










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCiencias marinas
versión impresa ISSN 0185-3880
Cienc. mar vol.48 Ensenada ene./dic. 2022 Epub 17-Nov-2023
https://doi.org/10.7773/cm.y2022.3256
Artículos
Distribución y abundancia de la merluza del Pacífico, Merluccius productus, y su relación con el ambiente en el golfo de California, México
1
 http://orcid.org/0000-0002-9044-6449
http://orcid.org/0000-0002-9044-6449
2
http://orcid.org/0000-0002-6638-6191
3
http://orcid.org/0000-0002-5007-9357
1
*
http://orcid.org/0000-0003-1608-8986
1Facultad de Ciencias del Mar, Universidad Autónoma de Sinaloa, 82017 Mazatlán, Sinaloa, Mexico.
2Instituto Nacional de Pesca y Acuacultura, Centro Regional de Investigación Acuícola y Pesquera-Guaymas, 85400 Guaymas, Sonora, Mexico.
3Cátedras CONACYT-Facultad de Ciencias del Mar, Universidad Autónoma de Sinaloa, 82000 Mazatlán, Sinaloa, Mexico.
Las condiciones climáticas y oceánicas determinan el patrón de distribución de la merluza del Pacífico (Merluccius productus). El presente trabajo tuvo como objetivo conocer la relación espaciotemporal entre la distribución y la abundancia de la merluza del Pacífico y el ambiente en el golfo de California. Se analizó la información obtenida de 6 cruceros de investigación realizados entre 2014 y 2019. Los resultados mostraron que la merluza del golfo de California alcanza tallas de 105 cm de longitud total y presenta dimorfismo sexual a tallas >73 cm de longitud total. La mayor biomasa del recurso (68,260.37 t) se encontró en febrero de 2019 entre los 200 y 300 m de profundidad, y la menor, en octubre de 2016 (6,917.80 t) a profundidades de 100-150 m. Los centros de abundancia se localizaron a temperaturas de fondo de entre 13 y 14 °C, al norte de la isla Ángel de la Guarda y entre las grandes islas (isla Tiburón e isla Ángel de la Guarda). Al ajustar modelos aditivos generalizados entre las capturas de merluza de cada estación y distintas variables ambientales, espaciales y temporales, se encontró que la temperatura de fondo fue la variable que mejor describió estos centros de abundancia.
Palabras claves: golfo de California; temperatura de fondo; profundidad; cruceros de investigación; modelos aditivos generalizados
Climate and oceanic conditions determine the Pacific hake (Merluccius productus) distribution pattern. The present work aimed to identify the spatiotemporal relationship between the distribution and abundance of Pacific hake and the environment in the Gulf of California. Information obtained from 6 research cruises carried out between 2014 and 2019 was analyzed. The results showed that the Gulf of California hake reaches a total length of 105 cm, exhibiting sexual dimorphism at total lengths >73 cm. The highest resource biomass (68,260.37 t) was found in February 2019 at depths between 200 and 300 m and the lowest was found in October 2016 (6,917.80 t) at depths between 100 and 150 m. These centers of abundance were located at bottom temperatures between 13 and 14 °C, north of Ángel de la Guarda Island and between the great islands (Tiburón Island and Ángel de la Guarda Island). After adjusting generalized additive models between seasonal hake catches and different environmental, spatial, and temporal variables, bottom temperature was found to be the variable that best described these centers of abundance.
Key words: Gulf of California; bottom temperature; depth; research cruises; generalized additive models
INTRODUCCIÓN
La merluza del Pacífico, Merluccius productus (Ayres, 1855), se distribuye a lo largo de Canadá y Estados Unidos de América, y en México hasta el golfo de Tehuantepec e incluso el golfo de California (Lloris et al. 2005). Es una especie demersal cuyo hábitat se extiende desde la plataforma continental hasta los 1,000 m de profundidad (FAO 1995). A lo largo de su distribución y considerando su estructura poblacional, se tienen identificadas más de 3 unidades de manejo (stocks): la primera, llamada stock costero, se distribuye desde el sur de California hasta el estrecho de la Reina Carlota, Canadá; la segunda, en la parte centro-sur del estrecho de Puget, Estados Unidos de América; y la tercera, en el estrecho de Georgia, Canadá (Iwamoto et al. 2004). En la corriente de California, la merluza presenta movimientos migratorios, hacia el norte para alimentarse y hacia el sur para reproducirse (Bailey et al. 1982). Estos movimientos son asociados a anomalías en la temperatura subsuperficial, la intensidad de la corriente subsuperficial hacia el norte y los cambios en la distribución de sus presas durante los años cálidos y los fríos (Smith et al. 1990, Dorn 1995).
Swartzman (1997) analizó la relación entre los cardúmenes de merluza y la temperatura de fondo en las costas de los Estados Unidos de América, mediante modelos aditivos generalizados y observó un patrón espacial latitudinal, donde las agregaciones en las latitudes norteñas se localizaron a mayor profundidad y alejadas de la costa. En este mismo sentido, Benson et al. (2002) exploraron la relación entre las condiciones del océano y la biomasa de merluzas maduras en aguas canadienses a partir de regresiones lineales y encontraron una relación inversa de la biomasa con las anomalías de surgencias, las cuales explicaron el 77% de la varianza de dichas biomasas. Agostini et al. (2006, 2008), a través de un modelo de regresión geoestadística, encontraron una relación significativa entre los movimientos migratorios de la merluza y la corriente subsuperficial y la profundidad.
El golfo de California alberga 4 masas de agua cuyo patrón de circulación está relacionado al sistema de vientos locales y a las corrientes de marea, lo cual genera surgencias constantes y alta productividad (Torres-Orozco 1993, Alvarez-Borrego 2007). Es por ello que el golfo de California es la región pesquera más importante de México (Lluch-Cota et al. 2007), y en él se han realizado numerosas investigaciones y exploraciones durante décadas. Derivado de estas, a principios de la década de los setenta, se documentó la presencia de la merluza en el golfo, con un hábitat preferencial al norte de la isla Tiburón, Sonora, y con picos máximos de abundancia en los meses de invierno y primavera (Mathews et al. 1974, Padilla-García 1981, Grande-Vidal 1983, Nevárez-Martínez et al. 2007). La distribución de la merluza en el golfo está asociada a las mayores capturas del recurso (INAPESCA 2020), así como al máximo reproductivo, lo cual indica que la actividad pesquera se realiza sobre agregaciones reproductivas (Denton-Castillo 2018).
De manera general, el conocimiento biológico de la merluza del Pacífico que habita en el golfo de California es escaso considerando que es un recurso con potencial de desarrollo pesquero (DOF 2018) y que las condiciones climáticas y oceánicas afectan su patrón de distribución y abundancia durante su ontogenia (Hammel et al. 2015). Este es el primer trabajo que relaciona de manera espaciotemporal el patrón de distribución y abundancia de M. productus con las variables ambientales en el golfo de California.
MATERIALES Y MÉTODOS
Recolección de datos
El material biológico se recolectó durante cruceros de investigación de pesca exploratoria en las regiones central y norte del golfo de California (Fig. 1), realizados en febrero y abril-mayo de 2014, diciembre de 2015, octubre-noviembre de 2016, abril de 2018 y febrero de 2019. En total se realizaron 142 lances de pesca y 127 lances con CTD; estos últimos se realizaron al término del lance de pesca. Los cruceros se realizaron a bordo del BIP XI del Instituto Nacional de Pesca y Acuacultura (México), equipado con una red de arrastre demersal (33.8 m de relinga superior, luz de malla de 6 pulgadas en el cuerpo y 3.5 pulgadas en el copo, tablas de metal) que era largada por la popa. El plan de muestreo incluyó la prospección a lo largo de la costa desde los 27.5° N hasta los 31.5° N y entre 110° W y 114.5° W (Fig. 1), lo que incluyó la costa de Sonora y la de la península de Baja California, hasta la isóbata de los 500 m. Los arrastres se realizaron a una velocidad promedio de 3 nudos.
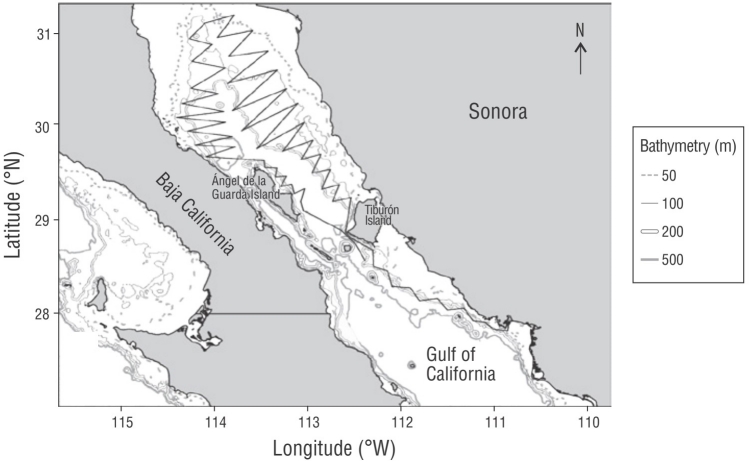
Figura 1 Derrotero de los cruceros (línea sólida gruesa) realizados de 2014 a 2019 para muestrear merluza del Pacífico en el golfo de California.
La captura total de cada lance se colocó sobre la cubierta del barco y se pesó separando a los organismos en grupos taxonómicos (moluscos, crustáceos, peces óseos y peces cartilaginosos). La identificación de los especímenes de merluza del Pacífico se corroboró empleando los caracteres diagnósticos descritos por Robertson y Allen (2002). Cada muestra consistió de 100 ejemplares que fueron medidos (longitud total, LT; ±1 cm) con un ictiómetro y pesados (masa corporal o peso total, en adelante PT para el propósito del estudio; ±1 g) con una báscula marca Torrey. Cada organismo fue eviscerado por la parte ventral para identificar macroscópicamente el sexo.
Estructura de talla y peso
La estructura de tallas de la merluza del Pacífico se analizó por sexo agrupando la LT en intervalos de 2 cm y el PT en intervalos de 200 g. Se evaluó la normalidad con una prueba Shapiro-Wilks, y las diferencias en tallas entre sexos se evaluaron con la prueba Kolmogorov-Smirnov.
Estimaciones de biomasa
La biomasa se calculó empleando 2 metodologías. La primera se realizó a través del método de área barrida (Pierce y Guerra 1994, Hernández-Herrera et al. 1998, Nevárez-Martínez et al. 2000, Rivera-Parra 2001). La estimación de biomasa (kg) se obtuvo con la siguiente fórmula:
con varianza
donde Y
i
es la captura total en el i-ésimo estrato,
A
i
es el área total en el i-ésimo estrato,
a
i
es el área barrida en el i-ésimo estrato,
Con la finalidad de incrementar la precisión en la estimación de la biomasa, como segunda opción se aplicó el método aleatorio estratificado (Schaeffer 1987, Nevárez-Martínez et al. 2000) utilizando un criterio batimétrico. El número de estratos se definió utilizando la regla de Sturges (1926):
mientras que el tamaño del intervalo (TI) se estimó como sigue:
donde n es el número de estaciones, pmax es la profundidad máxima por estación y pmin es la profundidad mínima por estación.
Distribución espacial
Se registró la ubicación geográfica inicial (largada) en cada lance de pesca. Dicha información se utilizó para conocer la distribución espacial y la distribución por talla promedio de la merluza del Pacífico. Las representaciones gráficas se realizaron con el software QGIS 3.6.1.
Selección de variables ambientales
Para analizar el patrón de distribución de la abundancia y las tallas de la merluza del Pacífico en el golfo de California en relación con el ambiente, se consideró el criterio de Swartzman (1997) para la selección de las variables. Como variables espaciales, se consideraron las coordenadas de la posición geográfica inicial de cada lance de pesca (latitud, longitud). Para las variables ambientales, se consideró la temperatura de fondo, la salinidad y la profundidad, obtenidas a través de los lances de CTD al finalizar los lances de pesca. De manera adicional, se utilizó la temporalidad a través de las variables mes y año (Tabla 1).
Tabla 1 Variables explicativas exploradas para predecir la abundancia de la merluza en el golfo de California.
| Variable | Unit | Description |
| Latitude, longitude | Degrees and decimal degrees | Geographic position at the start of a fishing set. |
| Month | Days | Month in which the survey took place; abundance seasonality assessment. |
| Year | Months | Year in which the survey took place. |
| Bottom temperature | Degrees Celsius | Bottom temperature obtained from each CTD set. |
| Salinity | Bottom salinity obtained from each CTD set. | |
| Depth | Meters | Bottom depth obtained from each CTD set. |
Elaboración de los modelos
Para evaluar las características ambientales de las áreas de ocurrencia de merluza del Pacífico en el golfo de California se construyeron modelos aditivos generalizados (MAG). Esta metodología tiene la ventaja de modelar relaciones no lineales entre las variables de respuesta y las variables predictoras mediante el uso de funciones de suavizado de las covariantes (s) (Kainge et al. 2017). La fórmula general de un MAG es la siguiente:
donde g es la función monotónica de enlace, μ i es el valor ajustado, X i es la variable predictora y f j es una función de suavizado con una distribución gaussiana, un enlace (link) tipo identidad (identity) y una base tipo placa delgada (thin plate), la cual ha demostrado buen desempeño cuando se utilizan bases de datos no muy grandes (Zuur et al. 2009). Primero, se utilizó como variable de respuesta la captura (kg) de cada lance de pesca, a la cual se le aplicó el logaritmo natural para estabilizar la razón media/varianza, y como primera aproximación, se utilizaron las variables ambientales (temperatura de fondo y salinidad), las variables espaciales (latitud, longitud y profundidad) y las variables temporales (mes y año) como variables predictoras. Luego, se generó un segundo MAG, donde la variable de respuesta fue la talla promedio en cada lance de pesca y las variables predictoras fueron las mismas variables de la aproximación anterior.
Selección del mejor modelo
La construcción de los MAG se realizó con el enfoque paso a paso (stepwise), es decir, añadiendo una variable a la vez para evaluar el grado de aporte de cada una de las covariables. Para esto, se utilizó la prueba del cociente de verosimilitud (likelihood ratio test) y el criterio de información de Akaike (CIA). Las variables que resultaron en un aporte significativo (P < 0.05) y causaron un decremento del CIA de más de 2 unidades fueron retenidas en el modelo final (Burnham y Anderson 2002). Se exploró de manera visual la distribución espacial y la distribución temporal de los residuales del modelo con una función de autocorrelación. Debido a que al utilizar variables que estén altamente correlacionadas entre sí se puede generar errores en el proceso de modelación, se aplicó una prueba de concurvidad en cada paso de la construcción del modelo. La concurvidad puede interpretarse como una extensión no lineal de la multicolinealidad (Wood 2006). Todo el proceso de modelación se efectuó utilizando las funciones del paquete mgcv (Wood 2006) del ambiente R (R Core-Team 2018).
RESULTADOS
Estructura de tallas y peso
De las 142 estaciones realizadas, 126 resultaron positivas a la presencia de la merluza del Pacífico (88.7%) y 16 fueron negativas (11.3%). En total, se capturaron 2,795 organismos, de los cuales 1,254 fueron machos (44.4%), 1,396 hembras (49.9%) y 145 indeterminados (5.2%). El intervalo de longitudes registrado osciló desde una talla mínima de 12.0 cm LT, para un macho capturado durante el crucero de octubre-noviembre de 2016, hasta una talla máxima de 105.0 cm LT, para una hembra capturada en la prospección de febrero de 2019, con una talla promedio de 42.0 cm LT. En las hembras, la talla fluctuó entre 16.0 y 105.0 cm LT, con una talla promedio de 44.6 cm LT; la longitud mínima prevaleció en febrero de 2014, diciembre de 2015 y octubre-noviembre de 2016, y la talla máxima, en febrero de 2019. En los machos, la longitud varió de 12.0 a 79.0 cm LT, con una talla promedio de 34.8 cm LT; la longitud mínima se encontró en diciembre de 2015, y la máxima, en febrero de 2014. Solo en los cruceros de febrero y diciembre existió diferencia significativa (Kolmogorov-Smirnov: P < 0.05). El peso total varió de 9.8 a 7,500.0 g; el menor peso correspondió a un macho capturado en diciembre de 2015, y el máximo, a una hembra capturada en febrero de 2014. El peso total en las hembras varió de 21.0 a 7,500.0 g, y en los machos, de 9.8 a 4,200.0 g. Los organismos de mayor peso, tanto para las hembras como para los machos, se observaron durante el mes de febrero de 2014 y 2019, respectivamente (Tabla 2). De manera general, se observó que los machos alcanzaron longitudes y pesos menores, a diferencia de las hembras.
Tabla 2 Resumen numérico del peso (masa corporal o peso total; TW, por sus siglas en inglés) y la talla (longitud total; TL, por sus siglas en inglés) de los individuos de merluza capturados durante los cruceros científicos de 2014 a 2019. Min., mínimo; Max., máximo.
| February 2014 | April-May 2014 | December 2015 | ||||||||||
| Males | Females | Males | Females | Males | Females | |||||||
| TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | |
| Min. | 16.1 | 24.0 | 16.0 | 23.0 | 16.4 | 25.0 | 16.5 | 25.0 | 12.5 | 9.8 | 16.0 | 21.1 |
| Mean | 45.0 | 841.9 | 50.9 | 1,379.3 | 39.8 | 501.8 | 49.9 | 789.4 | 22.2 | 81.2 | 42.0 | 996.8 |
| Max. | 85.5 | 4,200.0 | 99.5 | 7,500.0 | 73.0 | 1,920.0 | 93.2 | 5,100.0 | 68.1 | 1,878.5 | 96.0 | 5,446.3 |
| October-November 2016 | April 2018 | February 2019 | ||||||||||
| Males | Females | Males | Females | Males | Females | |||||||
| TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | TL (cm) | TW (g) | |
| Min. | 17.1 | 25.8 | 16.0 | 21.2 | 23.0 | 75.0 | 23.1 | 75.0 | 17.8 | 29.3 | 18.3 | 32.0 |
| Mean | 26.1 | 106.3 | 30.4 | 173.8 | 32.6 | 253.3 | 41.7 | 516.6 | 43.1 | 614.3 | 52.5 | 1,219.0 |
| Max. | 41.4 | 402.1 | 48.4 | 653.4 | 47.0 | 750.0 | 59.1 | 1,400.0 | 75.5 | 2,586.4 | 105.0 | 7,190.4 |
Estimación de biomasa
A partir de las estimaciones de biomasa por el método de área barrida, la menor biomasa se estimó para el crucero de octubre de 2016 (6,887.40 ± 127.30 t), y la mayor, para el de febrero de 2019 (64,823.80 ± 1,287.00 t). El menor coeficiente de variación (CV) se obtuvo en el crucero de abril de 2014 (CV = 2.5), y el mayor, en el crucero de febrero de 2014 (CV = 7.2). A partir del método estratificado, se encontró que las mayores biomasas ocurrieron entre los 200 y 300 m de profundidad, a excepción del crucero octubre-noviembre de 2016, cuando la mayor biomasa se encontró a profundidades de entre 350 y 400 m (Tabla 3). Los cruceros de diciembre de 2015 y abril de 2018 no fueron considerados, ya que el área prospectada fue menor que el 50%.
Tabla 3 Estimaciones de biomasa por crucero y error estándar (SE) para la merluza en el golfo de California. CV, coeficiente de variación.
| Year | Cruise | Estimated biomass (t) | SE | Stratification (t) | SE | CV |
| 2014 | February | 47,642.60 | 1,448.00 | 48,131.11 | 1,080.60 | 7.20 |
| 2014 | April-May | 41,431.99 | 1,393.00 | 40,019.65 | 139.05 | 2.50 |
| 2016 | October | 6,887.40 | 127.30 | 6,917.80 | 50.60 | 3.10 |
| 2019 | February | 64,823.80 | 1,287.00 | 68,260.37 | 547.38 | 3.65 |
Distribución espacial
Las estaciones con menores capturas fueron las cercanas a la costa de Sonora, donde las tallas promedio de merluza del Pacífico predominantes fluctuaron de 25.0 a 50.0 cm LT (Fig. 2). Las estaciones con mayores capturas fueron aquellas cerca de la costa de la península de Baja California, al norte de la isla Ángel de la Guarda y al norte de la isla Tiburón (Fig. 2), área donde se encontraron los organismos de menor (<25.0 cm LT) y mayor (>50.0 cm LT) talla promedio (Fig. 3).

Figura 2 Distribución de las capturas durante los cruceros de investigación realizados en febrero de 2014 (a), abril-mayo de 2014 (b), diciembre de 2015 (c), octubre de 2016 (d), abril de 2018 (e) y febrero de 2019 (f).

Modelación estadística
Las covariables utilizadas en el MAG presentaron una concurvidad menor que 0.3, es decir, poca colinealidad entre las variables utilizadas. La exploración visual de la distribución espacial de los residuales del modelo no mostró un patrón claro (Fig. S1). Con lo que respecta al análisis temporal, la función de autocorrelación mostró que no existió correlación con el tiempo; es decir, no depende un dato de otro (Fig. S2). La temperatura de fondo (intervalo óptimo de 13-14 °C) fue la variable ambiental más importante para la abundancia de este recurso, ya que describió el 20.6% de la variabilidad total, seguida de la variable temporal (mes = febrero), que describió el 15.1%, y la variable espacial (longitud = 13-14° W), que describió el 11.5%. El MAG describió el 38.8% de la variabilidad total (Fig. 4, Tabla 4). En cuanto al efecto que tienen las condiciones ambientales en la distribución de tallas de la merluza del Pacífico en el golfo de California, se encontró que ninguna covariable por sí sola describió significativamente dicha relación, pero se observó que la interacción de la temperatura de fondo con la variable mes explicó el 51.1% de la variación y la interacción de la longitud con la profundidad explicó el 21.5% de la variación (Fig. 5, Tabla 4).
Tabla 4 Modelos aditivos generalizados para la merluza del Pacífico en el golfo de California. AIC, criterio de información de Akaike.
| Abundance model without interaction | Explained deviance (%) | AIC | P |
| +s(Temperature) | 20.60 | 402.20 | <0.01 |
| +s(Salinity) | 2.98 | 416.80 | 0.08 |
| +s(Latitude) | 9.57 | 413.20 | 0.06 |
| +s(Longitude) | 11.50 | 408.20 | <0.01 |
| +s(Depth) | 7.39 | 414.60 | 0.10 |
| +s(Month) | 18.80 | 404.00 | <0.01 |
| +s(Year) | 0.21 | 419.69 | 0.64 |
| Abundance model with interactions | Explained deviance (%) | AIC | P |
| +s(Temperature) | 20.60 | 402.20 | <0.01 |
| +s(Month) | 18.80 | 403.00 | <0.01 |
| +s(Longitude) | 11.50 | 408.20 | <0.01 |
| +te(Longitude, month) | 32.50 | 387.90 | <0.01 |
| Total | 39.30 | ||
| Size model without interactions | Explained deviance (%) | AIC | P |
| +s(Temperature) | 0.11 | 869.21 | 0.73 |
| +s(Salinity) | 4.63 | 864.26 | 0.02 |
| +s(Latitude) | 3.39 | 865.64 | 0.05 |
| +s(Longitude) | 1.66 | 867.54 | 0.18 |
| +s(Depth) | 3.53 | 865.49 | 0.05 |
| +s(Month) | 34.90 | 823.46 | <0.01 |
| +s(Year) | 0.35 | 868.47 | 0.80 |
| Size model with interactions | Explained deviance (%) | AIC | P |
| +s(Longitude, depth) | 21.50 | 863.80 | <0.01 |
| +te(Temperature, month) | 51.10 | 814.70 | <0.01 |
| Total | 61.70 |
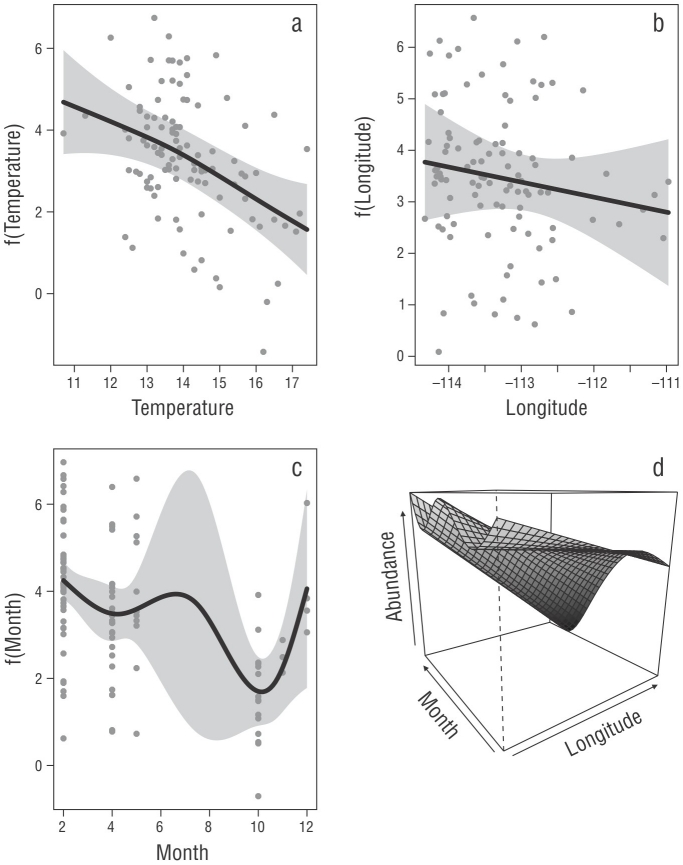
Figura 4 Gráficas de respuesta del modelo aditivo generalizado que muestran el efecto de las funciones significativas para describir las distribuciones espacial, ambiental y temporal de la merluza en el golfo de California. Temperatura (a), longitud (b), mes (c) e interacción mes/longitud (d). El eje y representa el efecto de la covariante con respecto a la abundancia. El área gris representa los intervalos de confianza al 95%.

DISCUSIÓN
Estructura de tallas y peso
En la costa noroccidental mexicana del Pacífico se localiza el golfo de California, un área altamente productiva que sustenta 2 de las pesquerías más importantes en México, la de Camarón y la de Pelágicos menores (Lluch-Cota et al. 2007). Sin embargo, en los últimos años, la merluza del Pacífico ha cobrado importancia por sus volúmenes de captura y sus características biológicas, como las tallas que alcanza. En el presente estudio encontramos organismos con longitudes máximas de 105.0 cm LT, talla máxima cercana a la reportada por Nevárez-Martínez et al. (2007) de 112.0 cm LT para el golfo de California, pero diferente a la reportada por Salinas-Mayoral (2018) de 27.5 cm de longitud patrón (32.5 cm LT) para la costa occidental de la península de Baja California.
Para las costas de Estados Unidos de América, Saunders y McFarlane (1997), Dark (1974) y Best (1962) reportaron tallas promedio máximas de 46.3, 61.8 y 80.0 cm LT, respectivamente. En lo referente a las longitudes máximas, al comparar en cada crucero las tallas por sexo, encontramos que las hembras predominaron a tallas >73.0 cm LT, mientras que los machos alcanzaron tallas menores. Nuestros resultados sugieren que la merluza del Pacífico que habita en el golfo de California presenta dimorfismo sexual. Esta característica fue reportada por Best (1962) para M. productus y para Merluccius hubbsi, con tallas asintóticas para hembras de 90.0 cm y para machos de 60.0 cm (Lorenzo y Defeo 2015). Además, de manera visual, las hembras fueron más robustas (redondas) en comparación con los machos, y aquellas con tallas >73.0 cm LT pesaron más que los machos.
Estimación de biomasas
La biomasa de la merluza del Pacífico del golfo de California ha sido estimada a partir de una serie de cruceros de prospección pesquera desde las décadas de los setenta y los ochenta. Mathews et al. (1974) y Grande-Vidal (1983) reportaron volúmenes de entre 4,000.0 y 28,000.0 t y entre 22,000.0 y 35,600.0 t, respectivamente. Estas estimaciones fueron menores que las obtenidas en este trabajo, las cuales fluctuaron entre 6,887.1 y 64,823.8 t, pero fueron similares a las reportadas por Nevárez-Martínez et al. (2007), a excepción de la estimación para abril de 2006, que fue de 145,000.0 t; este último máximo de biomasa se debió a que Nevárez-Martínez et al. (2007) realizaron 20 lances de pesca más. Por lo anterior, es notable que la población de merluza del Pacífico en el golfo de California ha sido explotada de manera sostenible, y ha sido catalogada como un recurso con potencial de desarrollo (DOF 2018, INAPESCA 2020).
En lo que respecta a los centros de abundancia de la merluza del Pacífico en este estudio, la concentración de organismos fue mayor en estaciones de 3 cruceros (Fig. 2a, b, f) donde la temperatura de fondo fue de 13.2 a 14.1 °C, y la profundidad, de 176-216 m. Este mismo patrón fue reportado por Mathews et al. (1974) y Nevárez-Martínez et al. (2007), quienes encontraron los mayores volúmenes a profundidades similares (184-366 m y 138-275 m, respectivamente). En el crucero de octubre de 2016, los centros de abundancia ocurrieron en estaciones con temperatura de fondo de 12.8 °C y mayor profundidad (350-462 m). Algo similar fue reportado por Nevárez-Martínez et al. (2007), quienes encontraron que la mayor biomasa ocurrió a profundidades de 413 m en noviembre de 2006. En este mismo sentido, Acevedo-Cervantes et al. (2009) reportaron la presencia de la merluza del Pacífico a profundidades de 360 a 450 m en septiembre de 2004.
Durante el crucero de octubre-noviembre de 2016, los centros de abundancia se desplazaron a profundidades mayores que 300 m. Acevedo-Cervantes et al. (2009) mencionaron que en el golfo de California existen 2 comunidades de peces, la primera formada por especies costeras y la segunda por especies típicas de aguas profundas, y esta última se caracteriza por ser poco diversa pero muy abundante. Estos autores consideraron a la merluza del Pacífico como parte de la segunda comunidad, ya que la encontraron a profundidades mayores que 360 m, lo cual coincide con la distribución vertical de las abundancias encontradas en las prospecciones analizadas en el crucero de octubre-noviembre de 2016.
Distribución espacial
La región centro-norte del golfo de California presenta diferentes características topográficas y batimétricas apropiadas para la distribución de la merluza del Pacífico. En la región central se ubica la cuenca Tiburón, entre la isla Ángel de la Guarda y la isla Tiburón, y la cuenca Delfín se ubica al norte de la isla Ángel de la Guarda; ambas cuencas presentan un fondo casi plano y profundidades de 500 y 900 m, respectivamente (Arrellano-Peralta y Medrano-González 2013). Además, la región centro-norte del golfo de California se caracteriza por ser altamente productiva, y el enriquecimiento en la columna de agua se origina por el proceso de mezcla intensa de masas de agua debido a las fuertes corrientes de marea (Lluch-Cota y Arias-Aréchiga 2000).
En este estudio encontramos que la distribución de la merluza del Pacífico es amplia, desde 50 hasta 500 m de profundidad. Sin embargo, la concentración de los organismos fue mayor en áreas con profundidades de entre 200 y 300 m y temperaturas de fondo de entre 13 y 14 °C (Fig. 4a). Estos intervalos de profundidad y temperatura son los óptimos para localizar los centros de abundancia de esta especie en la región centro-norte del golfo de California, lo cual concuerda con la profundidad (110-330 m) a la que opera la flota merlucera en el golfo (EDF 2017). Otra variable a considerar es la temporalidad, ya que los centros de abundancia se registraron a inicios de año (Fig. 4c), lo cual coincide con las fechas de las mayores capturas del recurso (INAPESCA 2020).
Modelación estadística
Los MAG evalúan relaciones no lineales y tienen la capacidad de relacionar los datos mediante funciones de suavizado (Wood 2006). En este sentido, Swartzman (1997) utilizó un MAG para evaluar el efecto del ambiente sobre la biomasa de los cardúmenes de merluza del Pacífico en la región centro-norte de la costa del Pacífico de Estados Unidos de América; en su estudio, encontraron cardúmenes más grandes y con mayor biomasa en el norte que en el sur, y el máximo de biomasa lo encontraron a temperaturas de 7 °C. Nuestro modelo indicó que la temperatura de fondo (intervalo óptimo 13-14 °C), la temporalidad (febrero) y la longitud (113-114 °W ) fueron, en ese orden, las variables más importantes para explicar los centros de abundancia de merluza del Pacífico en el golfo de California. Ressler et al. (2007) mencionaron que la temperatura determina la dinámica poblacional de la merluza del Pacífico, con 2 posibles efectos, la advección de corriente y el nado activo en respuesta a señales ambientales. Agostini et al. (2006) reportaron que la distribución de la abundancia de merluzas del Pacífico adultas en la corriente de California tiene mayor relación con el flujo de la corriente subsuperficial que con un intervalo de temperatura.
Existen trabajos que han relacionado el ambiente con aspectos biológicos del recurso. Por ejemplo, Kainge et al. (2017) relacionaron el ambiente (temperatura, oxígeno, profundidad y posición geográfica) con la estructura de tallas de Merluccius capensis y Merluccius paradoxus en Namibia, África, y encontraron que los organismos de tallas pequeñas habitaron zonas someras, a diferencia de los individuos grandes. En este sentido, el presente estudio mostró que los especímenes grandes de merluza del Pacífico en el golfo de California presentaron afinidad a temperaturas bajas (13.2-14.0 °C).
De manera general, en el golfo de California, M. productus presenta dimorfismo sexual, y las hembras alcanzan tallas (105.0 cm LT) y pesos (7,500.0 g) mayores que los machos. Las mayores abundancias se registraron durante los cruceros de febrero (47,642.0 y 64,823.8 t) y el centro de abundancia se localizó cerca de la costa de la península de Baja California, a profundidades de entre los 200 y 300 m y temperaturas de entre 13.2 y 14.0 °C. Debido a que la variable ambiental más significativa que afectó la distribución de la merluza fue la temperatura de fondo, es importante continuar el monitoreo de este recurso para poder detectar cambios en su distribución relacionados con los cambios en el ambiente. Este trabajo es el primer reporte que aborda la distribución y la abundancia en relación con el hábitat de la merluza del Pacífico en el golfo de California.
AGRADECIMIENTOS
EAT Agradece al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) la beca otorgada para realizar estudios de posgrado. Se agradece al Instituto Nacional de Pesca y Acuacultura (México) el financiamiento al proyecto Merluza, del cual se obtuvo la base de datos utilizada. EME agradece al CONACYT el apoyo otorgado a través del programa “cátedras” (proyecto no. 2137). LASG agradece al proyecto UAS-PTC-131 (DSA 511-6/17-7679) el apoyo de infraestructura y equipo. Los autores agradecen la opinión de 3 revisores anónimos, las cuales ayudaron a mejorar la calidad de nuestro manuscrito.
REFERENCIAS
Acevedo-Cervantes, A., López-Martínez, J., Herrera-Valdivia, E., Rodríguez-Romero, J. 2009. Análisis de la abundancia, dominancia y diversidad de la comunidad de peces demersales de profundidad de 90 a 540 metros en el Golfo de California, México = Abundance, dominance and diversity analysis of the demersal fish community analysis at 90 to 540 meters of depth in the Gulf of California, Mexico. Interciencia. 34(9):660-665. [ Links ]
Agostini, V.N., Francis, R.C., Hollowed, A.B., Pierce, S.D., Wilson, C., Hendrix, A.N. 2006. The relationship between Pacific hake (Merluccius productus) distribution and poleward subsurface in the California Current System. Can J Fish Aquat Sci. 63(12):2648-2659. https://doi.org/10.1139/F06-139 [ Links ]
Agostini, V.N., Hendrix, A.N., Hollowed, A.B., Wilson, C.D., Pierce, S.D., Francis, R.C. 2008. Climate-Ocean variability and Pacific hake: A geostatistical modeling approach. J Mar Sys. 71(3-4):237-248. https://doi.org.10.1016/j.jmarsys.2007.01.010 [ Links ]
Alvarez-Borrego, S. 2007. Oceanografía de la región de las Grandes Islas [Oceanography of the Grandes Islas región]. In: Danemann, G.D., Ezcurra, E. (eds.), Bahía de los Ángeles: Recursos naturales y comunidad. Línea Base 2007 [Bahía de los Ángeles Natural Resources and Community Baseline 2007]. San Diego (CA): Secretariía del Medio Ambiente y Recursos Naturales, Instituto Nacional de Ecología, Pronatura Noroeste, San Diego Natural History Museum. p. 45-65. [ Links ]
Arrellano-Peralta, V.A., Medrano-González, L. 2013. Mamíferos marinos en el golfo de California. Macroecología, impacto humano [Marine mammals in the Gulf of California. Macroecology, human impact]. Mexico: Universidad Nacional Autónoma de México. 256 p. [ Links ]
Bailey, K.M., Francis, R.C., Stevens, P.R. 1982. The life history and the fishery of Pacific Whiting, Merluccius productus. CalCOFI Rep. 23:81-98. [ Links ]
Benson, A.J., Trites, A.W. 2002. Ecological effects of regime shifts in the Bearing Sea and eastern North Pacific Ocean. Fish and Fish. 3(2):95-113. https://doi.org/10.1046/j.1467-2979.2002.00078.x [ Links ]
Best, E.A. 1962. Contribution to the biology of the Pacific hake, Merluccius productus (Ayres). CalCOFI Rep. (9):51-56. [ Links ]
Burnham, K.P., Anderson, D.R. 2002. Model Selection and Multimodel Inference: A practical Information-Theoretic Approach. 2nd ed. Berlin (Germany): Springer-Verlag. 488 p. [ Links ]
Dark, T.A. 1974. Age and growth of Pacific Hake, Merluccius productus. Fish Bull. 73(2):336-355. [ Links ]
Denton-Castillo, J. 2018. Agregaciones y Aspectos reproductivos de la merluza Merluccius productus (Ayres, 1855) en el centro y norte del Golfo de California [Aggregations and reproductive aspects of the Pacific hake Merluccius productus (Ayres, 1855) in the central and northern Gulf of California] [MSc thesis]. [Sinaloa (Mexico)]: Universidad Autónoma de Sinaloa. 108 p. [ Links ]
[DOF] Diario Oficial de la Federación. 2018 Jun 11. Carta Nacional Pesquera 2017. Mexico City: Secretaría de Gobernación. 106 p. [ Links ]
Dorn, M.W. 1995. The effects of age composition and oceanographic conditions on the annual migration of Pacific whiting, Merluccius productus. CalCOFI Rep. 36:97-105. [ Links ]
[FAO] Food and Agriculture Organization of the United Nations. 1995. Pacífico Centro-Oriental. Guía FAO para la Identificación de Especies para los Fines de la Pesca. Rome (Italy): FAO. 1813 p. [ Links ]
Grande-Vidal, J.M. 1983. Evaluación biotecnológica de los recursos demersales vulnerables a redes de arrastre de fondo en el Golfo de California 1978-1980 [Biotechnological assessment of demersal resources vulnerable to bottom trawls in the Gulf of California 1978-1980]. Ciencia Pesquera. 4:97-125. [ Links ]
Hamel, O.S., Ressler, P.H., Thomas, R.E., Waldeck, D.A., Hicks, A.C., Holmes, J.A., Fleischer, GW. 2015. Biology, fisheries, assessment and management of Pacific hake (Merluccius productus). In: Arancibia, H. (ed.), Hakes-Biology and Exploitation. United Kingdom: John Wiley and Sons. p. 234-262. https://doi.org/10.1002/9781118568262.ch9 [ Links ]
Hernández-Herrera, A., Morales-Bojórquez, E., Cisneros-Mata, M.A., Nevárez-Martínez, M.O., Rivera-Parra, G.I. 1998. Management strategy for the giant squid (Dosidicus gigas) fishery in the Gulf of California Mexico. CalCOFI Rep. 39:212-218. [ Links ]
[INAPESCA] Instituto Nacional de Pesca y Acuacultura. 2020. Informe técnico sobre la merluza del Pacífico (Merluccius productus), en aguas marinas del Golfo de California = Technical report on Pacific hake (Merluccius productus) in marine waters of Gulf of California. Guaymas (Sonora): CRIAP-INAPESCA. 23 p. Technical report CRIAP-GMAS/Merluza-008/2020; 22 may 2020. [ Links ]
Iwamoto, E., Ford, M.J., Gustafson, R.G. 2004. Genetic population structure of Pacific hake, Merluccius productus, in the Pacific Northwest. Environ Biol Fishes. 69:187-199. https://doi.org/10.1023/B:EBFI.0000022895.10683.c5 [ Links ]
Kainge, P., Van-der-Plas, A.K., Bartholomae CH, Wieland K. 2017. Effects of environmental variables on survey catch rates and distribution by size of shallow and deep-water Cape hakes, Merluccius capensis and Merluccius paradoxus off Namibia. Fish Oceanogr. 26(6):680-692. https://doir.org/10.1111/fog.12227 [ Links ]
Lloris, D., Matallanas, J., Oliver, P. 2005. Hakes of the World (Family Merlucciidae): an annotated and illustrated catalogue of hake species know to date. Rome (Italy): Food and Agriculture Organization of the United Nations. (FAO Species Catalogue for Fishery Purposes; No. 2). 57 p. [ Links ]
Lluch-Cota, S.E., Aragón-Noriega, E.A., Arreguín-Sánchez, F., Aurioles-Gamboa, D., Bautista-Romero, J.J., Brusca, R.C., Cervantes-Duarte, R., Cortés-Altamirano, R., del-Monte-Luna, P., Esquivel-Herrera, A., et al. 2007. The Gulf of California: Review of ecosystem status and sustainability challenges. Prog Oceanogr. 73(1):1-26. https://doi.org/10.1016/j.pocean.2007.01.013 [ Links ]
Lorenzo, M.I., Defeo, O. 2015. The biology and fishery of hake (Merluccius hubbsi) in the Argentinean-Uruguayan common fishing zone of the southwest Atlantic Ocean. In: Arancibia, H. (ed.), Hakes: Biology and Exploitation. United Kingdom: John Wiley and Sons Ltd. p. 185-210. https://doi.org/10.1002/9781118568262.ch7 [ Links ]
Mathews, C.P., Granados, J.L., Arvizu, J. 1974. Results of the exploratory cruises of the Alejandro de Humboldt in the Gulf of California. México. CalCOFI Rep. 17:101-111. [ Links ]
Nevárez-Martínez, M.O., Balmori-Ramírez, A., Santos-Molina, J.P., Cervantes-Valle, C., López-Martínez, J., Méndez-Tenorio, J.F., Anguiano-Carrazco, M.L., Acevedo-Cervantes, A., Miranda-Mier, E., Morales-Azpeitia, R., et al. 2007. Prospección de merluza y bacalao: distribución, tamaño poblacional e ictiofauna asociada en el Golfo de California = Prospecting for hake and cod: distribution, population size and ichthyofauna in the Gulf of California. Clave: SAGARPA-2003-C01-047. [ Links ]
Nevárez-Martínez, M..O, Hernández-Herrera, A., Morales-Bojórquez, E., Balmori-Ramírez, A., Cisneros-Mata, M.A., Morales-Azpeitia, R. 2000. Biomass and distribution of the jumbo squid (Dosidicus gigas; d’Orbigny, 1835) in the Gulf of California, Mexico. Fish Res. 49(2):129-140. https://doi.org/10.1016/s0165-7836(00)00198-3 [ Links ]
Padilla-García, M.A. 1981. Biomasa de sardina Sardinops sagax y merluza Merluccius productus [Biomass of sardine Sardinops sagax and hake Merluccius productus]. Ciencia Pesquera. 1:35-43. [ Links ]
Pierce, G.J., Guerra, A. 1994. Stock assessment methods used for cephalopod fisheries. Fis Res. 21:255-285. https://doi.org/10.1016/0165-7836(94)90108-2 [ Links ]
R Core Team. 2018. R: A language and environment for statistical computing. R Foundation for Statistical computing. Vienna (Austria). URL. https://www.R-project.org/. [ Links ]
Ressler, H.P., Holmes, J.A., Fleischer, G.W., Thomas, R.E, Cooke, K.C. 2007. Pacific hake, Merluccius productus, Autoecology: A timely review. Mar Fish Rev. 69(1-4):1-24. [ Links ]
Rivera-Parra, G.I. 2001. La pesquería del calamar gigante Dosidicus gigas (d’Orbigny, 1835) en el Golfo de California, México [The fishery of squid giant Dosidicus gigas (d’Orbigny, 1835) in the Gulf of California, Mexico] [dissertation]. [Colima (Mexico)]: Universidad de Colima. 90 p. [ Links ]
Robertson, D.R., Allen, G.R. 2002. Peces costeros del Pacífico oriental Tropical: Un sistema de información. Balboa (Panamá): Instituto Smithsonian de investigaciones Tropicales. [ Links ]
Salinas-Mayoral, C.A. 2018. Dinámica poblacional de la merluza del Pacífico Merluccius productus (Ayres, 1855), en la Costa Occidental de BCS, México [Population dynamics of the Pacific hake Merluccius productus (Ayres, 1855) on the West coast of BCS, Mexico] [MSc thesis]. [La Paz (Baja California)]: Centro de Investigaciones Biológicas del Noroeste. 84 p. [ Links ]
Saunders, M.W., McFarlane, G.A. 1997. Observations on the spawning distribution and biology of offshore pacific hake (Merluccius productus). CalCOFI Rep. 38:147-157 p. [ Links ]
Schaeffer, R.L., Mendenhall, W., Ott, L. 1987. Elementos de muestreo = Elementary Survey Sampling. México City: Editorial Iberoamérica. 321 p. [ Links ]
Smith, B.D., McFarlane, G..A, Saunders, M.W. 1990. Variation in Pacific hake (Merluccius productus) summer length-at-age near southern Vancouver Island and its relationship to fishing and oceanography. Can J Fish Aquatic Sci. 47(11):2195-2211. https://doi.org/10.1139/f90-244 [ Links ]
Sturges, H.A. 1926. The choice of a class interval. JSTOR. 21(153):65-66. https://doi.org/10.1080/01621459.1926.10502161 [ Links ]
Swartzman, G. 1997. Analysis of the summer distribution of fish schools in the Pacific Eastern Boundary Current. ICES J Mar Sci. 54(1):105-116. https://doi.org/10.1006/jmsc.1996.0160 [ Links ]
Torres-Orozco, E. 1993. Análisis volumétrico de las masas de agua en el Golfo de California [Volumetric analysis of water bodies in the Gulf of California] [MSc thesis]. [Ensenada (Baja California)]: Centro de Investigación Científica y de Educación Superior de Ensenada. 80 p. [ Links ]
Wood, S.N. 2006. Generalized Additive Models: An Introduction with R. 1st ed. Florida (USA): Chapman abd Hall/CRC. 410 p. [ Links ]
Zamora-García, O.G., Stavrinaky-Suárez, A., Márquez-Farías, J., Parra-Alaniz, J.C. 2017. Resultados del programa de técnicos a bordo de la pesquería de merluza en el alto golfo de California: primeros tres años de estudio (2015-2017) = Results of the technician program aboard the hake fishery in the upper Gulf of California: first three years of study (2015-2017). Mexico: Environmental Defense Fund de México. 63p. [ Links ]
Zuur, A.F., Ieno, E.N., Walker, N.J., Saveliev, A.A., Smith, G.M. 2009. Mixed Effects Models and Extensions in Ecology with R. New York: Springer. 563 p. http://dx.doi.org/10.1007/978-0-387-87458-6 [ Links ]
Recibido: 01 de Febrero de 2021; Aprobado: 01 de Mayo de 2021
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
