concluyeron que el crecimiento en biomasa es mejor indicador que la supervivencia para determinar el valor nutricional de una dieta.
Por otra parte, Lunn & Htoo (1997) estimaron un efecto positivo en la supervivencia de Macrobrachium rosenbergii cuando fue alimentado con Artemia enriquecida. Los mismos resultados fueron obtenidos para Farfantepenaeus paulensis (Martins et al. 2006). En contraste, en ensayos con postlarvas de Penaeus monodon, alimentadas con aceites de peces emulsificados con yema de huevo, no se registraron diferencias en la supervivencia (Kontara & Nurdjana 1992). En postlarvas de L. vannamei no se encontró efecto positivo en la supervivencia, ni en la resistencia a un estrés salino debido a la inclusión en la dieta de fosfatidilcolina obtenida de diferentes fuentes. Sin embargo, un incremento significativo se detectó en el crecimiento y en el contenido de ácidos grasos polinsaturados de los individuos evaluados (Coutteau et al. 1996; 2000). En juveniles de la misma especie González-Félix et al. (2002) tampoco encontraron efecto en la supervivencia cuando emplearon dietas con diferentes cantidades y fuentes de lípidos, aunque sí obtuvieron un efecto positivo sobre el crecimiento. Las elevadas supervivencias alcanzadas y los resultados de las pruebas de estrés en las diferentes clases de edad inducen a pensar que las dietas usadas en este trabajo (incluyendo la dieta control) garantizan el mantenimiento de las condiciones nutricionales mínimas para los estadios de postlarvas estudiados.
En los casos en que el periodo de los ensayos sea relativamente corto y el análisis del crecimiento y la supervivencia no muestren diferencias significativas entre los tratamientos, es conveniente emplear otros indicadores que permitan evaluar el efecto de los tratamientos en los organismos. Con este propósito se han empleado pruebas que miden la resistencia de los organismos a un estrés y los resultados son expresados generalmente con un índice que refleja la supervivencia de los organismos. Diversos factores influyen sobre el resultado de la prueba, pero en un diseño donde sólo una variable sea modificada, como es el caso de este trabajo (edad), las similitudes o diferencias encontradas en los resultados de la prueba, se deberán al efecto de esa variable.
Según Dhert et al. (1992a), el efecto beneficioso del suministro de ácidos grasos a través de las técnicas de bioencapsulación con Artemia como vector puede ser claramente demostrado mediante una prueba de estrés, aún antes de que las mortalidades se manifiesten. Por este motivo estos autores argumentan la necesidad de emplear este indicador para evaluar diariamente la condición de los animales y contar con elementos para modificar el manejo y las condiciones de cultivo. Los resultados del presente estudio se explican por la tolerancia de L. vannamei a medios donde la salinidad oscila entre 2 y 60 ups. Mair (1980) encontró que en los camarones se altera la preferencia por la salinidad al crecer y McGraw et al. (2002) lo demostraron en su trabajo sobre la aclimatación de postlarvas de L. vannamei a baja salinidad. Los resultados además corroboraron que las pruebas de estrés de salinidad pueden ser menos adecuadas para L. vannamei, ya que es una especie eurihalina que en condiciones naturales puede sobrevivir a salinidades entre 2 y 40 ups (Coutteau et al. 1996). Es conocido que L. vannamei incrementa la resistencia a la salinidad con el desarrollo (Dhert et al. 1992b; Samocha et al. 1998), por este motivo los autores recomiendan utilizar pruebas más drásticas para evaluar el efecto de los tratamientos en los organismos. Por lo anterior, para reforzar los resultados de la prueba de estrés de salinidad tradicionalmente empleada se han descrito otras pruebas donde se ha combinado el efecto de la salinidad con formalina (Samocha et al. 1998) y la salinidad con la temperatura (Briggs 1992). En el presente trabajo los resultados de la prueba mostraron que no hubo efecto de las diferentes dietas sobre este índice por lo que las dietas pueden ser consideradas igualmente satisfactorias.
Los nauplios de A. franciscana enriquecidos con lípidos favorecieron el crecimiento de L. vanna-mei cuando se suministraron a los primeros estadios de postlarva, sin manifestarse efecto significativo en la supervivencia ni en la resistencia al estrés salino. Si bien los protocolos de enriquecimiento de plantas de producción de larvas utilizan tiempos de enriquecimiento mucho mayores que los de este trabajo, el hecho de realizar la bioencapsulación de Artemia en un tiempo relativamente corto y encontrar efectos satisfactorios en el crecimiento de los individuos evaluados puede ser un indicador de que no es necesario realizar un enriquecimiento tan prolongado durante el que se reduce la concentración de oxígeno y se favorece el crecimiento bacteriano en el medio. Las ventajas o desventajas del uso de periodos cortos de enriquecimiento deberán evaluarse en otros estudios ya que pueden resultar muy atractivos para los productores. Desde el punto de vista técnico, los resultados del trabajo promueven el empleo de A. franciscana enriquecida en las primeras etapas de postlarvas e indican que es más importante garantizar una apropiada alimentación de estos estadios que reforzar la alimentación en las etapas más avanzadas.
AGRADECIMIENTOS
Los autores aprecian el apoyo recibido por Adriana Paredes, Gabriela Palomino, Gabriel Taboada y Claudia Durruty, miembros de la Unidad Multidisciplinaria de Docencia e Investigación de la Universidad Nacional Autónoma de México. Este trabajo fue parcialmente financiado por el proyecto No. DGAPA-UNAM In 220502-3.
LITERATURA CITADA
Beamish FWH, Miimi AJ, Leet PIKP (1975) Bioenergetic of Teleost fishes: Enviromental influence. In: Bolis Madreel LHP, Schmidt-Nielsen K (eds) Comparative Funcional Aspects of Structutral Materials. Nosth Holland Publications Company, Amsterdan. 600 pp. [ Links ]
Briggs MRP (1992) A stress test for determining vigour of post-larval Penaeus monodon Fabricius. Aquacult. Fish. Manage. 23: 633-637. [ Links ]
Camara MR, Coutteau P, Sorgeloos P (1997) Dietary phosphatidylcholine requirements in larval and postlarval Penaeus japonicus Bate. Aquacult. Nutr. 3: 39-47. [ Links ]
Castille FL, Samocha TM, Lawrence AL, He H, Frelier P, Jaenike F (1993) Variability in growth and survival of early postlarval shrimp (Penaeus vannamei Boone 1931). Aquaculture 113: 65-81. [ Links ]
Cheng ZJ, Hardy RW (2004) Protein and lipid sources affect cholesterol concentrations of juvenile Pacific white shrimp, Litopenaeus vannamei (Boone). J. Anim. Sci. 84: 1136-1145. [ Links ]
Clawson JA, Lovell RT (1992) Improvement of nutritional value of Artemia for hybrid striped bass/white bass (Morone saxatilis x M. chrysops) larvae by n-3 HUFA enrichment of nauplii with menhaden oil. Aquaculture 108: 125-134. [ Links ]
Coutteau P, Camara MR, Sorgeloos P (1996) The effect of different levels and sources of dietary phosphatidylcholine on the growth, survival, stress resistance, and fatty acid composition of postlarval Penaeus vannamei. Aquaculture 147: 261-273. [ Links ]
Coutteau P, Kontara EKM, Sorgeloos P (2000) Comparison of phosphatidylcholine purified from soybean and marine fish roe in the diet of postlarval Penaeus vannamei Boone. Aquaculture 181: 331-345. [ Links ]
Cuzon G, Lawrence AL, Gaxiola G, Rosas C, Guillaume J (2004) Nutrition of Litopenaeus vannamei reared in tanks or in ponds. Aquaculture 235: 513-551. [ Links ]
Czesny S, Kolkovski S, Dabrowski K, Culver D (1999) Growth, survival, and quality of juvenile walleye Stizostedion vitreum as influenced by n-3 HUFA enriched Artemia nauplii. Aquaculture 178: 103-115. [ Links ]
Dhert P, Lavens P, Sorgeloos P (1992a) Stress evaluation: A tool for quality control of hatchery-produced shrimp and fish fry. Aquacult. Eur. 17: 6-10. [ Links ]
Dhert PH, Lavens P, Sorgeloos P (1992b) A simple test for quality evaluation of cultured fry of marine fish. Med. Fac. Landbouww Univ. 57: 2135-2142. [ Links ]
Fernández-Reiriz MJ, Labarta U, Ferreiro MJ (1993) Effects of commercial enrichment diets on the nutritional value of the rotifer (Brachionus plicatilis). Aquaculture 112: 195-206. [ Links ]
Gara B, Shields RJ, McEvoy L (1998) Feeding strategies to achieve correct metamorphosis of Atlantic halibut, Hippoglossus hippoglossus L., using enriched Artemia. Aquacult. Res. 29: 935-948. [ Links ]
Gelabert R (2003) Bioencapsulation in Artemia: II. Influences of the particle concentration in the enrichment process. Aquaculture 216: 143-153. [ Links ]
Glencross BD, Smith DM (2001a) A study of the arachidonic acid requirements of the giant tiger prawn, Penaues monodon. Aquacult. Nutr. 7: 59-69. [ Links ]
Glencross BD, Smith DM (2001b) Optimizing the essential fatty acids, eicosapentaenoic and docosahexaenoic acid, in the diet of the prawn, Penaeus monodon. Aquacult. Nutr. 7: 101-112. [ Links ]
Glencross BD, Smith M, Curnow J, Smith D, Williams K (2001) The dietary protein and lipid requirements of post-puerulus western rock lobster, Panulirus cygnus. Aquaculture 199: 119-129. [ Links ]
Glencross BD, Smith M, Thomas M, Williams K (2002) Optimising the essential fatty acids in the diet for weight gain of the prawn, Penaeus monodon. Aquaculture 204: 85-99. [ Links ]
Gómez-Gil B, Cabanillas-Ramos J, Páez-Brambila S, Roque A (2001) Standardization of the bioencapsulation of enrofloxacin and oxytetracycline in Artemia franciscana Kellogg, 1906. Aquaculture 196: 1-12. [ Links ]
Gong H, Lawrence AL, Jiang DH, Gatesoupe FJ (2000) Lipid nutrition of juvenile Litopenaeus vannamei II. Active components of soybean lecithin. Aquaculture 190: 325-342. [ Links ]
González-Félix ML, Gatlin III DM, Lawrence AL, Pérez-Velázquez M (2002) Effect of dietary phospholipid on essential fatty acid requirements and tissue lipid composition of Litopenaeus vannamei juveniles. Aquaculture 205: 325-343. [ Links ]
Hagiwara A, Hino A (1990) Feeding history and hatching of resting eggs in the marine rotifer Brachionus plicatilis. Bull. Jap. Soc. Sci. Fish. 56: 1965-1971. [ Links ]
Han K, Geurden I, Sorgeloos P (2000) Enrichment strategies for Artemia using emulsions providing different levels of n-3 highly unsaturated fatty acids. Aquaculture 183: 335-347. [ Links ]
Immanuel G, Citarasu T, Sivaram M, Michael Babu M, Palavesam A (2007) Delivery of HUFA, probionts and biomedicine through bioencapsulated Artemia as a means to enhance the growth and survival and reduce the pathogenesity in shrimp Penaeus monodon postlarvae. Aquacult. Int. 15: 137-152. [ Links ]
Kanazawa A (1993) Essential phospholipids of fish and crustaceans. In: Kaushik SJ, Luquet P (eds) Fish Nutrition in Practice. Edition INRA. Paris, Les Colloques 61: 519-530. [ Links ]
Kanazawa A, Tokiwa S, Kayama M, Hirata M (1977) Essential fatty acids in the diet of prawn. 1. Effects of linoleic and linolenic acids on growth. Bull. Jap. Soc. Sci. Fish. 43: 1111-1114. [ Links ]
Kontara EK, Nurdjana ML (1992) Growth and survival of Penaeus monodon postlarvae fed with Artemia nauplii enriched with n-3 HUFA. Bull. Brackishwat. Aquacult. Dev. Cent. Jepara 9: 48-55. [ Links ]
Kontara EK, Coutteau P, Sorgeloos P (1997) Effect of dietary phospholipid on requirements for an incorporation of n-3 highly unsaturated fatty acids in postlarval Penaeus japonicus Bate. Aquaculture 158: 305-320. [ Links ]
Kuban FD, Lawrence AL, Wilkenfeld JS (1985) Survival, metamorphosis and growth of larvae from four penaeid species fed six food combinations. Aquaculture 47: 151-162. [ Links ]
Leger P, Bengtson DA, Simpson KL, Sorgeloos P (1986) The use and nutritional value of Artemia as a food source. Oceanogr. Mar. Biol. Annu. Rev. 24: 521-623. [ Links ]
Lovett DL, Felder DL (1990a) Ontogenetic change in digestive enzyme activity of larval and postlarval white shrimp Penaeus setiferus (Crustacea, Decapoda, Penaeidae). Biol. Bull. 178: 144-159. [ Links ]
Lovett DL, Felder DL (1990b) Ontogenetic changes in enzyme distribution and midgut function in developmental stages of Penaeus setiferus (Crustacea, Decapoda, Penaeidae). Biol. Bull. 178: 160-174. [ Links ]
Lunn UZ, Htoo DTT (1997) Bioencapsulation technology used in the larval rearing of Macrobrachium rosenbergii. Support to the special plan for shrimp and prawn farming. FAO. Myanmar: 1-16. [ Links ]
Mair JM (1980) Salinity and water-type preferences of four species of postlarval shrimp (Penaeus) from west Mexico. J. Exp. Mar. Biol. Ecol. 45: 69-82. [ Links ]
Martins TG, Cavalli RO, Martino RC, Rezende CEM, Wasielesky WJr (2006) Larviculture output and stress tolerance of Farfantepenaeus paulensis postlarvae fed Artemia containing different fatty acids. Aquaculture 252: 525-533. [ Links ]
McGraw WJ, Davis DA, Teichert-Coddington D, Rouse DB (2002) Acclimation of Litopenaeus vannamei poslarvae to low salinity: Influence of age, salinity endpoint, and rate of salinity reduction. J. World Aquacult. Soc. 33: 78-84. [ Links ]
Molony BW (1993) Effects of feeding history on mobilization and deposition of body constituents and on growth in juvenile Ambassis vachelli (Pisces: Chandidae). Mar. Biol. 116: 389-397. [ Links ]
Narciso L, Pousao-Ferreira P, Passos A, Luis O (1999) HUFA content and DHA/EPA improvements of Artemia sp. with commercial oils during different enrichment periods. Aquacult. Res. 30: 21-24. [ Links ]
Ogle JT (1992) Variability in growth of postlarval Penaeus vannamei. Gulf Res. Rep. 8: 423-426. [ Links ]
Palacios E, Racotta IS (2007) Salinity stress test and its relation to future performance and different physiological responses in shrimp postlarvae. Aquaculture 268: 123-135. [ Links ]
Palacios E, Bonilla A, Pérez A, Racotta IS, Civera R (2004) Influence of highly unsaturated fatty acids on the response of white shrimp (Litopenaeus vannamei) postlarvae to low salinity. J. Exp. Mar. Biol. Ecol. 299: 201-215. [ Links ]
Palacios E, Racotta IS, Heras H, Marty Y, Moal J, Samain JF (2002) Relation between lipid and fatty acid composition of eggs and larval survival in white pacific shrimp (Penaeus vannamei Boone, 1931). Aquacult. Int. 9: 531-543. [ Links ]
Rees JF, Cure K, Piyatiratitivorakul S, Sorgeloos P, Menasveta P (1994) Highly unsaturated fatty acid requirements of Penaeus monodon postlarvae: An experimental approach based on Artemia enrichment. Aquaculture 122: 2-3. [ Links ]
Rodríguez C, Pérez JA, Izquierdo MS, Cejas JR, Bolanos A, Lorenzo A (1996) Improvement of the nutritional value of rotifers by varying the type and concentration of oil and the enrichment period. Aquaculture 147: 93-105. [ Links ]
Sakamoto M, Holland DL, Jones DA (1982) Modification of the nutritional composition of Artemia by incorporation of polyunsaturated fatty acids using micro-encapsulated diets. Aquaculture 28: 311-320. [ Links ]
Samocha TM, Guajardo H, Lawrence AL, Castille FL, Speed M, McKee DA, Page KI (1998) A simple stress test for Penaeus vannamei postlarvae. Aquaculture 165: 233-242. [ Links ]
Sargent J, McEvoy L, Estévez A, Bell G, Bell M, Henderson J, Tocher D (1999) Lipid nutrition of marine fish during early development: current status and future directions. Aquaculture 179: 217-229. [ Links ]
Smith GG, Ritar AJ, Phleger CF, Nelson MM, Mooney B, Nichols PD, Hart P (2002) Changes in gut content and composition of juvenile Artemia after oil enrichment and during starvation. Aquaculture 208: 137-158. [ Links ]
Sorgeloos P, Coutteau P, Dhert P, Merchie G, Lavens P (1998) Use of Brine Shrimp, Artemia spp., in Larval Crustacean Nutrition: A Review. Rev. Fish. Sci. 6: 55-68. [ Links ]
Southgate PC, Lou DC (1995) Improving the n-3 HUFA composition of Artemia using microcapsules containing marine oils. Aquaculture 134: 91-99. [ Links ]
Tackaert W, Camara MR, Sorgeloos P (1991) The effect of dietary phosphatidylcholine in postlarval penaeid shrimp: I. Diet preparation. En: Lavens P, Sorgeloos P, Jaspers E, Ollevier F (eds) Larvi'91-Fish and Crustacean Larviculture Symposium. EAS Special Publication No. 15, Ghent: 76-79. [ Links ]
Watanabe T, Oowa F, Kitajima C, Fujita S (1978) Nutritional quality of brine shrimp, Artemia salina, as a living feed from the viewpoint of essential fatty acids for fish. Bull. Jap. Soc. Sci. Fish. 44: 1115-1121. [ Links ]
Wouters R, van Hauwaert A, Naessens E, Ramos X, Pedrazzoli A, Lavens P (1997) The effect of dietary n-3 HUFA and 22:6n-3/20:5n-3 ratio on white shrimp larvae and postlarvae. Aquacult. Int. 5: 113-1126. [ Links ]
Wouters R, Lavens P, Nieto J, Sorgeloos P (2001) Penaeid shrimp broodstock nutrition: an updated review on research and development. Aquaculture 202: 1-21. [ Links ]
Zar JH (1984) Biostatistical Analysis. Prentice-Hall. Englewood Cliffs. 718 pp. [ Links ]


{kind=link}