










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkCiencias marinas
versão impressa ISSN 0185-3880
Cienc. mar vol.48 Ensenada Jan./Dez. 2022 Epub 17-Nov-2023
https://doi.org/10.7773/cm.y2022.3239
Artículos
Conducta diurna de Lontra felina en 2 localidades: puerto Matarani y bahía La Ballenita, provincia de Islay, Arequipa, Perú

1Facultad de Ciencias Biológicas, Universidad Nacional de San Agustín de Arequipa, 04002 Arequipa, Peru.
Aunque numerosos estudios han descrito aspectos conductuales de la nutria marina Lontra felina en litorales costeros con presencia de humanos, tanto su conducta como sus patrones de actividad han sido escasamente estudiados en este tipo de hábitats en Perú. Observamos el comportamiento diurno de la nutria marina en 2 localidades con diferente impacto antrópico, puerto Matarani y bahía La Ballenita, entre el 01 de febrero y el 31 de mayo de 2019. Realizamos, en total, 75 eventos de observación, con 271 avistamientos y 12,405.4 s de observación. En el puerto Matarani, la nutria marina fue avistada con mayor frecuencia en el agua, mientras que en La Ballenita, la nutria marina fue mayormente avistada en las rocas (χ 2 = 43.8, g.l. = 1, P < 0.001). La frecuencia de observación de los diferentes tipos de conducta mostró diferencias significativas entre localidades: fue más frecuente observarla transitando y descansando en La Ballenita y transitando y buceando en Matarani (χ 2 = 52.6, g.l. = 5, P < 0.001). Las categorías conductuales no difirieron significativamente en los 4 meses evaluados en puerto Matarani; sin embargo, en La Ballenita registramos variaciones temporales. En ambas localidades, la nutria fue avistada con frecuencia cerca de la costa, con picos de alta actividad entre 1:30 PM y 5:30 PM en puerto Matarani y entre 05:30 AM y 09:30 AM en La Ballenita. Nuestros resultados sugieren que las variaciones conductuales entre estas localidades podrían estar relacionadas con la presencia de residuos pesqueros en puerto Matarani y la presencia de veraneantes en La Ballenita.
Palabras clave: nutria marina; comportamiento; patrones de actividad; litoral peruano; actividades humanas
Although various studies have described behavioral aspects of the marine otter, Lontra felina, on coastal shores with human presence, both its behavior and its activity patterns have been scarcely studied on these types of habitats in Peru. We observed the diurnal behavior of marine otters in 2 locations with different anthropic impacts, Puerto Matarani and La Ballenita Bay, between 01 February and 31 May 2019. We carried out a total of 75 observation events, with 271 sightings and 12,405.4 s of observation. In Puerto Matarani marine otters were sighted more frequently in the water, while in La Ballenita, marine otters were sighted mostly on the rocks (χ 2 = 43.8, d.f. = 1, P < 0.001). The frequency of observations for the different types of behavior showed significant differences between locations: they were more frequently observed moving and resting in La Ballenita and moving and diving in Matarani (χ 2 = 52.6; d.f. = 5; P < 0.001). The behavioral categories did not differ significantly in the 4 months evaluated in Puerto Matarani, however, in La Ballenita we recorded temporal variations. In both locations, otters were frequently sighted near the coast, with peaks of high activity between 1:30 PM and 5:30 PM in Puerto Matarani and between 05:30 AM and 09:30 AM in La Ballenita. Our results suggest that the behavioral variations between locations could be related to the presence of fishing residues in Puerto Matarani and the presence of vacationers in La Ballenita.
Key words: marine otter; behavior; activity patterns; Peruvian coastline; human activities
INTRODUCCIÓN
La nutria marina Lontra felina se distribuye a lo largo del océano Pacífico Sur, desde Perú (9°04ʹ S) hasta Chile, en cabo de Hornos (56° S), y el extremo sur de Argentina, en isla de los Estados (54° S) (Castilla y Bahamondes 1979, Ostfeld et al. 1989, Sielfeld 1992, Parera 1996, Alfaro-Shigueto et al. 2011). Esta especie ocurre tanto en ambientes hidrográficos de los Andes peruanos (BMOD 1968, Hvidberg-Hansen 1970, AEDES 1998, Zeballos et al. 2002, Medina et al. 2018) como en ambientes marinos (Estes 1986). En los litorales costeros, la nutria marina utiliza una franja de entre 100-150 m mar adentro y 30 m en tierra (Castilla y Bahamondes 1979; Medina-Vogel et al. 2007, 2008).
Numerosos estudios han descrito aspectos conductuales de la nutria marina en litorales costeros con presencia de humanos (Medina-Vogel et al. 2007, 2008; Badilla y George-Nascimento 2009; Cursach et al. 2012). En estos ambientes, la conducta oportunista de la nutria marina le permite coexistir con humanos no hostiles, pero muestra un patrón opuesto en áreas de alta hostilidad (Badilla y George-Nascimento 2009, Cursach et al. 2012). A pesar de su persistencia en ambientes antrópicos, la conducta y los patrones de actividad de L. felina han sido escasamente estudiados en litorales con presencia humana en Perú.
La nutria marina tiene una amplia distribución en el litoral peruano (Apaza y Romero 2012), donde su presencia y comportamiento son dependientes de la intensidad de las actividades humanas (Medina-Vogel et al. 2007, 2008; Badilla y George-Nascimento 2009). En la actualidad, la información sobre cómo influyen las perturbaciones antrópicas, como las actividades portuarias y la visitación de playas, en el comportamiento de este depredador aún es limitada. Este problema dificulta la toma de decisiones efectivas para la conservación de este carnívoro, el cual está catalogado como en peligro de extinción en la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza. El objetivo de esta investigación fue comparar la conducta diurna de la nutria marina entre 2 localidades con diferente impacto antrópico de la provincia de Islay en Arequipa, Perú.
Las actividades portuarias influyen en la conducta de la nutria en la provincia de Islay. Se espera mayor actividad conductual de nutrias en localidades con menor influencia de impacto antrópico, es decir, en ambientes naturales donde el impacto es ausente o menor, en contraste con localidades donde las actividades portuarias industriales y de pesca son mayores.
MATERIALES Y MÉTODOS
Área de estudio
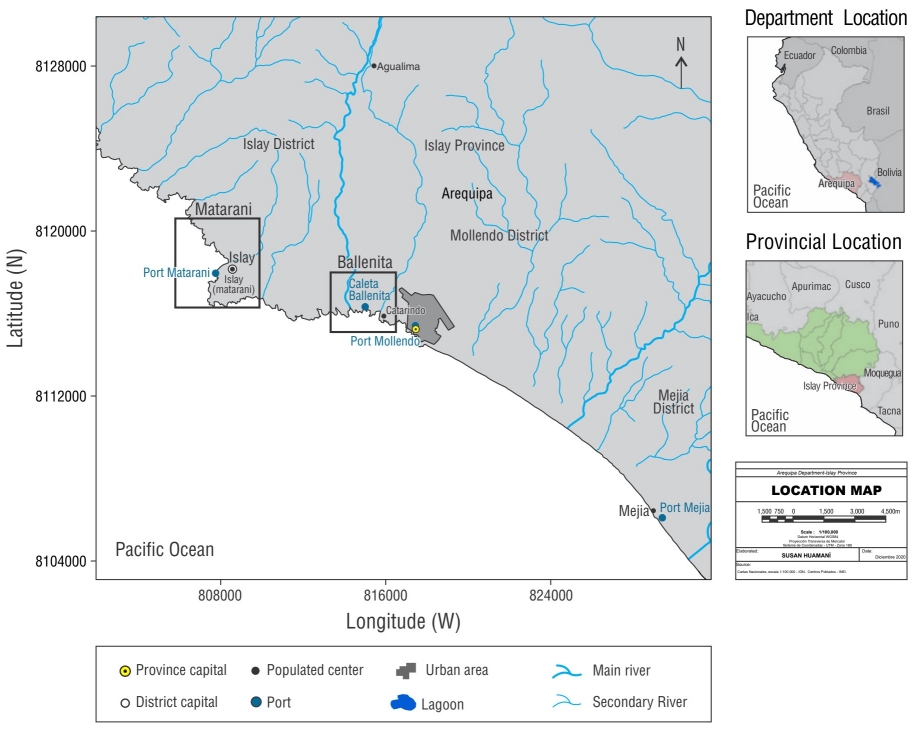
El estudio se realizó en 2 localidades, puerto Matarani (16°59ʹ59.39ʺ S, 72°6ʹ23.06ʺ W) y bahía La Ballenita (17°0ʹ56.32ʺ S, 72°2ʹ28.56ʺ W), ubicadas políticamente en la provincia de Islay, en el departamento de Arequipa, al sur del litoral peruano. El puerto Matarani comprende el muelle Pesquero y el muelle Terminal Internacional del Sur en el distrito de Islay. La Ballenita se conforma de una playa con visitación de veraneantes, rodeada de acantilados en sus extremos, y un islote (Fig. 1).
Recolección de datos
Con la finalidad de registrar el mayor número de actividades conductuales de L. felina, se tomaron los datos según el método de observación ad libitum (Altmann 1974). Observamos el comportamiento de las nutrias marinas entre el 01 de febrero y el 31 de mayo de 2019, durante los periodos estacionales de verano y otoño austral. Para evitar alteraciones en el comportamiento de las nutrias marinas, realizamos las observaciones desde lugares estratégicos (acantilados, peñascos y muelles) procurando un amplio ángulo de observación para diferenciar 2 tipos de superficie, acuática y rocosa, como indicadores de hábitat (Villegas 2000). Las observaciones se realizaron entre 05:30 AM y 5:30 PM, a lo que se le denominó una sesión de observación. A su vez, cada sesión de observación se subdividió en intervalos de una hora denominados eventos de observación, con un máximo de 13 eventos de observación para cada sesión. Contabilizamos los avistamientos de nutria marina durante cada sesión de observación según las recomendaciones de Badilla y George-Nascimento (2009). Los avistamientos se realizaron en 3 horarios: de 05:30 AM a 09:30 AM, de 09:30 AM a 1:30 PM y de 1:30 PM a 5:30 PM. Se cronometró y registró la ocurrencia de 6 pautas conductuales: alimentación, tránsito, buceo, socialización, acicalamiento y descanso. La alimentación es el consumo de presas, y se realiza en agua y en rocas; la nutria marina sostiene con sus extremidades anteriores la presa sobre su pecho mientras se mantiene flotando en el agua (Pizarro 2014). El tránsito es la conducta de mayor frecuencia en las nutrias en el sur del Perú, y se considera como el desplazamiento de un lugar a otro; la contaminación por relaves mineros, la intensa pesca artesanal y la alta concentración de asentamientos humanos hacen que los individuos de L. felina se desplacen en búsqueda de recursos (Pizarro 2014). El acicalamiento se presenta una vez al día, generalmente en las rocas, entre algas o en su madriguera; el acicalamiento es la acción de rascado contra las rocas y el rascado del cuello con las extremidades posteriores, a modo de arreglo asistido, con mordidas o lamidas (Pizarro 2014). El descanso es el periodo de reposo donde las nutrias observan a su alrededor durante 5-18 s (Castilla y Bahamondes 1979).
Elaboramos mapas de distribución de los avistamientos en el puerto Matarani y en La Ballenita con la finalidad de posicionar espacialmente los avistamientos de la nutria marina. Adicionalmente, clasificamos la presencia/ausencia (registros 1/0) de actividades antrópicas (pesca y actividades portuarias) delimitando una línea paralela a la costa, establecida a 50 m de distancia y perpendicular al sitio de observación. Utilizamos esta distancia como referencia debido a que la nutria marina utiliza una franja de entre 100-150 m mar adentro y 30 m en tierra (Castilla y Bahamondes 1979; Medina-Vogel et al. 2007, 2008). Finalmente, dividimos los avistamientos como lejos y cerca de la costa (50 m) empleando la distancia perpendicular a la costa establecida previamente (Badilla y George-Nascimento 2009).
Análisis de datos
Utilizamos la prueba de χ 2 para determinar si existen diferencias en las frecuencias de avistamientos según el tipo de superficie (roca y agua), presencia/ausencia de actividades humanas, el tipo de conducta entre las localidades de estudio y la actividad diurna según horario (Greenwood y Nikulin 1996). Comparamos la conducta de buceo entre ambas localidades mediante la prueba de Wilcoxon-Mann-Whitney para 2 muestras (Zar 1996). Finalmente, evaluamos si existen diferencias en presencia o ausencia de actividades antrópicas utilizando la prueba de probabilidad exacta de Fisher.
RESULTADOS
Conducta diurna
Realizamos, en total, 75 eventos de observación, con 271 avistamientos y 12,405.4 s de actividad. En el puerto Matarani realizamos 52 eventos de observación, con 119 avistamientos y 6,721.8 s de actividad; en La Ballenita, 23 eventos de observación con 152 avistamientos y 5,683.6 s de actividad.
La pauta más frecuente en el puerto Matarani fue el tránsito, con un 61.3% de los avistamientos y el 39.8% del tiempo cronometrado. Esta conducta se realizó con mayor frecuencia en el agua que en las rocas. De 73 avistamientos, el 72.6% de los registros correspondieron al tránsito en agua. En cuanto al buceo, este representó el 28.6% de los registros y fue la segunda actividad más realizada. De los 34 avistamientos que fueron cronometrados en esta localidad, 3 de ellos se registraron mostrando a los animales con presa, y la duración promedio de buceo fue de 68.2 s. Por su parte, las conductas menos frecuentes fueron la alimentación (el 1.7% de los avistamientos y el 7.6% del tiempo cronometrado) y el acicalamiento (el 1.7% de los avistamientos y el 0.9% del tiempo cronometrado; Tablas 1, 2).
Tabla 1 Número de avistamientos de las categorías conductuales de la nutria marina en puerto Matarani y La Ballenita (Perú) en 2019.
| Behavioral category |
Puerto Matarani | La Ballenita | Total | |||||
| Water | Rock | Subtotal | Water | Rock | Subtotal | |||
| Feeding | 1 | 1 | 2 | 1 | 2 | 3 | 5 | |
| Moving | 53 | 20 | 73 | 37 | 33 | 70 | 143 | |
| Diving | 34 | - | 34 | 13 | - | 13 | 47 | |
| Socializing | 4 | 0 | 4 | 4 | 0 | 4 | 8 | |
| Grooming | 0 | 2 | 2 | 0 | 29 | 29 | 31 | |
| Resting | 0 | 4 | 4 | 0 | 33 | 33 | 37 | |
| Total | 92 | 27 | 119 | 55 | 97 | 152 | 271 | |
Tabla 2 Duración (en segundos) de categorías conductuales de la nutria marina en puerto Matarani y La Ballenita (Perú) en 2019.
| Behavioral category | Puerto Matarani | La Ballenita | Total | |||||
| Water | Rock | Subtotal | Water | Rock | Subtotal | |||
| Feeding | 49.0 | 465.0 | 514.0 | 18.0 | 282.0 | 300.0 | 814.0 | |
| Moving | 2,371.6 | 305.8 | 2,677.4 | 1,960.2 | 351.6 | 2,311.8 | 4,989.2 | |
| Diving | 2,317.4 | - | 2,317.4 | 1,438.8 | - | 1,438.8 | 3,756.2 | |
| Socializing | 811.0 | 0.0 | 811.0 | 334.0 | 0.0 | 334.0 | 1,145.0 | |
| Grooming | 0.0 | 60.0 | 60.0 | 0.0 | 643.0 | 643.0 | 703.0 | |
| Resting | 0.0 | 342.0 | 342.0 | 0.0 | 656.0 | 656.0 | 998.0 | |
| Total | 5,549.0 | 1,172.8 | 6,721.8 | 3,751.0 | 1,932.6 | 5,683.6 | 12,405.4 | |
En La Ballenita, la conducta más frecuente fue el tránsito, con el 46.1% de los avistamientos y el 40.7% del tiempo cronometrado. En esta localidad, el tránsito de la nutria marina fue considerablemente similar tanto en agua como en rocas (52.9% y 47.1%, respectivamente), seguido por el descanso (21.7%) y el acicalamiento (19.1%). Por su parte, el buceo representó el 8.6% de las categorías conductuales registradas. De 13 avistamientos, solo en una oportunidad observamos la manipulación de una presa. La duración promedio del buceo en esta localidad fue de 110.7 s. La conducta menos frecuente fue la alimentación (el 2.0% de los avistamientos y el 5.3% del tiempo cronometrado; Tablas 1, 2).
En el puerto Matarani, la nutria marina fue avistada con mayor frecuencia en el agua (el 77.3% de los avistamientos y el 82.5% del tiempo de cronometrado), mientras que en La Ballenita fue mayormente avistada en las rocas (el 63.8% de los avistamientos y el 34.0% del tiempo cronometrado; χ 2 = 43.80, g.l. = 1, P < 0.001). La frecuencia de observación de las diferentes pautas de conducta mostró diferencias significativas entre localidades (χ 2 = 52.60, g.l. = 5, P < 0.001). Fue más frecuente observar a la nutria marina transitando y descansando en La Ballenita y transitando y buceando en Matarani. Las pautas conductuales de alimentación y socialización no presentaron diferencias significativas entre localidades. En Matarani, el tránsito fue más frecuente en agua que en roca (χ 2 = 5.16, g.l. = 1, P < 0.005), mientras que en La Ballenita, no hubo diferencias significativas. La duración de los buceos fue significativamente mayor en Matarani (U = 295.5, P < 0.0001). Respecto al acicalamiento y al descanso, estos solo fueron registrados en ambientes rocosos de ambas localidades.
En el puerto de Matarani las distintas categorías conductuales no difirieron significativamente en la frecuencia de avistamientos en los 4 meses evaluados (febrero-marzo: χ 2 = 3.65, g.l. = 5, P = 0.601; marzo-abril: χ 2 = 2.90, g.l. = 5, P = 0.715; abril-mayo: χ 2 = 6.15, g.l. = 5, P < 0.001). De febrero a marzo, la frecuencia de avistamientos de tránsito incrementó en 44.2% y el buceo en 40.0%; entre marzo y abril, el tránsito disminuyó en 51.2% y el buceo en 57.9%; y entre abril y mayo, el tránsito aumentó, nuevamente, en 33.4% y el buceo en 28.6% (Fig. 2). En La Ballenita, las categorías conductuales difirieron en su frecuencia de avistamientos. Entre febrero-marzo y marzo-abril, no se realizaron comparaciones. Por su parte, entre abril y mayo encontramos diferencias significativas (χ 2 = 9.284, g.l. = 5, P < 0.001). Entre febrero y marzo hubo una disminución en la frecuencia de las categorías conductuales: tránsito, buceo y socialización, las cuales llegaron a cero registros. De marzo a abril hubo un ligero incremento en el tránsito y el descanso, y entre abril y mayo el tránsito incrementó en 53.1%, el buceo en 81.8%, el acicalamiento en 93.1% y el descanso en 81.8% (Fig. 3).

Figura 2 Frecuencias de avistamientos conductuales de nutria marina durante febrero, marzo, abril y mayo de 2019 en puerto Matarani (Perú). ALI: alimentación; TRA: tránsito; BUC: buceo; SOC: socialización; ACI: acicalamiento; y DES: descanso.

Figura 3 Frecuencias de avistamientos conductuales de nutria marina durante febrero, marzo, abril y mayo de 2019 en La Ballenita (Perú). ALI: alimentación, TRA: tránsito, BUC: buceo, SOC: socialización, ACI: acicalamiento y DES: descanso.
La distribución de avistamientos según la distancia (cercanía/lejanía) a la costa no mostró diferencias significativas (χ 2 = 0.56, g.l. = 1, P < 0.4439), aunque fue frecuente observar a la nutria marina cerca de la costa en ambas localidades (Tabla 3). En Matarani hubo mayor registro de avistamientos cerca de la costa en presencia de actividades humanas (96.6%) que en ausencia de estas; sin embargo, no hubo diferencias significativas (prueba de la probabilidad exacta de Fisher, P = 1). En La Ballenita, los avistamientos fueron mayores en ausencia de actividades humanas y cerca de la costa (98.7%) que en presencia de estas (prueba de la probabilidad exacta de Fisher, P = 1).
Tabla 3 Registro de avistamientos de nutria marina de acuerdo con la cercanía/lejanía de la costa y presencia (P)/Ausencia (A) de actividades humanas en las localidades de Matarani y La Ballenita (Perú) en 2019.
| Location | Puerto Matarani | La Ballenita | Total | ||
| P | A | P | A | ||
| Near the coast | 106 | 4 | 2 | 143 | 255 |
| Far from the coast | 9 | 0 | 0 | 7 | 16 |
| Total | 115 | 4 | 2 | 150 | 271 |
Actividad diurna según horario
Encontramos diferencias significativas en las frecuencias de avistamientos entre localidades según los horarios (χ 2 = 4.7, g.l. = 2, P < 0.001). En puerto Matarani el mayor registro de actividad conductual se registró entre 1:30 PM y 5:30 PM, con el 40.3% del número de avistamientos y el 73.2% del tiempo cronometrado. En La Ballenita, el máximo valor de actividad conductual se presentó entre 05:30 AM y 09:30 AM, con el 36.8% de registro de avistamientos y el 22% del tiempo cronometrado (Fig. 4).

DISCUSIÓN
La conducta diurna de la nutria marina descrita en este estudio indica que esta especie persiste en ambientes con diferente impacto antrópico, como las actividades portuarias y la presencia de veraneantes. En ambas localidades, las categorías conductuales observadas fueron alimentación, tránsito, buceo, socialización, acicalamiento y descanso. La frecuencia de ocurrencia de las categorías conductuales de tránsito y buceo sugieren que, en el área de estudio, la nutria marina podría tener un comportamiento generalista y oportunista, el cual ha sido descrito en otras localidades de su distribución geográfica (Badilla y George-Nascimento 2009, Cursach et al. 2012, Poblete et al. 2019).
Es posible que la mayor cantidad de avistamientos de nutria marina realizando tránsito acuático y buceo en puerto Matarani esté relacionada con la menor exposición a las olas de esta localidad, debido a la presencia de un rompeolas y muelles, lo que permite también mayores frecuencias de socialización y menores tiempos de descanso. Por el contrario, en La Ballenita, la mayor exposición a las olas podría explicar las frecuencias similares entre el tránsito terrestre y el tránsito acuático y los consecuentes mayores tiempos de descanso y acicalamiento. Este fenómeno fue descrito en el norte de Chile (Villegas et al. 2007), donde se observó un patrón opuesto en el litoral central (Badilla y George-Nascimento 2009). No obstante, es importante considerar que otros factores ambientales, como la disponibilidad de recursos, la presencia de actividades humanas o las diferencias en la productividad, también están relacionados con este comportamiento (Villegas et al. 2007, Badilla y George-Nascimento 2009). Sugerimos que nuestra hipótesis respecto a la influencia de la exposición a las olas sea puesta a prueba en futuros estudios que permitan determinar, bajo un enfoque cuantitativo, cómo se relacionan las diferencias del oleaje con el tránsito de este mustélido marino.
Con relación a las variaciones conductuales durante el tiempo de muestreo, supusimos la posibilidad de que la presencia de patrones conductuales similares durante los meses de estudio en el puerto Matarani podría estar relacionada con el desarrollo de actividades pesqueras en esta localidad. De esta forma, los residuos pesqueros podrían representar un recurso constante para la nutria marina, lo cual altera su comportamiento. Por otra parte, los diferentes patrones de comportamiento registrados en La Ballenita pueden explicarse por las fluctuaciones de la presencia humana en esta localidad. Por ejemplo, entre los meses de febrero y marzo, la presencia constante de veraneantes y sus impactos asociados, como el ruido y los desechos sólidos y orgánicos, y la presencia de perros vagabundos alteran la calidad de hábitat para la nutria marina, lo cual explica la ausencia de registros de esta especie en ese periodo de tiempo. Sugerimos que esta inferencia se interprete con cautela, debido a que es posible que otros factores no considerados en este estudio puedan explicar las diferencias entre ambas localidades. Generalmente, las variaciones en el tiempo dedicado a diversas conductas pueden estar mediadas por la presencia de competidores, la temporada de cría o las fluctuaciones ambientales en diferentes épocas del año (Badilla y George-Nascimento 2009).
Otro aspecto importante a considerar es la mayor presencia de la nutria marina cerca de la costa en ambas localidades. Sin embargo, en puerto Matarani, la presencia de la nutria se realizó en presencia de humanos, mientras que en La Ballenita, el uso de la costa se realizó en ausencia de humanos. La capacidad que exhibe la nutria para aprovechar residuos de pesca (Cursach et al. 2012) podría explicar su mayor ocurrencia en el puerto Matarani, en tanto que la presencia de veraneantes no ofrecería desechos aprovechables por las nutrias; además, las actividades invasivas de los veraneantes y sus mascotas pueden promover una percepción de mayor amenaza en la nutria marina (Badilla y George Nascimento 2009). Numerosos estudios sugieren que, aunque la nutria marina puede persistir en ambientes con presencia humana, sus respuestas ante diferentes actividades humanas dependen de la hostilidad de estas (Medina-Vogel et al. 2007, 2008; Badilla y George-Nascimento 2009; Cursach et al. 2012).
Respecto a los patrones de actividad diurna, nuestros resultados divergen entre ambas localidades de estudio. Según los avistamientos, la mayor actividad de las nutrias a partir del mediodía en puerto Matarani (localidad con presencia de residuos de pesca) es comparable con los resultados presentados por Badilla y George-Nascimento (2009). Por su parte, el pico de actividad matutino de la nutria marina en La Ballenita puede explicarse por la menor actividad humana durante la mañana. Un factor que refuerza esta hipótesis está relacionado con la ausencia de veraneantes en La Ballenita durante mayo, tiempo en el cual la nutria marina retorna a su hábitat en La Ballenita e incrementa sus actividades.
Los resultados que presentamos en este estudio representan la primera aproximación al entendimiento de la conducta diurna de la nutria marina en la provincia de Islay, en donde las actividades humanas ejercerían diferentes impactos a la biodiversidad asociada a este litoral marino. Medina-Vogel et al. (2008) sugieren que el desarrollo humano en litorales costeros puede incrementar la perturbación al hábitat de este depredador, debido a la contaminación del aire y el agua, la presencia de ruido, la presencia de perros vagabundos, la iluminación artificial y la competencia por recursos. Bajo este panorama, es fundamental determinar cómo responde este depredador a perturbaciones cada vez más prevalentes. De esta manera, será posible brindar recomendaciones plausibles a los tomadores de decisiones para conservar efectivamente a la nutria marina. Los datos provenientes de esta investigación pueden ser utilizados como base para futuros esfuerzos de conservación e investigación en el área de estudio y otras localidades en diferentes latitudes.
AGRADECIMIENTOS
Agradecemos a la Universidad Nacional de San Agustín (UNSA) el financiamiento del proyecto de investigación según el Contrato de Subvención N°TP-16-2018 UNSA. Al Servicio Nacional Forestal y de Fauna Silvestre agradecemos la autorización de la investigación (Resolución de Dirección General N°385-2019-MINAGRI-SERFOR-DGGSPFFS e Informe Técnico N°0688-2019-MINAGRI-SERFOR/DGGSPFFS-DGSPFS). La ONG Procarnivoros apoyó desinteresadamente en campo y gabinete. Al Instituto Científico Michael Owen Dillon IMOD agradecemos el apoyo en el respaldo académico y científico en las solicitudes de ingreso a las áreas de estudio. Los biólogos Carlos Ruelas, Jhon Muñuico, Harold Calderon, Stephanie Salazar y Mary Bedoya participaron en la toma de datos en campo. Richard Felix apoyó con la elaboración de mapas.
REFERENCIAS
AEDES. 1998. Estudio de la biodiversidad vegetal y animal, cuenca del Cotahuasi: riqueza faunística Zona II. Serie Agenda Local 21 de La Unión. Arequipa (Peru): AEDES. 425 p. [ Links ]
Alfaro-Shigueto, J., Valqui, J., Mangel, J.C. 2011. Nuevo registro de la nutria marina Lontra felina (Molina, 1782) al norte de su distribución actual = New record of the marine otter Lontra felina (Molina, 1782) north to its current distribution. Ecología Aplicada. 10(2):87-91. [ Links ]
Altmann, J. 1974. Observational Study of Behavior. Sampling Methods. Behavior. 49(3-4):227-267. https://doi.org/10.1163/156853974x00534 [ Links ]
Apaza, M., Romero, L. 2012. Distribución y observaciones sobre la población de la nutria marina Lontra felina (Molina 1782) en el Perú = Distribution and observations on the population of marine otters Lontra felina (Molina 1782) in Peru. Rev Peru biol. 19(3):285-298. https://doi.org/10.15381/rpb.v19i3.1064 [ Links ]
Badilla, M., George-Nascimento, M. 2009. Conducta diurna del chungungo Lontra felina (Molina, 1782) en dos localidades de la costa de Talcahuano, Chile: ¿Efectos de la exposición al oleaje y de las actividades humanas? = Daylight behavior of the sea otter Lontra felina (Molina, 1782) in two localities off the coast of Talcahuano, Chile: effects of wave exposure and human activities? Rev Biol Mar Oceanogr. 44(2):409-415. http://doi.org/10.4067/S0718-19572009000200014 [ Links ]
[BMOD] British Ministry of Overseas Development. 1968. Appendix III to Recommendations on the Conservation of Wildlife and the Establishment of National Parks and Reserves in Peru (unpublished report). England: BMOD. 100 p.+11 maps. [ Links ]
Castilla, J.C., Bahamondes, I. 1979. Behavioural and ecological observations in Lutra felina (Molina 1782) (Carnivora: Mustelidae) in Central and North-Central zones of Chile = Observaciones conductuales y ecológicas sobre Lutra felina (Molina) 1782 (Carnivora: Mustelidae) en las zonas Central y Centro-Norte de Chile. Arch Biol Med Exp. 12:119-132. [ Links ]
Cursach, J.A., Rau, J.R., Ther, F., Vilugrón, J., Tobar, C.N. 2012. Sinantropía y conservación marina: el caso del chungungo Lontra felina en el sur de Chile = Synanthropy and marine conservation: the case of the marine otter Lontra felina in Southern Chile. Rev biol mar oceanogr. 47(3):593-597. http://doi.org/10.4067/S0718-19572012000300022 [ Links ]
Estes, J.A. 1986. Marine otters and their environment. Ambio. 15(3):181-183. https://www.jstor.org/stable/4313245 [ Links ]
Greenwood, P.E., Nikulin, M.S. 1996. A guide to chi-squared testing. USA: Wiley-Interscience. 304 p. [ Links ]
Hvidberg-Hansen, H. 1970. A survey of the coast otter Lutra felina (Molina, 1782) in the peruvian rivers of Camaná and Ocoña. [place unknown]: Food and Agriculture Organization of the United Nations. 116 p. [ Links ]
Medina, C., Díaz, D., Málaga, B., Medina, Y., López, E. 2018. Dieta de Lontra felina (Carnívora, Mustelidae) en la parte alta del río Ocoña, Arequipa - Perú. IV Congreso Latinoamericano, VIII Congreso Boliviano de Mastozoología; 10-13 Jul 2018. La Paz (Bolivia): Colección Boliviana de Fauna (Museo Nacional de Historia Natural-Instituto de Ecología Universidad Mayor de San Andrés). 289 p. [ Links ]
Medina-Vogel, G., Boher, F., Flores, G., Santibáñez, A., Soto-Azat, C. 2007. Spacing behavior of marine otters (Lontra felina) in relation to land refuges and fishery waste in central Chile. J Mammal. 88(2):487-494. https://doi.org/10.1644/06-MAMM-A081R1.1 [ Links ]
Medina-Vogel, G., Merino, L.O., Monsalve-Alarcón, R., Vianna, J.-de-A. 2008. Coastal-marine discontinuities, critical patch size and isolation: implications for marine otter conservation. Animal Conservation. 11(1):57-64. https://doi.org/10.1111/j.1469-1795.2007.00151.x [ Links ]
Ostfeld, R.S., Ebensperger, L., Klosterman, L.L., Castilla, J.C. 1989. Foraging, activity budget and social behavior of the South American marine otter Lutra felina (Molina, 1782). Nat Geogr Res. 54:422-238. [ Links ]
Parera, A. 1996. Las nutrias verdaderas de la Argentina. Buenos Aires: Boletín Técnico de la Fundación Vida Silvestre Argentina. 39 p. [ Links ]
Pizarro Neyra, J. 2014. La Nutria Marina en Tacna: Biología, Comportamiento, Ecología y Conservación. Tacna (Peru): Proyecto NMAM. [ Links ]
Poblete, A.A., Górski, K., Moscoso, J. 2019. Estimación de dietas del chungungo Lontra felina (Molina, 1782) en dos localidades de la región del Biobío, Chile = Estimation of the diet of chungungo Lontra felina (Molina, 1782) in two locations of the coast of the Biobío Region, Chile. Gayana (Concepc.). 83(1):1-9. https://dx.doi.org/10.4067/S0717-65382019000100001 [ Links ]
Sielfeld, W. 1992. Abundancias relativas de Lutra felina (Molina, 1782) y L. provocax (Thomas, 1908) en el litoral de Chile. Ciencias del mar. 2:3-12. [ Links ]
Villegas, L.N. 2000. Bioecología de la nutria marina Lutra felina (Molina 1782). “Chungungo”, en la provincia de Islay (Arequipa- Perú), 1982 [BSc thesis]. [Arequipa (Peru)]: Universidad Nacional de San Agustín UNSA. 74 p. [ Links ]
Villegas, M.J., Aron, A., Ebensperger, L.A. 2007. The influence of wave exposure on the foraging activity of marine otter, Lontra felina (Molina, 1782) (Carnivora: Mustelidae) in northern Chile. J Ethol. 25:281-286. https://doi.org/10.1007/s10164-006-0024-x [ Links ]
Zar, J. 1996. Biostatistical analysis. Englewood Cliffs (USA): Prentice Hall. 662 p. [ Links ]
Zeballos, H., López, E., Barrionuevo, R. 2002. Registro y distribución de los mustélidos de los Andes y la costa sur del Perú. Revista de investigación Dilloniana de la UNSA. 2:156-161. [ Links ]
Recibido: 20 de Diciembre de 2020; Aprobado: 05 de Noviembre de 2021
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
