










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de análisis de la conducta
versión impresa ISSN 0185-4534
Rev. mex. anál. conducta vol.36 no.2 México ene. 2010
Artículos de investigación empírica
Estrategias de búsqueda y consumo de alimento en grupos de ratas expuestos a diferentes distribuciones de alimento
Foraging strategies in groups of rats exposed to different arrangement of food
Luis Alfaro*, Óscar García–Leal* y Rosalva Cabrera**1
*CEIC, Universidad de Guadalajara
**FES Iztacala, UNAM — CEIC, Universidad de Guadalajara
Correspondencia:
Centro de Estudios e Investigaciones en Comportamiento,
CEIC, Centro Universitario de Ciencias Biológicas y Agropecuarias,
Universidad de Guadalajara. Francisco de Quevedo 180.
Col. Arcos Vallarta CP 44130 Guadalajara,
Jalisco, México.
Email: luisalfaroh@gmail.com,
oscargl@cencar.udg.mx,
rosalva@unam.mx.
Recibido: Mayo 6, 2010
Revisado: Junio 15, 2010
Aceptado: Junio 29, 2010
Resumen
Este estudio evaluó las estrategias de búsqueda y consumo de alimento en grupos de ratas, variando el número de integrantes por grupo y la distribución espacial de fuentes de alimento. Grupos de ratas fueron expuestos a una tarima de madera con depósitos de alimento. Los grupos evaluados fueron: Matriz n=4, constituido por cuatro integrantes y expuesto a 4 depósitos con alimento cercanos que conformaron una matriz; Matriz n=8, expuesto a la misma distribución de depósitos que el grupo anterior, pero constituido por ocho integrantes; Zig–Zag n=4, expuesto a una distribución irregular de cuatro depósitos con alimento y distantes entre sí, conformado por cuatro integrantes y Zig–Zag n=8, constituido por ocho integrantes y expuesto a la distribución irregular y distante. Los resultados mostraron que la distribución de los depósitos produjo diferencias en la cantidad de depósitos explotados por todos los integrantes del grupo, así como tendencias diferentes en las estrategias producir–parasitar, mientras que el número de integrantes por grupo tuvo efectos en la proporción de sujetos que ejecuta la respuesta de producir y en la regularidad de los patrones de llegada a los depósitos de alimento. Estos resultados se discuten en términos del modelo de maximización de tasa.
Palabras clave: búsqueda de alimento, distribución espacial, tamaño de grupo, estrategias, ratas.
Abstract
This study evaluated searching and intake strategies in groups of rats; group size and arrangement of sources with food were varied. Group of rats were exposed to a wood table with food containers. Group Matrix n=4 integrated by four rats was exposed to 4 containers which were arranged in a matrix figure and were near; Group Matrix n=8 was exposed to same arrangement of containers y was integrated by eight rats; Zig–Zag n=4 was exposed to 4 containers which were away and arranged in irregular figure, four rats were in the group; Zig–Zag n=8 was exposed to same arrangement and was composed with eight rats. Data show that arrangement of sources with food had effects in the number of containers exploted by all rats in the group and in the tendency of producing–scrounging strategies; the number of rats by group produced changes on the proportion in that producing and scrounging responses were performed in each group and on pattern of arrive to sources of food. These results were analyzed according to maximization rate model.
Key words: searching food, spatial arrangement, group size, strategies, rats.
Introducción
En situaciones de búsqueda de alimento, los organismos pueden rastrear el alimento de manera individual, de tal manera que la obtención de recursos alimenticios depende de la habilidad de cada sujeto para localizar, muestrear y explotar fuentes potenciales de alimento; también pueden hacerlo de manera grupal, esto es, cuando los organismos pueden acceder a las fuentes de alimento en función de sus habilidades para descubrirlas y/o a partir de observar a otros individuos y aprovechar los recursos que éstos han descubierto y dejan disponibles (Cabrera, Durán & Nieto, 2006a; 2006b; Dall, Giraldeau, Olsson, McNamara & Stephens, 2005; Hewitson, Gordon & Dumont, 2006). El interés de este estudio es abordar las situaciones de búsqueda de alimento bajo condiciones grupales.
Según Barnand y Sibly (1981), en situaciones grupales de búsqueda de alimento es posible identificar dos tipos de respuestas mutuamente excluyentes para acceder al alimento en un momento determinado: 1) Producir, cuando la emisión de una respuesta permite a un sujeto encontrar y tener acceso a una cantidad de alimento y generalmente, poder consumirlo parcialmente antes que otros integrantes del grupo. 2) Parasitar, cuando un sujeto consume parte del alimento contenido en una fuente al arribar a ella una vez que ha sido liberada por otro sujeto.
Habitualmente, se observa que los sujetos alternan entre producir y parasitar para acceder al alimento (Giraldeau & Lefebvre, 1986; Cabrera, Durán & Nieto, 2006a). Cuando un sujeto a lo largo de diferentes episodios de búsqueda de alimento emite mayor proporción de respuestas de producción en relación a las respuestas de parasitar, se dice que despliega una estrategia de "productor". Por otra parte, cuando un sujeto emite mayor proporción de respuestas de participar del alimento producido por otros en relación a las respuestas de producir, se dice que despliega una estrategia de "parásito" (Barnand & Sibly, 1981; Caraco & Giraldeau, 1991). Dado que los organismos pueden elegir entre estas dos estrategias, resulta interesante preguntarse ¿qué factores determinan que un organismo siga una estrategia específica en un momento particular?
Al respecto, Parker (1984) menciona que la estrategia seguida por un individuo depende en gran medida de las estrategias que están siendo jugadas por otros individuos, de tal manera que cada integrante sigue una estrategia que le permite maximizar el pago neto que recibe al final del episodio de forrajeo. Así, en situaciones de forrajeo grupal existe una interdependencia económica concurrente entre los pagos y costos de diferentes individuos en función de las estrategias jugadas por cada uno (Giraldeau & Caraco, 2000).
El modelo de maximización de la tasa (Vickery, Giraldeau, Templeton, Kramer & Chapman, 1991) plantea que cuando un productor encuentra una fuente de alimento consume parcialmente ésta de manera exclusiva (ventaja de producción), la cual permanece hasta que los parásitos arriban a la fuente y consumen junto con el productor la comida restante, misma que es repartida en porciones similares para el total de sujetos que explotan la fuente, es decir, entre el total de parásitos y el productor (Veáse Giraldeau & Livoreil, 2000).
La proporción de productores es afectada por factores relativos a las características de la población como: tamaño del grupo, entre más grande es el grupo se tiende a incrementar el uso de parasitar como estrategia de búsqueda (Coolen, 2002); nivel de demanda energética, a menor demanda energética se incrementa la cantidad de sujetos que utilizan parasitar como estrategia de búsqueda (Koops & Giraldeau, 1996); experiencia previa en la ejecución de la respuesta de producir, entre más sujetos cuenten con experiencia en la emisión de la respuesta de producción se tiende a disminuir la proporción de sujetos que adoptan parasitar como estrategia para acceder al alimento (Alfaro y Cabrera, 2009; Cabrera, Durán & Nieto, 2006a).
A su vez, la estrategia de parasitar es más eficiente a menor proporción de parásitos en un grupo (Caraco & Giraldeau, 1991); otra ventaja de esta estrategia es que los sujetos tienen un menor desgaste energético, ya que no invierten tiempo en buscar, descubrir y dejar disponible el alimento (Beauchamp, 2008); además, incrementa la probabilidad de acceso al alimento una vez que éste se encuentra disponible, lo cual la hace una estrategia altamente efectiva (Koops & Giraldeau, 1996).
Otros factores que modulan la proporción con que confluyen ambas estrategias en un grupo son los relativos a las características de la parcela, tales como: 1) La distancia entre fuentes de alimento potenciales, la cual puede determinar el tiempo requerido para buscar, descubrir y explotar una fuente alimenticia; también influye en el tiempo de permanencia en una fuente descubierta (Livoreil & Giraldeau, 1997); además implica cambios en el riesgo predatorio (Barrette & Giraldeau, 2008) y conlleva gastos de energía diferentes. 2) Las fuentes de alimento potenciales proporcionadas por la parcela (Koops & Giraldeau, 1996); 3) El tipo de alimento contenido en términos del costo para su manipulación y consumo (Alfaro & Cabrera, 2009) y de la calidad del mismo; 4) La densidad de alimento que contiene cada fuente (Cabrera, Durán & Nieto, 1996 a) y 5) El costo de respuesta para dejar disponible el alimento (Cabrera, Durán & Nieto, 1996 b).
Dado lo anterior, resulta interesante preguntarse ¿Cómo interactúan las características de la población y las características de la parcela en las estrategias de búsqueda y consumo de alimento?
El presente trabajo constituye trata de dar respuesta a esta pregunta y tuvo los siguientes objetivos experimentales: 1) evaluar los efectos de variar el número de integrantes que conforma un grupo sobre las estrategias de búsqueda y consumo de alimento de sus diferentes integrantes y 2) evaluar el efecto de dos distribuciones de alimento, una de ellas con depósitos de alimento cercanos y con una distribución geométrica regular versus otra en la que hay mayor distancia entre depósitos y su configuración es irregular sobre estas estrategias.
Método
Sujetos
Los sujetos fueron 24 ratas Wistar, experimentalmente ingenuas, de aproximadamente cuatro meses al inicio del experimento, provenientes del Bioterio del Centro de Estudios e Investigaciones en Comportamiento de la Universidad de Guadalajara. Todos los sujetos fueron sometidos a un régimen de privación de alimento para mantenerlos al 80% ± 3% de su peso en libre alimentación. Para su identificación, cada sujeto fue marcado con color vegetal marca McCormick en el dorso, los colores fueron rojo, azul, amarillo y verde o combinaciones de dos de estos colores, los sujetos que no fueron marcados son identificados como blancos.
Aparatos
El aparato experimental consistió en una tarima de madera que midió 180 cm de largo por 120 cm de ancho, la cual descanso sobre una base de madera de 20 cm de altura. Sobre cada orilla de la tarima había una pared de policarbonato transparente de 50 cm de altura. En la tarima hubo 12 perforaciones circulares, cuyo diámetro fue de 6.0 cm; la separación mínima entre perforaciones fue de 30cm y la máxima de 150 cm. Debajo de cada perforación se colocó un depósito de plástico cuya profundidad fue de 2 cm. Los depósitos estuvieron cubiertos por una compuerta de madera cuyo peso fue de 48 g, la cual podía ser desplazada para dejar disponible el alimento.
Materiales
Se utilizó una cámara de video, una computadora personal y una consola para reproducción y análisis de datos.
Situación experimental
Los experimentos se llevaron a cabo en un laboratorio de 5 m x 3 m x 3m en cuyo suelo se colocó una tarima. La cámara de video se situó en arnés aéreo, el cual se ubicó a 2.5 m del suelo y sobresaliendo 25 cm de la pared, permitiendo filmar toda la superficie del aparato.
Procedimiento
Los sujetos fueron asignados aleatoriamente a uno de cuatro grupos experimentales; dos grupos estuvieron conformados por 4 sujetos y otros dos grupos por 8 sujetos cada uno. Cada grupo de sujetos fue expuesto al aparato experimental, cuyos depósitos estuvieron con la compuerta cerrada, de tal manera que no podía ser identificado si contenían o no alimento. Las sesiones experimentales finalizaron cuando todos los depósitos con alimento fueron agotados o una vez que transcurrieron 15 minutos. Los sujetos fueron expuestos a esta situación durante 5 sesiones consecutivas, realizándose una sesión por día. Todos los grupos podían encontrar cuatro de los doce depósitos conteniendo alimento. Los diferentes grupos experimentales estuvieron definidos por la distribución espacial de los depósitos con alimento (Ver Figura 1) y el número de sujetos por grupo.

Dos grupos, uno conformado por 4 integrantes (n=4) y otro por 8 integrantes (n=8) fueron expuestos a una condición en la que los cuatro depósitos con alimento estuvieron contiguos y contuvieron 3 g de semillas de girasol por sujeto. De tal manera que la disponibilidad de alimento quedo restringida a una esquina del aparato, formándose un cuadro entre los depósitos con alimento disponible, siendo de 30 cm la distancia entre depósitos, a estos grupos se le denominó Grupos Matriz (ver parte izquierda de Figura 1).
Los Grupos Zig–Zag n=4 y n=8 fueron expuestos a una condición en la que también sólo 4 de los 12 depósitos contuvieron alimento (3 gr por sujeto). En esta condición, dos depósitos conteniendo alimento estuvieron ubicados de forma alternada en una hilera periférica de cuatro depósitos, mientras que los otros dos depósitos se ubicaron de manera también alternada en la otra hilera periférica. Así, entre dos filas consecutivas la distancia entre depósitos con alimento fue de aproximadamente 60 cm (ver parte derecha de Figura 2).
Definición de categorías conductuales
1) Visita a un depósito abierto fue registrada cuando un sujeto introdujo el hocico en el depósito abierto por al menos 3 segundos. 2) Producir fue registrada cuando un sujeto desplazó la compuerta de un depósito de alimento, dejando éste disponible. 3) Parasitar fue registrada cuando un sujeto visita un deposito previamente abierto por otro sujeto y consume parte del alimento disponible.
Registro y Análisis de Datos
Todas las sesiones fueron video–grabadas y a posteriori dos observadores independientes revisaron las sesiones completas y realizaron los registros; éstos fueron considerados para análisis sólo cuando el índice de concordancia entre observadores fue superior a 90%. Los observadores registraron la frecuencia de visitas realizada por cada uno de los sujetos y el tiempo de ocurrencia con base en el inicio de la sesión, lo cual permitió identificar el orden de llegada de los diferentes integrantes de cada grupo para visitar las fuentes de alimento útiles y explotarlas; también se contabilizó la frecuencia de las respuestas de producir y parasitar. Al inicio y al final de cada sesión fue registrado el peso corporal de cada uno de los sujetos.
Resultados
A partir de las videograbaciones, se identificó el orden en el cual los integrantes de cada uno de los grupos arribaron a los depósitos con alimento en cada una de las sesiones experimentales, el registro inició desde el momento en que la compuerta del depósito fue desplazada hasta que el alimento se agotó; lo cual proporcionará información sobre el patrón de respuestas desplegado por cada sujeto en cada una de las sesiones. También se calculó la proporción en que cada una de los sujetos emitió la respuesta de producir alimento en relación a la respuesta de parasitar; este índice proveerá información sobre la tendencia de cada integrante del grupo por seguir una estrategia para obtener y consumir el alimento.
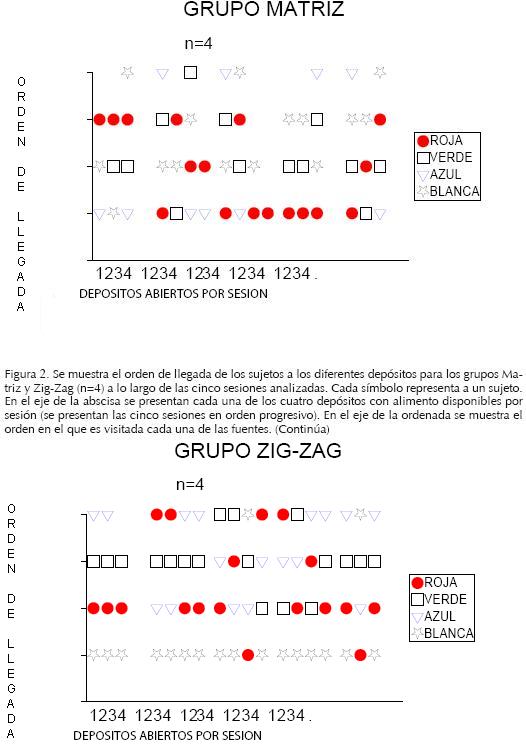
Orden de llegada a depósitos
Los sujetos que arribaron en primer lugar a cada depósito fueron aquellos que ejecutaron la respuesta de abrirlo, de tal manera que el alimento contenido en éste estuviera disponible para ser consumido, es decir, fueron los sujetos productores de alimento (valor 1 en la ordenada de las figuras); los sujetos que visitaron el depósito en segundo lugar, fueron los parásitos de primer nivel, esto es, fueron los sujetos que consumen en primera instancia el alimento que el productor dejó disponible (valor 2); los valores 3 en adelante de la ordenada indican sujetos que parasitaron el alimento a niveles medios o bajos.
En las Figuras 2 y 3 puede apreciarse que los sujetos no explotaron todos los recursos disponibles, ya que sólo en el grupo Matriz n=8 los sujetos abrieron el 100% de los depósitos con alimento, los grupos Matriz n=4 y Zig–Zag n=4 abrieron el 85% de depósitos, mientras que el grupo Zig–Zag n=8 abrió el 75%.
Asimismo, puede verse que los depósitos abiertos no siempre fueron visitados por todos los integrantes del grupo; así, en los grupos Matriz una menor proporción de depósitos abiertos fue visitada por todos los miembros del grupo, el 47% de sujetos para Matriz n=4 y el 20% para Matriz n=8. Por su parte, en los grupos Zig–Zag hubo una mayor proporción de depósitos visitados por todos los integrantes, superando el 90% en ambos grupos.
Además, estos datos permiten identificar que en el Grupo Matriz n=4 todos los integrantes ejecutaron al menos una vez la respuesta para dejar disponible el alimento, mientras que en el Grupo Matriz n=8 sólo el 62% de los sujetos ejecutó esta respuesta; por su parte, en ambos grupos Zig–Zag el 50% de los sujetos ejecutó esta respuesta (ver Figuras 2 y 3).
En cuanto a la consistencia con la cual fue ejecutada la respuesta de producir, la Figura 2 muestra que la proporción de sujetos que emitió esta respuesta un mayor número de veces y a través de varias sesiones fue superior en el grupo Matriz n=4, en este grupo el 50% de los sujetos (rojo y azul) emitió la respuesta en al menos tres sesiones con una frecuencia superior a 6; en el grupo Zig–Zag n=4 sólo el sujeto blanco fue productor consistente, lo cual con un valor relativo de 25%.
En la Figura 3 puede observarse que en el Grupo Matriz n=8 el 25% de los sujetos produjo alimento bajo estos criterios, –sujetos verde y amarillo–; finalmente, en el grupo Zig–Zag n=8 el 12.5% fue productor consistente —el sujeto azul–.
La consistencia de la respuesta de parasitar en primer nivel en el grupo Matriz n=4 fue de 50%, ya que los sujetos verde y blanco parasitaron durante 4 de las 5 sesiones, teniendo una frecuencia de 7 y 6 depósitos parasitados respectivamente. En el grupo Zig–Zag n=4 el 50% de los sujetos fue parasito consistente, pues el sujeto rojo parásito en 5 sesiones con una frecuencia de 10 depósitos y el sujeto azul lo hizo en 3 sesiones con una frecuencia de 5 (ver Figura 2).
El grupo Matriz n=8 tuvo el 38% de parásitos consistentes, ya que los sujetos verde, rojo con amarillo y rojo con azul parasitaron en al menos 3 sesiones y con una frecuencia de 4 depósitos. En el grupo Zig–Zag n=8 sólo el sujeto azul parásito en tres sesiones con una frecuencia de 4 depósitos visitados (ver Figura 3).
Índice Producir–Parasitar
Otra medida sobre las estrategias de búsqueda y consumo de alimento fue el índice producir–parasitar, el cual fue calculado de acuerdo a la siguiente fórmula:
frecuencia de producir / frecuencia de producir + frecuencia de parasitar
Los índices calculados para cada sujeto en cada una de las sesiones se presentan en la Figura 4; aquellos sujetos que en al menos dos sesiones aparecen entre los valores 0.67 y 1.0 son considerados con tendencia a producir, ya que estos valores indican que emitieron en mayor proporción la respuesta de producir que la de parasitar; los sujetos con al menos dos sesiones en los valores entre 0.34 y 0.66 son sujetos que usan ambas estrategias de manera intercambiable, pues a través de las sesiones las usan casi en la misma proporción y por lo mismo, se considera que siguen una estrategia mixta; los sujetos que en al menos dos sesiones aparecen entre los valores 0.0 y 0.33 son considerados con tendencia a parasitar, pues su proporción de respuestas de parasitar fue bastante superior a la de producir.
La Figura 4 muestra que en el Grupo Matriz (n=4) el 50% de los sujetos produjeron y el 50% restante parasitaron; así, los sujetos azul y rojo presentaron tendencia a producir, mientras que los sujetos blanco y verde tuvieron tendencia a parasitar. Por su parte, en los sujetos del Grupo Matriz (n=8) prevaleció la tendencia a parasitar y a usar estrategia mixta, ya que el 62% de los sujetos mostró la tendencia a parasitar de manera casi absoluta y el 38% restante (sujetos verde, amarillo y blanco) uso una estrategia mixta.
En el Grupo Zig–Zag (n=4) sólo el 25% de los sujetos exhibió tendencia a producir –el sujeto blanco–el 75% restante mostró tendencia a parasitar. En lo que concierne al Grupo Zig–Zag (n=8) el sujeto azul fue el único que tuvo tendencia a producir (12.5%), mientras que el 87.5% mostró tendencia a parasitar.
Peso corporal
Como ya se mencionó lo sujetos también fueron pesados al final de la sesión, lo cual permitió identificar el consumo de alimento neto de los integrantes en cada sesión; a partir de esto, se calculó el porcentaje de peso con el que los sujetos finalizaron la sesión.
Esta variable, en primer término nos muestra que en los cuatro grupos evaluados hubo un incremento en el porcentaje de peso ganado por los sujetos conforme transcurrieron las sesiones; lo cual por una parte indica que a lo largo de las sesiones los grupos fueron explotando los recursos de manera más eficiente y por otra, que hubo tal abundancia de recursos que los sujetos lograron incrementar su peso corporal a través de las sesiones experimentales (ver Figuras 5 y 6).
La fígura 5 muestra que en los Grupos Matriz y Zig–Zag n=4 los sujetos que ganaron más peso fueron los parásitos de primer nivel, mientras que en el grupo Matriz el mejor productor –roja– fue quien ganó menor peso y en el grupo Zig–Zag la ganancia para el productor –blanca– fue intermedia en relación a la de los demás miembros de su grupo.
En la Figura 6 se muestra que en el Grupo Matriz n=8 el sujeto rojo–azul fue quien ganó mayor peso, fue parásito de nivel intermedio; mientras que el sujeto amarilla fue quien ganó menos peso, este sujeto fue un buen productor en este grupo. En el Grupo Zig–Zag n=8, nuevamente un sujeto parásito –verde– fue quien ganó mayor peso; el sujeto rojo con verde fue quien ganó menos peso, este sujeto fue parásito de nivel bajo. En ambos grupos, los productores ganaron peso a nivel intermedio.
Discusión
Como puede notarse, los dos indicadores de las estrategias seguidas por los integrantes de un grupo para obtener y consumir alimento reportados previamente, proporcionan información complementaria, ya que el orden de llegada a cada depósito útil permite visualizar de manera general la actividad desplegada por todo el grupo durante cada sesión, permitiendo identificar consistencias de las secuencias seguidas por cada sujeto; mientras que el índice producir–parasitar refleja la propensión de los sujetos a seguir una de la estrategias evaluadas tanto intra sesión como entre sesiones.
El primer indicador mostró que en la situación experimental de este estudio no fueron explotados el total de los recursos disponibles; ya que en lo general, no fueron abiertos todos los depósitos con alimento y los depósitos abiertos no fueron visitados por todos los sujetos, lo cual puede indicar que hubo un superávit de recursos.
También mostró que la distribución espacial de las fuentes de alimento tuvo efectos tanto sobre el nivel de aprovechamiento de los recursos por todos los miembros del grupo como sobre la regularidad de las estrategias seguidas por los sujetos. Con relación al aprovechamiento, en los grupos con una distribución irregular de los recursos en la situación experimental y con una mayor distancia entre depósitos (Zig–Zag n=4 y n=8) hubo una mayor proporción de depósitos visitados por todos los integrantes del grupo, lo cual implica que cuando existe una mayor dificultad para obtener recursos, éstos se aprovechan más eficientemente (Livoreil & Giral–deau, 1997). En los que respecta a la regularidad de las estrategias, en los grupos con mayor distancia entre depósitos uno de los sujetos por grupo siguió de manera consistente la respuesta de producir (p.e. en el grupo Zig–Zag n=4, el sujeto blanco y en el Zig–Zag n=8 el sujeto azul), mientras que en los grupos con depósitos cercanos los sujetos alternan esta respuesta con la de parasitar, tal y como lo demuestra la diferencia en el porcentaje de sujetos que ejecutó la respuesta de producir entre ambas distribuciones de depósitos con alimento.
Los datos obtenidos con el cálculo del índice productor–parásito nuevamente indican un efecto de la distribución espacial de los depósitos, ya que en la condición de depósitos distante los sujetos tienden a mostrar predominantemente sólo una de las estrategias, esto es, producen de manera consistente o parasitan; en tanto que en los grupos expuestos a depósitos cercanos los sujetos juegan con mayor tendencia una estrategia mixta, es decir, alternan entre producir y parasitar
Por su parte, el número de integrantes por grupo también tuvo efectos sobre la proporción de sujetos que siguió la estrategia de producir con relación a la proporción de sujetos que siguió la de parasitar. En los grupos con cuatro integrantes, la proporción de sujetos con tendencia a producir fue superior a la de los grupos con ocho integrantes, lo cual pudo ser función simplemente del incremento de la densidad de sujetos por área (Coolen, 2002; Zamora, López, Andrade, Hernández, Rosas, Zermeño & Cabrera, 2007), lo cual ocasiona que el número de depósitos con alimento per cápita sea menor en los grupos con más integrantes; sin embargo, aun cuando este factor estuvo presente, no puede considerarse que haya sido determinante para este resultado, pues los sujetos fueron expuestos a la situación durante 5 sesiones, es decir, cada sujeto tuvo 20 oportunidades de ejecutar la respuesta de abrir un depósito.
Con relación al peso ganado por los sujetos al final de la sesión, los datos muestran que en lo general la estrategia de parasitar pudo haber sido más rentable que la de producir, puesto que los sujetos que más peso ganaron fueron parásitos. Al respecto, el modelo de maximización de la tasa (Livoreil & Giraldeau, 1997; Vickery et al, 1991) propone que conforme se incrementa el tamaño del grupo la estrategia de parasitar se hace menos rentable con relación a la de producir; sin embargo, dado que la preparación experimental de este estudio mantuvo constante la cantidad de alimento per cápita, los parásitos no tuvieron pérdidas cuando se incrementó la población.
Por otra parte, la estrategia de parasitar pareció ser más eficiente en los grupos con menos integrantes, ya que en éstos los parásitos alcanzan mayor ganancia de peso que en los grupos con más integrantes; lo anterior pudo ser función de que a menor número de integrantes por grupo, mayor proporción de sujetos explotó cada depósito, lo cual hace que las ganancias totales obtenidas sean mayores. Adicionalmente, un mayor número de integrantes por grupo ocasiona que la tasa de consumo momentánea sea menor, ya que incrementa la competencia por acceder a cada depósito abierto.
Ahora bien, parece claro que la estrategia de parasitar otorga ganancias a los sujetos que la siguen, pero ¿cuál es la ganancia de la estrategia de producir? El modelo de maximización de la tasa propone que los productores tienen la ventaja de primacía, es decir, son los primeros en acceder a los recursos y durante un tiempo los explotan de manera exclusiva, lo cual ocasiona que mantengan esta estrategia; las ganancias en peso mostraron que los productores ganan peso a niveles intermedios, lo cual puede indicar que el gasto de energía de la respuesta de búsqueda puede ser alto, pero que el ser consumidores iniciales compensa ese gasto y les permite tener la ingesta necesaria para mantenerse cerca del promedio de consumo de su grupo (Livoreil & Giraldeau, 1997; Vickery et al, 1991).
Para finalizar, es importante mencionar que es poca la diferencia entre los pesos ganados por los diferentes integrantes de los grupos evaluados, lo cual apoya lo propuesto por las teorías de optimización de recursos en grupos animales, en el sentido de que los sujetos se distribuyen los recursos de manera similar (Fretwell & Lucas, 1970) y para ello siguen las mejores estrategias en un episodio particular de búsqueda de alimento (Parker, 1984).
Referencias
Alfaro, L., & Cabrera, R. (2009). Una evaluación de diferentes estrategias productor parásito en grupos de ratas. XIX Congreso Mexicano de Análisis de la Conducta. Aguascalientes, México. [ Links ]
Barnand, C.J., & Sibly, R.M. (1981). Producers and scroungers: a general model and its application to captive focks of house sparrows. Animal Behaviour, 24, 543–550. [ Links ]
Barrette, S., & Giraldeau, L–A. (2008). Evidence against maximization of gross rate of seed delivery to the burrow in food–hoarding eastern chipmunks, tamias striatus. Animal Behaviour, 75, 655–661. [ Links ]
Beauchamp, G. (2008). A spatial model of producing and scrounging. Animal Behaviour, 76, 1935–1942. [ Links ]
Cabrera, R., Durán, A., & Nieto, J. (2006a). Aprendizaje social y estrategias de forrajeo en parvadas de palomas: efectos de la cantidad de alimento. Revista Mexicana de Psicología, 23, 111–121. [ Links ]
Cabrera, R., Durán, A., & Nieto, J. (2006b). Aprendizaje social de respuestas óptimas y estrategias productor–parásito en parvadas de palomas. Psicothema, 18, 724–729. [ Links ]
Caraco,T., & Giraldeau, L.–A. (1991). Social foraging: producing and scrounging in a stochastic environment. Journal of Theorical Biology, 153, 559–583. [ Links ]
Coolen, I. (2002). Increasing foraging group size increases scrounger use and reduces searching efficiency in nutmeg mannikins (Lonchura punctulata). Behavioral Ecology Sociobiology, 52, 232–238. [ Links ]
Dall, S.R.X., Giraldeau, L.–A., Olsson, O., McNamara, J.M., & Stephens, D.W. (2005). Information and its use by animals in evoluctionary ecology. Trends in Ecology and Evolution, 20, 187–193. [ Links ]
Fretwell, S.D., & Lucas, H.L. (1970). On territorial behaviour and other factors influencing habitat distribution in birds. Acta Biotheoretica, 19, 16–36. [ Links ]
Giraldeau, L.–A., & Caraco, T.(2000). Social Foraging Theory. Princeton University Press: New Jersey. [ Links ]
Giraldeau, L.–A., & Lefebvre, L. (1986). Exchangeable producer and scrounger roles in a captive fock of feral pigeons: a case for scrounger skill pool effect. Animal Behaviour, 34, 797–803. [ Links ]
Giraldeau, L.–A., & Livoreil, B. (2000). Game Theory and Social Foraging. En: L.A. Dugatkin y H. K. Reeven (Eds). Game Theory and Animal Behavior (pp. 16–37). Oxford University Press: Chicago. [ Links ]
Hewitson, L., Gordon, I. J., & Dumont, B. (2006). Social context affects patch–leaving decisions of sheep in a variable environment. Animal Behaviour, 74, 239–246. [ Links ]
Koops, M.A., & Giraldeau, L.–A. (1996). Producer–scrounger foraging games in starlings: a test of rate–maximizing and risk–sensitive models. Animal Behaviour, 51, 773–783. [ Links ]
Livoreil, B., & Giraldeau, L–A. (1997). Patch departure decisions by spice finches foraging singly or in groups. Animal Behaviour, 54, 967–977. [ Links ]
Parker, G.A. (1984). Evolutionarily stable strategies. En J.R. Krebs & N.B. Davies (Eds). Behaviorual ecology: an evolutionary approach, 2da edición. (pp.30–61). Sunderland: Sinauer. [ Links ]
Vickery, W.L., Giraldeau, L.–A., Templeton, J. J., Kramer, D.L., & Chapman, C.A. (1991). Producers, scroungers and group foraging. Animal Nature, 137, 847–863. [ Links ]
Zamora, A. J., López, M. E., Andrade, L.P., Hernández, D.F., Rosas, M., Zermeño, A., & Cabrera, R. (2007). El estudio del aprendizaje social en grupos de aves. XVIII Congreso Mexicano de Análisis de la Conducta. Xalapa, México. [ Links ]
Nota
Este proyecto de investigación fue posible gracias a que Rosalva Cabrera disfruta de año sabático en la FES Iztacala, UNAM para realizar Estancia de Investigación en el Centro de Estudios e Investigaciones en Comportamiento, CEIC de la Universidad de Guadalajara. Proyecto financiado por apoyo PROMEP/103.5/08/2919 y la beca CONACYT 214051 otorgada al primer autor. Contribución: los tres autores trabajaron en la colección y análisis de datos, obtención de recursos y preparación del manuscrito.

