










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de análisis de la conducta
Print version ISSN 0185-4534
Rev. mex. anál. conducta vol.37 n.1 México Jan. 2011
https://doi.org/10.5514/rmac.v37.i1.24685
Artículos empíricos
Substitución asimétrica entre metanfetamina y anfetamina: Estudio de discriminación de drogas
Asymmetric substitution between methamphetamine and amphetamine: Study of drug discrimination
Florencio Miranda*, Benita Cedillo–Ildefonso, Juan C. Jiménez, Gloria Bedolla–Núñez y Sandra Torres–Rodríguez
FES Iztacala Universidad Nacional Autónoma de México.
La correspondencia debe dirigirse a
Florencio Miranda, Av. de los Barrios 1,
Los Reyes Iztacala Tlalnepantla,
Edo. de México 54090, México,
correo: fmirandah@yahoo.com.
Recibido: Julio 12, 2010
Revisado: Noviembre 13, 2010
Aceptado: Febrero 25, 2011
Resumen
La metanfetamina (MET) es una forma estructural y funcionalmente similar a la anfetamina (ANF). Sin embargo, la MET difiere de la ANF en que a dosis comparables, entran al cerebro niveles más altos de MET, haciéndola un psicoestimulante más potente que la ANF. A pesar de esto, se ha reportado que tanto la MET como la ANF producen efectos conductuales similares. Los procedimientos de discriminación de drogas son herramientas conductuales que se utilizan para evaluar si dos drogas producen efectos conductuales similares o diferentes. Este estudio, evaluó la capacidad de varias dosis de ANF para substituir dosis equivalentes de MET en ratas entrenadas a discriminar MET de salina utilizando la aversión condicionada al sabor como procedimiento de discriminación de drogas. Después de que las ratas aprendieron la discriminación MET–salina, se evaluaron en pruebas de substitución varias dosis de MET y ANF. Los resultados mostraron que las diferentes dosis de MET que se evaluaron produjeron una substitución dependiente de la dosis. Sin embargo, las diferentes dosis de ANF produjeron una substitución parcial dependiente de la dosis, utilizando dosis equivalentes de MET. Estos resultados sugieren que la MET produce efectos conductuales más altos que la ANF en un procedimiento de discriminación de drogas.
Palabras clave: Metanfetamina, anfetamina, discriminación de drogas, ratas.
Abstract
Methamphetamine (METH) is a form of amphetamine (AMPH) that is structurally and functionally similar. However METH differs from AMPH in that, at comparable doses, much higher levels of METH get into the brain, making it a more potent psychostimulant than AMPH. Despite of this, it has been reported that both METH and AMPH produce similar behavioral effects. Drug discrimination procedures are behavioral tools used to assess if two drugs produce similar or different behavioral effects. In the present study we evaluated the ability of various doses of AMPH to substitute for equivalent doses of METH in rats trained to discriminate METH from saline using a conditioned taste aversion as the drug discriminating procedure. After rats learned the discrimination METH–saline, various doses of METH or AMPH were evaluated on substitution tests. The results showed that different doses of METH produced a dose–dependent substitution. However, different doses of AMPH produced a partial dose–dependent substitution using equivalent doses of METH. These results suggest that METH produces higher behavioral effects than AMPH on a drug discrimination procedure.
Key words: Methamphetamine, amphetamine, drug discrimination, rats.
Las drogas tipo anfetamina (ANF) son parte de una clase de drogas llamadas psicoestimulantes, las cuales pueden alterar profundamente las funciones corporales y cerebrales. Estas drogas producen sentimientos de euforia, aumento del estado de alerta, excitación y disminución del apetito (Murray, 1998). El uso crónico de estas drogas, principalmente de la metanfetamina (MET), un derivado N–metilado de la ANF, ha alcanzado proporciones epidémicas en diferentes ciudades del mundo (Meredith, Jaffe, Ang–Lee, & Saxon, 2005), incluyendo México (García–Aurrecoechea, Mejía–Cobarrubias, Díaz–Negrete, & Balanzario–Lorenzana, 1999). Sin embargo, hasta el momento aún no hay un tratamiento efectivo para prevenir o tratar la adicción a la MET. Se necesita más investigación sobre los mecanismos neurobiológicos de la adicción a la MET antes de sugerir un programa preventivo o de tratamiento farmacológico efectivo. No obstante, también se puede utilizar una estrategia complementaria comparando los efectos conductuales de la MET con un psicoestimulante prototipo que tenga una estructura y función similar, tal como la ANF.
La ANF y la MET son drogas psicoestimulantes que comparten una estructura química muy similar y tienen propiedades farmacocinéticas similares (Melega, Williams, Schmitz, & DiStefeano, 1995). También comparten el mecanismo neurobiológico que produce un aumento en la concentración de dopamina (DA) en el núcleo accumbens (Rothman & Baumann, 2003). Ambas drogas actúan sobre el transportador de las vesículas presinápticas para promover un aumento en la concentración citosólica de la DA, y al mismo tiempo invierten la función del trasportador de la recaptura de la DA (DAT por sus siglas en inglés) facilitando su flujo al espacio intersináptico (Rothman & Baumann, 2003).
A pesar de estas similitudes, se ha reportado que hay algunas diferencias entre la MET y la ANF. Es aceptado comúnmente que la MET es más adictiva que la ANF y ha sido considerada como una forma potente y altamente adictiva de la ANF por NIDA (Nacional Institute of drug Abuse Research Report, 2006). También se ha reportado que la MET tiene una vida media mayor que otros psicoestimulantes, y como consecuencia, produce efectos conductuales de mayor duración, de 8–13 horas para MET, 2–4 horas para ANF y 1–3 horas para cocaína. Adicionalmente, la MET tiene una alta solubilidad en lípidos, permitiendo una transferencia más rápida de la droga a través de la barrera hematoencefálica (Barr, Panenka, MacEwan, Thornton, Lang, Honer, & Lecompte, 2006). A pesar de esto, se ha reportado que monos de la India se autoadministran la MET y la ANF a tasas comparables (Balster & Schuster, 1973) y ambos psicoestimulantes producen conductas estereotipadas similares (Kuczenski, Segal, Cho, & Melega, 1995). Un modelo conductual que podría aclarar sí la MET produce efectos conductuales más fuertes que la ANF es el procedimiento de discriminación de drogas (Miranda & Velázquez, 1998). En este modelo, los animales se entrenan a discriminar la droga A de salina y en pruebas de substitución o generalización se substituye la dosis de entrenamiento de la droga A por diferentes dosis de la droga B o de prueba. Así, los resultados de las pruebas de substitución podrían proporcionar información adicional acerca de si la MET es más potente que la ANF.
En el presente estudio se evaluó si los animales entrenados a discriminar 1.0 mg/kg de MET de salina producen una substitución igual o menor con una dosis equivalente de ANF. Adicionalmente, también se evaluó si animales entrenados a discriminar 1.0 mg/kg de ANF producen una substitución similar con una dosis equivalente de MET.
Método
Sujetos
Se utilizaron 20 ratas machos de la cepa Wistar de aproximadamente 120 días de edad al inicio de la investigación, cuyo peso promedio fue de 200 a 250 g, provenientes del Bioterio General de la FES–Iztacala. Las ratas se alojaron individualmente en cajas–hogar de acero inoxidable y bajo un ciclo luz–oscuridad (luz: 8:00 am–8:00 pm) y a una temperatura ambiente de 23 (±1) grados centígrados. La comida siempre estuvo disponible.
Procedimiento
Las ratas fueron privadas de agua y entrenadas a tomar agua por 20 min al día durante 7 días consecutivos. Posteriormente fueron divididas en dos grupos (n=10), el grupo MET (gMET) y el grupo ANF (gANF) y entrenadas por 2 días a tomar una solución de sacarina durante 10 min al día.
Entrenamiento de discriminación de drogas
Las ratas fueron entrenadas a discriminar MET (gMET) o ANF (gANF) de salina con el procedimiento del condicionamiento de aversión a los sabores (CAS). El procedimiento para adquirir la discriminación constó de dos tipos de ensayos:
Ensayos droga.
A las ratas del grupo gMET se les administró 1.0 mg/kg de MET y a las ratas del grupo gANF se les administró 1.0 mg/kg de ANF y 30 minutos después se les permitió el acceso a la sacarina durante 10 minutos. Después de finalizar este período, se les administró a ambos grupos 2.0 ml/kg de 0.17 M de LiCl.
Ensayos salina.
A las ratas de ambos grupos se les administró salina y 30 minutos después se les permitió el acceso a la sacarina durante 10 minutos. Después de finalizar este período, se les administró 1.0 ml/kg de salina.
Entre los ensayos droga y los ensayos salina hubo dos días de descanso, donde se les permitió el acceso al agua simple durante 30 minutos en las cajas–hogar. El ciclo ensayo droga–ensayo salina se repitió 9 ocasiones en un orden aleatorio, con la restricción de que no tuvieran lugar más de dos ensayos droga consecutivos.
Pruebas de substitución.
Los ensayos en estas pruebas se hicieron sobre un ciclo de 4 días. En el día 1, las ratas se sometieron a un procedimiento similar al que se sometieron en el ensayo droga. En el día 2, a los sujetos se les permitió consumir agua simple durante 30 minutos en sus cajas–hogar. El día 3 fue idéntico a los ensayos salina. El día 4 fue de prueba, la cual consistió en administrar una dosis de la droga de prueba a ambos grupos (diferentes dosis de MET o diferentes dosis de ANF). El procedimiento fue similar al utilizado en el día 1 con excepción de que se utilizó una prueba de dos botellas y no se administró el LiCl. Esta prueba consistió en permitirles el acceso durante 10 minutos a dos botellas, una con agua simple y otra con sacarina. Cada dosis evaluada se administró 30 min antes del acceso a las dos botellas.
Drogas
Las drogas que se utilizaron en esta investigación fueron sulfato de d–anfetamina y clorhidrato de metanfetamina (Sigma, St. Louis, MO, USA). Todas las dosis de las drogas se calcularon de acuerdo al peso de la sal y se administraron intraperitonealmente en un volumen de 1.0 ml/kg excepto el LiCl que se administró a una dosis de 2.0 ml/kg de 0.17 M. La solución con sabor fue sacarina (Elly Lilly, México) al 0.15% (p/v) en agua destilada. La MET y la ANF se administraron 30 min antes de iniciar las sesiones experimentales.
Análisis estadístico
Se registró el consumo de sacarina en los ensayos droga y los ensayos salina y se analizó con un ANOVA factorial de dos factores, con los tipos de ensayos (ensayo droga–ensayo salina) como primer factor y el número de ensayo (se analizaron los últimos tres ensayos de cada condición) como segundo factor. Se registró también el consumo de líquidos en las pruebas de substitución. A partir de estos datos se calculó un índice de aversión a la sacarina el cual se obtuvo con la formula A/A+B; donde A fue el consumo de sacarina y B fue el consumo de agua. Con este índice, un valor de 1.0 indicó preferencia por la sacarina y un valor de 0.0 una aversión por la sacarina. Estos datos se analizaron usando un ANOVA de una vía; cuando los ANOVAs fueron significativos, se llevó a cabo un análisis de comparaciones posteriores con la prueba Newman Keuls. En todas las pruebas, el nivel de rechazo del error tipo I fue de 0.05.
Resultados
Adquisición de la discriminación droga–salina
Las ratas de ambos grupos aprendieron la discriminación entre el psicoestimulante y salina (Figura 1). En el grupo gMET, la comparación del consumo de sacarina entre la línea base, el primer ensayo droga y el primer ensayo salina mostró que no hubo diferencias significativas (F[2,27]=0.942, p=0.402). Sin embargo, cuando la administración de MET fue seguida por los apareamientos sacarina–LiCl, se observó una reducción en el consumo de la solución de sacarina. Un ANOVA factorial de dos vías reveló diferencias significativas en el consumo de sacarina entre los últimos 3 ensayos droga y los últimos 3 ensayos salina (condición tipo de ensayo; F[1,54]=112.422, p=0.00001). Los efectos del número de ensayo (F[2,54]=0.258, p=0.774) y la interacción tipo de ensayo y número de ensayo (F[2,54]=0.573, p=0.567) no fueron significativos. Resultados similares se obtuvieron en el grupo gANF. La comparación del consumo de sacarina durante la línea base, el primer ensayo droga y el primer ensayo salina no mostró diferencias significativas (F[2,27]=0.462, p=0.635). El consumo de sacarina durante los últimos 3 ensayos droga fue diferente del consumo de sacarina durante los últimos 3 ensayos salina (F[1,54]=77.743, p=0.0001) y los efectos del número de ensayo (F[2,54]=0.747, p=0.479) y la interacción tipo y número de ensayo (F[2,54]0.253, p=0.778) no mostraron diferencias significativas.

Pruebas de substitución
La Figura 2 muestra los resultados de las pruebas de substitución con MET y ANF en los grupos gMET y gANF. En el caso del grupo gMET la administración de diferentes dosis de MET evaluadas en la prueba de dos botellas produjo una substitución dependiente de la dosis de MET (gráfico A). La evaluación de la dosis de 1.0 mg/kg de MET replicó el control discriminativo de la dosis de entrenamiento de la MET. Un ANOVA de una vía reveló diferencias significativas (F[4,45]=10.381, p=0.0001) y las comparaciones posteriores con la prueba Newman Keuls revelaron que los tratamientos con salina, 0.1 y 0.3 mg/kg de MET fueron significativamente diferentes de la dosis de entrenamiento de MET. La administración de diferentes dosis de ANF en animales entrenados a discriminar MET de salina también produjo una substitución dependiente de la dosis de ANF (gráfico B). Sin embargo, fue necesario aumentar la dosis de la ANF (1.3 mg/kg) para observar un control similar al que produjo la dosis de entrenamiento de la MET. Un ANOVA de una vía reveló diferencias significativas (F[5,54]=9.128, p=0.0001) y las comparaciones posteriores revelaron que las condiciones de salina, 0.1, 0.3 y 1.0 mg/kg de ANF fueron significativamente diferentes de la dosis de entrenamiento de MET.
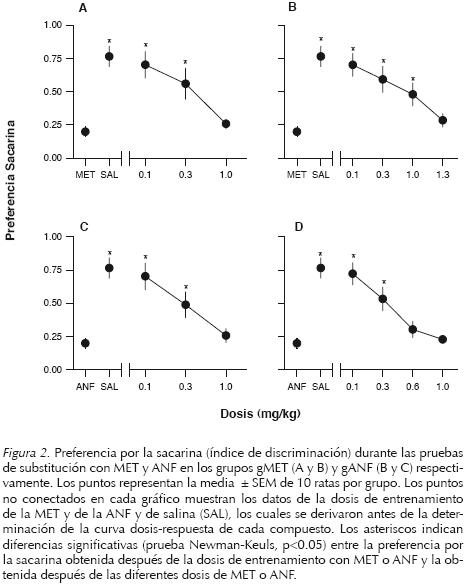
En el caso del grupo gANF la administración de diferentes dosis de ANF también produjo una substitución dependiente de la dosis de ANF (gráfico C). Un ANOVA de una vía reveló diferencias significativas (F[4,45]=10.204, p=0.0001) y las comparaciones posteriores mostraron que las condiciones de salina, 0.1 y 0.3 mg/kg de ANF fueron significativamente diferentes de la dosis de entrenamiento de la ANF. Sin embargo, una dosis relativamente menor de MET (0.6 mg/kg) produjo un control discriminativo similar al producido por la dosis de entrenamiento de ANF (gráfico D). Un ANOVA de una vía mostró diferencias significativas (F[5.54]=14.453, p=0.0001) y las comparaciones posteriores revelaron que las condiciones de salina, 0.1 y 0.3 mg/kg de MET fueron significativamente diferentes de la dosis de entrenamiento de ANF.
El consumo total de líquidos durante las pruebas de substitución (datos no mostrados) no fue alterado en el grupo gMET durante la administración de MET (F[4,45]=0.624, p=0.648) y ANF (F[5,54]=1.51, p=0.345). Tampoco se alteró el consumo total en el grupo gANF durante la administración de ANF (F[4,45]=0.456, p=0.768) y MET (F[5,54]=0.557, p=0.732).
Discusión
El propósito del presente estudio fue examinar si la MET es relativamente más potente que la ANF. Se encontró que la administración de MET o ANF puede inducir un control discriminativo sobre el consumo de sacarina en un procedimiento de discriminación de drogas utilizando el CAS. Este control fue evidente en dos situaciones. La primera de ellas, fue durante el aprendizaje de discriminación MET–salina (grupo gMET) o ANF–salina (grupo gANF). Cuando la administración de la droga de entrenamiento precedió los apareamientos sacarina–LiCl, las ratas disminuyeron el consumo de sacarina. Sin embargo, el consumo de sacarina aumentó o se mantuvo al nivel basal, cuando la administración de salina precedió los apareamientos sacarina–salina. La segunda situación que muestra el control discriminativo de la droga de entrenamiento fue durante las pruebas de substitución con diferentes dosis de MET (en grupo gMET) o ANF (en grupo gANF), donde las ratas mostraron un control discriminativo sobre el consumo de sacarina. Este control fue una función directa de las dosis de prueba de MET o ANF (Figura 2). A mayor similitud entre la dosis de prueba y la dosis de entrenamiento, menor preferencia por la sacarina. Estos datos son similares a los de estudios previos utilizando como droga de entrenamiento a la ANF o MET con procedimientos de discriminación de drogas utilizando el CAS (Miranda, Hermosillo, Sánchez, & Velázquez–Martínez, 2005; Miranda, Sandoval–Sánchez, Cedillo, Jiménez, Millán–Mejía, & Velázquez–Martínez, 2007; Miranda, Jiménez, Cedillo, Sandoval–Sánchez, Millán–Mejía, Sánchez–Castillo, & Velázquez–Martínez, 2009) o con procedimientos operantes de dos palancas (Ho & Huang, 1975; Munzar, Kutkat, Millar, & Goldberg, 2000).
También se encontró en las pruebas de substitución o generalización que en ratas entrenadas a discriminar MET de salina (grupo gMET), la administración de ANF a una dosis equivalente de MET produjo una substitución parcial. El aumento a una dosis mayor de ANF produjo una substitución total. En contraste, en ratas entrenadas a discriminar ANF de salina (grupo gANF), la administración de una dosis relativamente más pequeña de MET produjo una substitución total. Estos resultados sugieren que la MET es más potente que la ANF, al menos, en los efectos discriminativos que ambas drogas producen utilizando el CAS como técnica de entrenamiento discriminativo. Los efectos discriminativos de las drogas adictivas en animales de laboratorio tienen una buena correspondencia con sus efectos subjetivos en humanos y se han utilizado para predecir la probabilidad de que una droga tenga propiedades adictivas (Carter & Griffiths, 2009). De esta forma los datos del presente experimento coinciden con sugerencias de que la MET es más potente que la ANF (Nacional Institute of drug Abuse Research Report, 2006). En contraste a los resultados del presente experimento, otros han reportado que no hay diferencias entre las dos drogas a dosis similares utilizando procedimientos de discriminación de drogas con técnicas operantes de dos palancas (Kuhn, Appel, & Greenberg, 1974). Aunque la razón de esta discrepancia no es clara, se podría sugerir que el procedimiento para establecer la discriminación usando los efectos de las drogas como estímulos discriminativos es un factor importante. Mientras que los estudios con técnicas operantes reportan un fracaso en detectar diferencias entre la MET y la ANF, en el presente estudio se utilizó el procedimiento del CAS. Algunos investigadores han sugerido que el entrenamiento de discriminación de drogas con CAS es más sensible que el utilizado con técnicas operantes (Davis, Stevenson, Cañadas, Ullrich, Rice, & Riley, 2009). Además, es más rápido y puede evitar concentraciones toxicas de las drogas utilizadas para el aprendizaje de la discriminación (Quertemont, 2003).
Los resultados de la presente investigación son consistentes con reportes previos que demostraron que la MET es más potente que la ANF en otros modelos de conducta. Por ejemplo, algunos estudios han reportado que la administración de dosis bajas de MET (0.5 mg/kg) es más potente que la administración de una dosis equivalente de ANF para inducir locomoción (Hall, Stanis, Marquez, & Gulley, 2008). También se ha reportado que la MET produce prolongadas respuestas post estereotipadas comparada con dosis equivalentes de ANF después de someterlas a un modelo de exposición "binge" (Segal & Kuczenski, 1997).
Diversos factores podrían participar en la potencia de la MET sobre la ANF. Por ejemplo, la MET tiene una vida media mayor en el sistema nervioso central que otros psicoestimulantes y debido a su naturaleza lipofílica , la MET penetra más rápido al sistema nervioso central y es más potente que la ANF (Barr et al., 2006). Otro factor que podría contribuir a la mayor potencia de la MET en comparación con la ANF es el efecto de la MET sobre el DAT. El DAT es el blanco principal para los psicoestimulantes como la MET y la ANF y es el mecanismo más importante para la eliminación de la DA del espacio intersináptico, por lo tanto regula la fuerza y duración de la señalización de la DA. Un estudio reciente demostró que en un sistema in vitro, las corrientes celulares mediadas por el DAT fueron mayores con la estimulación de MET que con la estimulación de ANF y que al mismo voltaje y la misma concentración, la MET liberó cinco veces más DA que la ANF. El mismo estudio reportó que la administración sistémica de MET inhibió la eliminación de de DA regulada por DAT en una forma más eficiente que la que produce la ANF (Goodwin, Larson, Swant, Sen, Javitch, Zahniser, DeFelice, & Khoshbouei, 2009). Estos datos sugieren que la MET tiene efectos más fuertes que la ANF en la fisiología celular modulada por el DAT y que esto podría contribuir a efectos más adictivos de la MET.
En conclusión, los resultados de esta investigación indican que tanto la MET como la ANF son capaces de adquirir propiedades discriminativas en un procedimiento de discriminación de drogas con el CAS. Además, se mostró que la MET es más potente que la ANF, al menos en los efectos discriminativos de ambas drogas.
Referencias
Barr, A. M., Panenka, W. J., MacEwan, G. W., Thornton, A. E., Lang, D. J., Honer, W. G., & Lecomte, T. (2006). The need for speed: an update on methamphetamine addiction. Journal of Psychiatry and Neuroscience, 31 (5): 301–13. [ Links ]
Balster, R. L., & Schuster, C. R. (1973). A comparison of d–amphetamine, l–amphetamine, and methamphetamine self–administration in rhesus monkeys. Pharmacology, Biochemistry & Behavior, 1, 67—71; available via: http://dx.doi.org/10.1016/0091–3057(73)90057–9. [ Links ]
Carter, L. P., & Griffiths, R. R. (2009). Principles of laboratory assessment of drug abuse liability and implications for clinical development. Drug and Alcohol dependence, 105 (1), s14–s25; available via: http://dx.doi.org/10.1016/j.drugalcdep.2009.04.003. [ Links ]
Davis, C. M., Stevenson, G. W., Cañadas, F., Ullrich, T., Rice, K. C., & Riley, A. L. (2009). Discriminative stimulus properties of naloxone in Long–Evans rats: assessment with the conditioned taste aversion baseline of drug discrimination learning. Psychopharmacology (Ber), 203. 421–429; available via: http://dx.doi.org/10.1007/s00213–008–1233–5. [ Links ]
García–Aurrecoechea, V. R., Mejía–Cobarrubias, T., Diaz–Negrete, D. B., & Balanzario–Lorenzana, M. C. (1999). Estudio clínico y epidemiológico del consumo de metanfetamina entre pacientes de primer ingreso a tratamiento en la Unidad de Tratamiento y Consulta Externa de Tijuana. CIJ, Informe de investigación, 99–03. [ Links ]
Goodwin, J. S., Larson, G. A., Swant, J., Sen, J., Javitch, J. A., Zahniser, N. R., DeFelice, L. J., & Khoshbouei, H. (2009). Amphetamine and methamphetamine differentially affect dopamine transporters in vitro and in vivo. The Journal of Biological Chemistry, 284 (5), 2978–2989; available via: http://dx.doi.org/10.1074/jbc.M805298200. [ Links ]
Hall, D. A., Stanis, J. J., Marquez, H. A., & Gulley, J. M. (2008). A comparison of amphetamine– and methamphetamine–induced locomotor activity in rats: evidence for qualitative differences in behavior. Psychopharmacology, 195, 469–478; available via: http://dx.doi.org/10.1007/s00213–007–0923–8. [ Links ]
Ho, B. T., & Huang, J. T. (1975). Role of dopamine in D–amphetamine–induced discriminative responding. Pharmacology Biochemistry and Behavior, 3, 1085–1092; available via: http://dx.doi.org/10.1016/0091–3057(75)90021–0. [ Links ]
Kuczenski, R., Segal, D. S., Cho, A. K., & Melega W. P. (1995). Hippocampus norepinephrine, caudate dopamine and serotonin, and behavioral responses to the stereoisomers of amphetamine and methamphetamine. Journal of Neuroscience, 15, 1308—1317. [ Links ]
Kuhn, D. M., Appel, J. B., & Greenberg, I. (1974). An analysis of some discriminative properties of d–amphetamine. Psychopharmacologia, 39, 57–66; available via: http://dx.doi.org/10.1007/BF00421458. [ Links ]
Melega, W., Williams, A., Schmitz, D., DiStefano, E., & Cho, A. (1995). Pharmacokinetic and pharmacodynamic analysis of the actions of d–amphetamine and d–methamphetamineon the dopamine terminal. The Journal of Pharmacology and Experimental Therapeutics, 274, 90—96. [ Links ]
Meredith, C. W., Jaffe, C. J., Ang–Lee, K., & Saxon, A. J. (2005). Implications of chronic methamphetamine use: a literature review. Harvard Review of Psychiatry, 13, 141–154; available via: http://dx.doi.org/10.1080/10673220591003605. [ Links ]
Miranda, F., Hermosillo, A., Sánchez, H., & Velázquez–Martínez, D. N. (2005). Mecanismos no dopaminérgicos en las propiedades discriminativas de la anfetamina: efectos de agonistas serotonérgicos. Revista Mexicana de Psicología, 22(2), 553–559. [ Links ]
Miranda, F., Jiménez, J. C., Cedillo, L. N., Sandoval–Sánchez, A., Millán–Mejía, P., Sánchez–Castillo, H., & Velázquez–Martínez, D. N. (2009). The GABAB antagonist 2 hydroxysaclofen reverses the effects of the baclofen on the discriminative stimulus effects of D–amphetamine in the conditioned taste aversion procedure. Pharmacology Biochemistry and Behavior, 93, 25–30; available via: http://dx.doi.org/10.1016/j.pbb.2009.04.002. [ Links ]
Miranda, F., Sandoval–Sánchez, A., Cedillo, L. N., Jiménez, J. C., Millán–Mejía, P., & Velázquez–Martínez, D. N. (2007). Modulatory role of 5–HT1B receptors in the discriminative signal of amphetamine in the conditioned taste aversion paradigm. Pharmacological Reports; 59, 517–24. [ Links ]
Miranda, F., & Velázquez, D. N. (1998). Control de estímulos con anfetamina utilizando el procedimiento del condicionamiento aversivo a los sabores. Revista Mexicana de Psicología, 15(2), 141–148 [ Links ]
Munzar, P., Kutkat, S. W., Miller, C. R., & Goldberg, S. R. (2000). Failure of baclofen to modulate discriminative–stimulus effects of cocaine or methamphetamine in rats. European Journal of Pharmacology, 408, 169–174; available via: http://dx.doi.org/10.1016/S0014–2999(00)00772–X. [ Links ]
Murray, J. B. (1998). Psychophysiological aspects of amphetamine methamphetamine abuse. The Journal of Psychology, 132, 227–237; available via: http://dx.doi.org/10.1080/00223989809599162. [ Links ]
NIDA Research Report (2006). Methamphetamine abuse and addiction. NIH Publication no. 06–4210: 1—8. [ Links ]
Quertemont, E. (2003). Discriminative stimulus effects of ethanol with a conditioned taste aversion procedure: lack of acetaldehyde substitution. Behavioural Pharmacology, 14, 343–350. [ Links ]
Rothman, R. B., & Baumann, M. H. (2003). Monoamine transporters and psychostimulant drugs. European Journal of Pharmacology, 479, 23–40; available via: http://dx.doi.org/10.1016/j.ejphar.2003.08.054. [ Links ]
Segal, D. S., & Kuczenski, R. (1997). Repeated binge exposures to amphetamine and methamphetamine: behavioral and neurochemistry characterization. The Journal of Pharmacology and Experimental Therapeutics, 282, 561–573. [ Links ]
* La contribución de los autores fue equiparable. Esta investigación fue financiada por PAPIIT–UNAM IN301809 y CONACyT 60872.

