










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkUniversidad y ciencia
versão impressa ISSN 0186-2979
Universidad y ciencia vol.24 no.3 Villahermosa Dez. 2008
Artículos
Comportamiento reproductivo de ovejas F1 (Damara x Merino) sincronizadas con CIDR y dos tiempos de aplicación de GnRH
Reproductive performance of F1 (Damara x Merino) ewes synchronised with CIDR and two times of GnRH injection
JJ Martínez–Tinajero1*, MT Sánchez Torres–Esqueda2, G Torres–Hernández2, JG Herrera–Haro2, L Bucio–Alanís3, R Rojo–Rubio4 y J Hernández–Martínez4
1 Facultad de Ciencias Agrícolas de la Universidad Autónoma de Chiapas. Calle Río Cosalapa No. 23, Fraccionamiento Las Hortensias, CP. 30797, Tapachula, Chiapas, México. Tel. (962) 64 70128 Fax: (964) 62 70439. (JJMT) *Correo electrónico: jaimej@unach.mx, jaimejorge@gmail.com
2 Programa de Ganadería del Colegio de Postgraduados, Montecillos, Texcoco, estado de México. (MTSTE)(GTH)(JGHH)
3 Campus Córdoba del Colegio de Postgraduados, Córdoba, Veracruz. (LBA)
4 Centro Universitario UAEM–Temascaltepec. Universidad Autónoma del Estado de México. (RRR)(JHM)
Artículo recibido: 16 de julio de 2006
Aceptado: 8 de agosto de 2008
RESUMEN
Este experimento se realizó para evaluar el comportamiento reproductivo en ovejas F1 (Damara x Merino) importadas de Australia, sincronizadas con un dispositivo liberador de la hormona (CIDR) y dos tiempos de aplicación de hormona liberadora de gonadotropinas (GnRH). Cuarenta y cinco ovejas F1 de primer parto con 18.08 ± 0.07 meses de edad y 43.3 ± 5.6 kg de peso corporal, fueron asignadas aleatoriamente a uno de tres tratamientos: T1 (n = 15): CIDR por doce días; T2 (n = 15): CIDR por doce días mas 25 μg de GnRH 48 h antes del retiro del dispositivo y T3 (n = 15): CIDR por doce días mas 25 μg de GnRH al momento del retiro del CIDR. El porcentaje de presentación de estros (100%) fue similar (p > 0.05) para T1, T2 y T3. El intervalo (h) de inicio del estro sincronizado después del retiro del CIDR fue diferente (p <0.05) en T2 y T3 (43.86 ± 5.42 y 45.73 ± 3.36 h) comparado con T1 (T1 = 37.33 ± 7.90 h). El porcentaje de fertilidad a estro sincronizado fue similar (p > 0.05) para T1, T2 y T3 (60, 53.3 y 46.6%, respectivamente). La prolificidad promedio general fue de 100.8% y fue similar (p > 0.05) entre tratamientos. La concentración de progesterona determinó que las ovejas probadas tuvieron un cuerpo luteo funcional al comienzo del experimento. Se concluye que el uso combinado de CIDR mas GnRH, 48 h antes y al momento del retiro del dispositivo, no tiene efecto sobre el comportamiento reproductivo en ovejas F1 (Damara x Merino).
Palabras clave: GnRH, CIDR, reproducción, sincronización de estros, progesterona, ovejas.
ABSTRACT
The experiment was carried out to evaluate the reproductive performance of F1 (Damara x Merino) ewes imported from Australia, synchronised with controlled internal drugs (CIDR) and two times of injection of gonadotropin releasing hormone (GnRH). Forty five F1 ewes at first lambing, of 18.08 ± .07 months of age and 43.3 ± 5.6 kg of body weight were randomly assigned to one of three treatments: T1 (n = 15): CIDR for 12 days; T2 (n = 15): CIDR for 12 days plus 25 μg of GnRH 48 h before CIDR withdrawal and T3 (n = 15): CIDR for 12 days plus 25 μg of GnRH at the moment of CIDR withdrawal. The percentage of estrus presentation (100%) was similar (p > 0.05) for T1, T2 and T3. The interval (h) of the onset of synchronised estrus after CIDR removal was different (p <0.05) for T2 and T3 (43.8 ± 5.4 and 45.7 ± 3.3 h) compared with T1 (37.3 ± 7.9 h). The percentage of fertility at the synchronised estrus was similar (p > 0.05) for T1, T2 and T3 (60.0, 53.3 y 46.6%, respectively). The general average prolificacy was 100.8% and similar (p > 0.05) among treatments. The concentration of progesterone insured that the tested ewes had a functional corpus luteum at the beginning of the experiment. It is concluded that the combined use of CIDR and GnRH, 48 h before and at the moment of CIDR removal, has no effect on the reproductive performance of F1 (Damara x Merino) ewes.
Key words: CIDR, GnRH, reproduction, estrus synchronisation, progesterone, ewes.
INTRODUCCIÓN
Durante los últimos años, el manejo reproductivo del ganado ovino ha evolucionado considerablemente mediante el empleo de tecnologías económicamente factibles, las cuales han incrementado la productividad en los rebaños. Este desarrollo se caracteriza por la utilización de diferentes métodos para inducir la sincronización de estros, los cuales ya han sido ampliamente revisados. Entre los métodos hormonales se hallan los progestágenos (Simonetti et al. 1999) y las prostaglandinas (Hernández et al. 2001). Los métodos biológicos comprenden el empleo del efecto macho (González et al. 2000) y del efecto hembra (Álvarez & Zarco 2001).
Los tratamientos hormonales para la sincronización de estros, así como para la inducción de ovulaciones, se han basado en la utilización combinada de hormonas con el uso de un agente bloqueador de los ciclos estrales y ovulaciones fisiológicas (progesterona y sus derivados sintéticos), así como agentes inductores de la ovulación y apoyo luteotrópico, como la gonadotropina coriónica equina (eCG) y hormona liberadora de gonadotropinas (GnRH) (Amiridis et al. 2005). Sin embargo, existe escasa información sobre la utilización de GnRH en ganado ovino, ya que la mayoría de las investigaciones se han generado en bovinos (Sepúlveda et al. 2003; Gutiérrez et al. 2005) y en condiciones diferentes a las de las zonas tropicales de México. La finalidad de combinar dispositivos para sincronizar el estro, más la aplicación de GnRH, permitirían inducirlo fuera de la época reproductiva, y así agrupar los estros en tiempos más cortos, concentrar los partos en determinada época del año y reducir en forma considerable el intervalo entre partos (Akif & Kuran 2003).
La raza Damara, originaria de Namibia, África, y desarrollada comercialmente desde 1996 por los Australianos (Fleet & Bennie 2002). Esta raza se caracteriza por su rusticidad a condiciones adversas y presentar actividad ovárica durante todo el año, acompañada de altos porcentajes de fertilidad y prolificidad. Por estas características, fueron importados a México varios sementales y hembras puras de esta raza. Las cruzas de Damara con Merino, se efectuaron con la finalidad de mejorar el comportamiento productivo y reproductivo de las razas nativas de México.
El objetivo del presente estudio tuvo como finalidad, evaluar la respuesta a sincronización de estros (porcentaje de hembras que presentan estros e inicio del estro en horas), porcentaje de fertilidad y prolificidad de ovejas F1 (Damara X Merino) sincronizadas con un dispositivo liberador de la hormona (CIDR) durante 12 días impregnado con 0.3 g de progesterona y dos tiempos de aplicación de GnRH bajo condiciones de clima tropical húmedo de México.
MATERIALES Y MÉTODOS
El estudio se inició en noviembre de 2002 y finalizó en mayo de 2003 en el Campus Córdoba del Colegio de Postgraduados, ubicado en el km 26.4 de la carretera federal Córdoba–Veracruz (18° 27' N y 96° 21' O) a 300 msnm. El clima de la región es AW2 (w) (i) g.e. que corresponde a un clima cálido húmedo con lluvias en verano, con menos de cinco por ciento de lluvia invernal y el mes más caliente antes del solsticio de verano. La precipitación anual promedio es 2 200 mm y la temperatura media anual 27.3 °C (del Amo et al. 1991). Se utilizaron 45 ovejas de primer parto, 18.08 ± 0.07 meses de edad, distribuidas en tres tratamientos: tratamiento 1 (T1: n = 15, PV: 42.3 ± 0.8 kg) CIDR impregnado con 0.3 g de progesterona natural (Eazi–Breed© CIDR©, Inter Ag, Nueva Zelanda) durante 12 días, tratamiento 2 (T2: n = 15, PV: 44.3 ± 1.2 kg): CIDR por 12 días más 25 μg de GnRH (Ovalyse, Pharmacia & Upjohn) por vía parenteral 48 h antes del retiro del dispositivo y tratamiento 3 (T3: n = 15, PV: 40.8 ± 1.6 kg): CIDR por 12 días más 25 μg de GnRH aplicado por vía parenteral al momento del retiro del dispositivo. Todos los animales, previo al inicio del experimento, se desparasitaron con 4 ml de albendazole (Albendavedi con Se Co, Vedi), se inyectaron con 1 ml/animal de vitamina ADE (Vigantol, Bayer) y se vacunaron con 2.5 ml de Bacterina toxoide (Triangle 8V, Fort Dodge). Durante todo el experimento las ovejas se mantuvieron en diferentes corrales (6 X 8 m) con comederos y bebederos, de acuerdo al tratamiento establecido, donde fueron alimentadas con pasto Taiwan (Pennisetum purpureum) ad libitum, 800 g animal–1 día–1 de un suplemento alimenticio con 22% de proteína (sorgo molido 40 %, maíz molido 20 %, pasta de soya 15 %, harina de pollo 10%, melaza 6%, grasa de sobrepaso 5 %, urea 2 % y mezcla de sales minerales 2 %) y agua fresca ad libitum.
El momento del retiro del dispositivo se consideró como el día cero. La detección de estros se realizó durante los primeros cinco días después de haber retirado los CIDR. En los primeros tres días, las ovejas se expusieron a los sementales cada dos horas y cada 12 horas en los dos días subsecuentes. Seis sementales de fertilidad comprobada se utilizaron en empadres anteriores. Cuando la oveja permitió la monta del semental, se consideró que ésta se encontraba en estro. Asimismo, cuando una oveja fue copulada por primera vez por el semental en turno, otro servicio se dio a las 12 horas posteriores con otro semental e inmediatamente se retiró del sitio de apareamiento.
Para conocer el estado fisiológico reproductivo de las ovejas al inicio y al final del experimento se determinó la concentración plasmática de progesterona. Las muestras de sangre se recolectaron cada 48 h, seis días antes de la aplicación del CIDR hasta 18 días después de su retiro (Roelofs et al. 2006), en 30 ovejas que se seleccionaron en forma aleatoria (n = 10/tratamiento). La obtención de sangre se hizo por punción de la vena yugular y se almacenó en tubos de ensayo, la cual fue centrifugada (3000 g x 15 min) y congelada a – 4 °C hasta ser analizada. Las concentraciones de progesterona se determinaron por radioinmunoensayo (RIA) en fase sólida con un paquete comercial (COAT–A–COUNT®, DPC, USA) en el laboratorio de Fisiología de la Reproducción de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Nacional Autónoma de México (Srinkandakumar et al. 1986). Las ovejas con concentraciones de progesterona superiores a 1 ng mL–1 en 5 muestras consecutivas se consideraron con cuerpo lúteo funcional (Vincent et al. 2000).
El porcentaje de presentación de estros se determinó durante los cinco días posteriores al retiro del dispositivo, inicio del estro, porcentaje de fertilidad y prolificidad y concentración de progesterona plasmática. Con el propósito de evaluar el porcentaje de fertilidad y prolificidad entre los grupos experimentales se realizó el diagnóstico de gestación por ecografía transrectal a los 50 días postmonta y así registrar los partos de abril a mayo.
El porcentaje de presentación de estros se determinó dividiendo el número de ovejas que exhibieron estro durante el tratamiento entre el número total de ovejas sometidas a ese tratamiento. La fertilidad se refirió al número total de ovejas diagnosticadas gestantes entre el número de ovejas servidas por el semental. El inicio del estro se determinó en el momento en que la oveja se dejó montar por el semental. La prolificidad se determinó dividiendo el número de corderos nacidos entre el número de ovejas paridas multiplicado el resultado por 100.
Los datos obtenidos del inicio del estro se analizaron con ANOVA y la concentración de progesterona plasmática se analizó mediante medidas repetidas del procedimiento GLM del paquete estadístico SAS (Anonymous 1999). El porcentaje de presentación de estros, fertilidad y prolificidad se analizaron utilizando la prueba de distribución libre Ji Cuadrada. La comparación de medias se realizó utilizando la prueba de Tukey (Steel y Torrie, 1981). El modelo estadístico utilizado para el inicio del estro en horas fue:
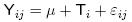
donde: Yij = valor de la variable en la j–ésima repetición del i–ésimo tratamiento.
μ = media general.
Ti = efecto del i–ésimo tratamiento i = 1, 2, 3.
εij= error experimental 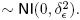
RESULTADOS
El 100 % de las ovejas presentaron estro durante los primeros cinco días posteriores a la remoción del CIDR y no se estimaron diferencias (p > 0.05) entre tratamientos. En las primeras 24 h ninguna oveja de los tres tratamientos presentó signos de estro (Figura 1). De las 24 a las 36 h se detectó estro en 10 ovejas (22.2%), de las 36 a las 48 h se observó estro en 28 ovejas (62.2 %) y después de las 48 h se presentó estro en siete ovejas (15.5%) de los estros totales.
El tiempo promedio del inicio del estro fue de 42.31 ± 6.80 h después del retiro de los dispositivos no existiendo diferencias (p > 0.05) entre tratamientos. Los valores promedio del inicio del estro fueron 37.33 ± 7.90, 43.86 ± 5.42 y 45.73 ± 3.36 h para T1, T2 y T3, respectivamente.
El porcentaje de fertilidad fue de 60.0, 53.3 y 46.6 %, para T1, T2 y T3, respectivamente (Tabla 2) (1). No se estimaron diferencias significativas entre los tratamientos (p > 0.05). Del mismo modo, el porcentaje de prolificidad fue de 100, 102.6 y 100 % para T1, T2 y T3, respectivamente, y estos valores no fueron significativamente diferentes (p > 0.05) entre los grupos experimentales.


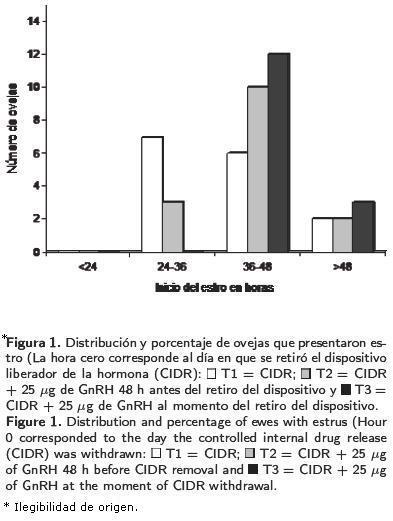
El promedio general en la concentración de progesterona durante toda la fase experimental fue vamente, resultaron diferentes (p < 0.05) entre los tres tratamientos. En la Figura 2 se observa la variación del esteroide durante el periodo de estudio.
DISCUSIÓN
Los resultados del inicio del estro fueron similares a los reportados por Kridli et al. (2003), quienes registraron intervalos que oscilaron entre las 40 a 50 h de presentación de estros después del retiro del CIDR. Aun cuando el inicio del estro fue similar en los tres tratamientos, se observó que las ovejas de T2 y T3 presentaron una mayor agrupación de los estros entre las 36 y 48 h después de la remoción del CIDR, lo cual coincide con el tiempo promedio (48 h) de presentación de estros citado en la literatura (Martínez et al. 2007). Sin embargo, el mayor porcentaje de fertilidad, se presentó en las ovejas de T1, que presentaron estro de manera más dispersa entre las 24–48 h posteriores del retiro del CIDR.
Martínez et al. (2007) obtuvieron el 80% de fertilidad al sincronizar ovejas de la raza Blackbelly, con acetato de medroxiprogesterona (MPA) y eCG manejadas en condiciones tropicales. Estos resultados difieren a los obtenidos en este estudio (53.3 %), mientras que en el porcentaje de prolificidad obtuvieron el 108% fue similar al presente estudio (100.8 %). Por otro lado, Martínez et al. (2006) obtuvieron resultados similares en fertilidad (42.2%) y similares en prolificidad (111.1 %) al sincronizar ovejas F1 (Damara X Merino) con progesterona natural y eCG inseminadas intrauterinamente con laparoscopía en condiciones tropicales.
Las concentraciones de progesterona determinadas por RIA, indicaron que todas las ovejas que se seleccionaron para detectar los niveles de progesterona en sangre, estaban ciclando al inicio del experimento, ya que se obtuvieron valores mayores de 2 ng/ml de sangre.
En la literatura científica no se encontraron datos de sincronización de estros en ovejas Damara y F1 (Damara x Merino), por lo cual no es posible establecer una comparación con el mismo tipo de animales y bajo condiciones similares. Esta cruza de ovejas aún cuando son de pelo y se han desarrollado en países tropicales, presentan lana en dos tercios de su cuerpo, por lo que se podría inferir que esto provoque estrés calórico por la alta cantidad de humedad ambiental existente en la zona de estudio, provocando un efecto negativo en su respuesta reproductiva. Esto coincidió con Chemineau (1993), quien mencionó que tanto en las ovejas como en las cabras, expuestas a estrés calórico durante la época reproductiva, pueden producir reducciones drásticas en la tasa de fertilidad y prolificidad.
Boggio (1997) registró que la utilización de GnRH en ovinos tiene como objetivo inducir la ovulación, dato que coincide con los resultados de este estudio, ya que a pesar de que no existieron diferencias estadísticas entre tratamientos, numéricamente las ovejas a las que se aplicó GnRH (T2 y T3) presentaron estros mas agrupados que las del grupo testigo. La disminución en la respuesta a la aplicación de GnRH, probablemente se debió a que se aplicó en una dosis única, por lo que no se estimuló el mantenimiento de los receptores a esta hormona, dato que concuerda con lo reportado por Sakurai et al. (1997), en donde mencionaron que se requieren aplicaciones continuas del factor liberador de gonadotropinas para que haya una respuesta a la aplicación de GnRH.
Nagatani et al. (1998) observaron que diferentes estímulos ambientales y la nutrición son considerados entre los factores más importantes para regular la función reproductiva de los animales, por lo que posiblemente los porcentajes de fertilidad obtenidos en el experimento, aún cuando no son bajos, pudieron haber estado influidos por el genotipo de las ovejas en estudio y su interacción con el ambiente en que fueron manejadas, ya que como se mencionó anteriormente fueron importadas de Australia. Así mismo, Hopkins et al. (1980) detectaron que las ovejas mantenidas en temperaturas ambientales altas, presentaron una disminución en su comportamiento reproductivo y productivo.
Por otro lado, diferentes autores (Bartlewski et al. 2001; Deligiannis et al. 2005; Reyna et al. 2005) mencionaron que la hormona GnRH y la PGF2α al ser utilizadas en combinación con progesterona ó progestágenos, sincronizan e incrementan la tasa de ovulación (Wolfenson et al. 1994; Pursley et al. 1997; Twagiramungu et al. 1995), dato que no concuerda con la presente investigación, ya que el bajo porcentaje de prolificidad indica que existió una baja tasa de ovulación en las ovejas evaluadas.
Las concentraciones de progesterona determinadas por RIA indicaron que todas las ovejas muestreadas ciclaron al inicio del experimento, ya que se obtuvieron valores mayores de 2 ng/ml de sangre, lo que indicó que los animales fueron capaces de responder a un tratamiento para sincronizar estro.
Por los resultados obtenidos en el presente estudio se concluye que el uso combinado de un CIDR impregnado con 0.3 g de progesterona durante 12 días más 25 μg de GnRH, 48 h antes y al momento del retiro del dispositivo no tiene efecto sobre el comportamiento reproductivo en ovejas F1 (Damara x Merino).
LITERATURA CITADA
Akif C, Kuran M (2003) Effects of a single injection of hCG or GnRH agonist on day 12 postmating on fetal growth and reproductive performance of sheep. Vet. Méx. 80(1–2): 81–90. [ Links ]
Álvarez RL, Zarco QLA (2001) Los fenómenos de bioestimulación sexual en ovejas y cabras. Vet. Méx. 32(2):117–129. [ Links ]
Amiridis GS, Valasi I, Menegatos I, Rekkas C, Goulas P, Papanikolaou T, Deligiannis C (2005) Luteal stage dependence of pituitary response to gonadotrophin releasing hormone in cyclic dairy ewes subjected to synchronization of ovulation. Reprod. Fert. Develop. 17(8): 769–774. [ Links ]
Anonymous (1999) User's Guide: Statistics. SAS Institute Inc. Version 8. Cary. 3848 pp. [ Links ]
Bartlewski PM, Beard AP, Chapman. CL, Nelson ML, Palmer B, Aravindakshan J, Cook SJ, Rawilngs NC (2001) Ovarian responses in gonadotrophin–releasing hormone–treated anoestrous ewes: follicular and endocrine correlates with luteal outcome. Reprod. Fert. Dev. 13(2–3): 133–142. [ Links ]
Boggio DJC (1997) Sobrevivencia in vitro de embriones ovinos congelados convencionalmente preparados para transferencia directa en 1.5 M–etilenglicol. Prod. Ovina 10: 63–74. [ Links ]
Chemineau P (1993) Environment and animal reproduction. World Anim. Rev. 77: 2–14. [ Links ]
Del Amo S, Soto M, Cendrero L, Hidding L., Lagunas E (1991) Atlas climatológico del municipio de Córdoba. Serie Estudios Climatológicos No. 5. Instituto de Ecología., A.C. Xalapa. 73 pp. [ Links ]
Deligiannis C, Valasi I, Rekkas CA, Goulas P, Theodosiadou E, Lainas T, Amiridis GS (2005) Synchronization of Ovulation and Fixed Time Intrauterine Insemination in Ewes. Reprod. Domestic Anim. 40(1): 6–10. [ Links ]
Fleet MR, Bennie MJ (2002) A comparison of Damara crossbred and Merino lambs wool. Austral. Anim. Prod. 24: 69–72. [ Links ]
González RG, Vázquez M, Duarte A, González RA (2000) Efecto del morueco y la época de empadre sobre el comportamiento reproductivo en ovejas Pelibuey y Blackbelly. En: XXVIII Reunión Anual de la Asociación Mexicana de Producción Animal; Tapachula, Chiapas, México. 150 pp. [ Links ]
Gutiérrez AJC, Palomares NR, Sandoval MJ, De Ondíz SA, Portillo MG, Soto BE (2005) Uso del protocolo ovsynch en el control del anestro postparto en vacas mestizas de doble propósito. Rev. Cient. FCV–LUZ, 15(1): 7–13. [ Links ]
Hernández CJ, Valencia MJ, Zarco QLA (2001) Regresión del cuerpo lúteo y presentación del estro en ovejas con dos inyecciones de prostaglandina con 8 días de intervalo. Téc. Pec. Méx. 39(1): 53–58. [ Links ]
Hopkins PS, Nolan CJ, Pepper PM (1980) The effects of heat stress on the development of the foetal lamb. Aust. J. Agric. Res. 31: 763–771. [ Links ]
Kridli RT, Hussein MQ, Humphrey WD (2003) Effect of royal jelly and GnRH on the estrus synchronization and pregnancy rate in ewes using intravaginal sponges. Small Ruminant Res. 49(1): 25–30. [ Links ]
Martínez TJJ, Izaguirre FF, Sánchez OL, Castillo GCG, Martínez PG, Torres HG (2007) Comportamiento reproductivo de ovejas barbados barriga negra sincronizadas con mpa y diferentes tiempos de aplicación de eCG durante la época de baja fertilidad. Revista Científica, FCV–LUZ, 17(1): 47–52. [ Links ]
Martínez TJJ, Sánchez–Torres EMT, Bucio AL, Rojo RR, Mendoza MGD, Cordero MJL, Mejía VO (2006) Efecto de eCG e inseminación laparoscópica sobre el comportamiento reproductivo en ovejas F1 (Damara X Merino). Revista Científica, FCV–LUZ, 16(1): 72–77. [ Links ]
Nagatani S, Guthikonda P, Thompson RC, Tsukamura H, Maeda KI, Foster D (1998) Evidence for GnRH regulation by Leptin: Leptin administration prevents reduced pulsatile LH secretion during fasting. Neu–roendocrinology 67: 370–376. [ Links ]
Pursley JR, Wiltbank MC, Stevenson JS, Ottobre JS, Garverick HA (1997) Pregnancy rates per artificial insemination for cows and heifers inseminated at synchronization of ovulation. J. Dairy Sci. 80: 295–300. [ Links ]
Reyna J, Thomson P, Evans G, Maxwell C, (2005) Synchronization of ovulation in Merino ewes with GnRH in the breeding and non–breeding season. Reprod. Fert. Dev. 17(2): 320–320. [ Links ]
Roelofs JB, Van Eerdenburg FJCM, Hazeleger W, Soede NM, Kemp B (2006) Relationship between progesterone concentrations in milk and blood and time of ovulation in dairy cattle. Anim. Reprod. Sci. 91(3–4): 337–343. [ Links ]
Sakurai H, Adams BM, Adams TE (1997) Concentration of gonadotropin releasing hormone receptor messenger ribonucleic acid in pituitary tissue of orchidectomized sheep: Effect of passive immunization against gonadotropin releasing hormone. J. Anim. Sci. 75: 189–194. [ Links ]
Sepúlveda N, Risopatrón J, Rodríguez F, Rodero E (2003) Fertilidad en vacas lecheras asociada a la sincronización de celos e inseminación a tiempo fijo utilizando GnRH y PGF2α. Rev. Cient. FCV–LUZ, 13(3):182–186. [ Links ]
Simonetti L, Ramos G, Gardón JC (1999) Estrus presentation and distribution in ewes treated with intra–vaginal sponges impregnated with medroxiprogesterone acetate (MAP) in combination with pregnant mare serum gonadotropin (PMSG). Braz. J. Vet. Res. Anim. Sci. 36(5): 102–117. [ Links ]
Srinkandakumar ARHI, Ellanworth M, Archabald LF, Liao A, Godke RA (1986) Comparison of a solid phase, no extraction radioimmunoassay for progesterone with an extraction assay for monitoring luteal function in the mare, bitch and cow. Theriogenology 26: 779–793. [ Links ]
Steel GR, Torrie HJ (1981). Principles and Procedures of Statistics a Biometrical Approach. Second (Ed.). McGraw Hill. NY. 633 pp. [ Links ]
Twagiramungu H, Guilbault LA, Deufour JJ (1995) Synchronization of ovarian follicular waves with a gonodotropin–releasing hormone agonist to increase the precision of estrus in cattle: A review. J. Anim. Sci. 73: 3141–3151. [ Links ]
Vincent JN, McQuown EC, Notter DR (2000) Duration of the seasonal anestrus in sheep selected for fertility in a fall–lambing system. J. Anim. Sci. 78: 1149–1154. [ Links ]
Wolfenson D, Thatcher WW, Savio JD, Badinga L, Lucy MC, (1994) The effect of a GnRH analogue on the dynamics of follicular development and synchronization of estrus in lactating cyclic dairy cows. Theriogenology, 42(4): 633–644. [ Links ]

