











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The ocean floor provides an excellent sedimentary record that can help us recognize and understand paleoceanographic and paleoclimatic changes (Esparza-Álvarez 1999). The biogenic component in these sediments is important because it is largely represented by phyto- and zooplankton (Aguirre 1989). Moreover, it provides a record of the dominant organic productivity and the sedimentological and climatological conditions in the basin where it settled (Emery and Hülsemann 1961). Much of this record is made up of minerals and consequently tends to be affected by dissolution during diagenesis (Molina 2017). Nevertheless, palynomorphs are highly resistant to acidification and are very useful when reconstructing peleoceanographic characteristics. Laminated sediments, in particular, tend to preserve organic matter with almost no alteration and thus make for high-resolution records. Marine palynomorphs comprise the resistant cysts of different algae (e.g., Chlorophyceae), dinoflagellates, acritarchs, and diapausing copepods.
Dinoflagellates
Dinoflagellates are unicellular microorganisms living in fresh water and seawater that range between 50 and 200 µm in size (Bütschli 1885, Fensome et al. 1993). Presently 1,555 free-living marine dinoflagellate species are recognized around the world’s oceans (Gómez 2005). The life cycle of dinoflagellates has 2 main stages, one consisting of mobile planktonic cells and another of non-motile benthic cells that form cysts. Only 96 living species (6% of the total) produce resistant organic cysts ( Zonneveld and Pospelova 2015). These cysts can persist for long periods of time, so they are known to have existed for longer than 200 million years (Evitt 1985, Fensome et al. 1993). In addition, cysts tend to be abundant along continental margins (Stover et al. 1996), where the areas with the highest primary productivity are found. The concentrations of these cysts in sediments are directly associated with the abundance of copepods in their planktonic phase (Peña-Manjarrez et al. 2009).
Copepods
Copepods are a group of small crustaceans (0.2-28 mm) comprising ~12,000 species (Vives and Shmeleva 2007). Although they live in all types of environments and are found at every latitude, these species are mostly marine copepods. Marine planktonic copepods belonging to the order Calanoida produce diapausing eggs when environmental conditions (temperature, salinity, light, etc.) are not suitable for a dormant state (Fernández-de-Puelles 2015). These eggs are so resistant that they have been described in pre-Holocene marine sediments (Uye et al. 1979). This crustacean order can be used as a proxy for water masses and marine currents (Lavaniegos 2014), and it has even been used as a biological indicator of the effects of climate phenomena such as El Niño (Jiménez-Pérez and Lara-Lara 1988).
Regional climatic factors
Two major climate events occurring in the northeastern Pacific Ocean have been studied, namely El Niño/Southern Oscillation (ENSO) and the Pacific Decadal Oscillation (PDO). The ENSO phenomenon exhibits interannual periodicity, is significantly associated with sea surface temperature (SST) anomalies, and presents phases that range from positive, or warm (El Niño), to negative, or cold (La Niña). During El Niño events surface waters in the eastern Pacific Ocean become warmer and subtropical and tropical water masses shift to mid latitudes. Conversely, La Niña events are characterized by the cooling of waters in the central and eastern equatorial Pacific. Under La Niña conditions, trade winds strengthen and upwelling of cold waters intensifies along the eastern equatorial Pacific (NOAA 2018). The PDO exhibits a periodicity of 20-30 years and presents a positive phase (PDO+), or warm phase, and a negative phase (PDO-), or cold phase. ENSO influences mainly the tropics, whereas the PDO affects the North Pacific and North America (Maturana et al. 2004). The most important climate variations occur when the PDO and ENSO are in phase.
Primary productivity
Variability in primary productivity in the northeastern Pacific Ocean is associated with changes in the physicochemical conditions in the water column, which undergo interannual modulations during El Niño and La Niña events (Juárez et al. 2014). Laminated sediments in this basin indicated an increase in the abundance of heterotrophic dinoflagellate species during the cold phases of the PDO and ENSO in the 20th century (Castañeda-Quezada 2016).
When upwelling intensifies, turbulence in the water column increases and so does nutrient availability in the euphotic zone. As a result, diatoms, the main nutrient source for heterotrophic dinoflagellates, and copepods that feed on these heterotrophs proliferate. Conversely, when turbulence decreases and nutrients become less available, a thermocline and/or nutricline can form and prevent the nutrient-rich cold water from upwelling, thus reducing productivity. These low-turbulence conditions are more favorable for the proliferation of autotrophic dinoflagellates because, compared to diatoms, these dinoflagellates are more capable of moving vertically in the water column.
Objectives
During the 1811-1907 (dates of the Gregorian calendar, or common era, CE) period, the sea surface temperature index (SSTI) was dominated by negative anomalies (Jones and Mann 2004), suggesting that primary productivity in the Magdalena Basin increased in that period. Through this study, we aim to elucidate primary productivity and temperature conditions in the southern continental margin of the Baja California Peninsula (Mexico) during the 19th century. The hypothesis, to this end, is that during the 1811-1907 CE period, abundances of autotrophic and heterotrophic dinoflagellates and of copepods were associated with oceanographic conditions and regional forcings such as ENSO and PDO in the Magdalena Basin, Baja California Sur. The aim of the present study was to determine the association of palynological content in laminated sediments with proxies and reconstructions of regional climate indices available for this period. With this, we intend to expand our knowledge on the evolution of oceanographic and climatic conditions in the northeastern Pacific.
Materials and methods
Study area
The Magdalena Basin (23.42ºN, 111.23ºW; Fig. 1) is influenced to the north by the southern portion of the California Current and to the south by the North Equatorial Current. Bioturbation of the basin seafloor is absent because of the low oxygen levels (Hartnett et al. 1998, Esparza-Álvarez 1999, van Geen et al. 2003), allowing for the preservation of marine palynomorphs in laminated sediments.
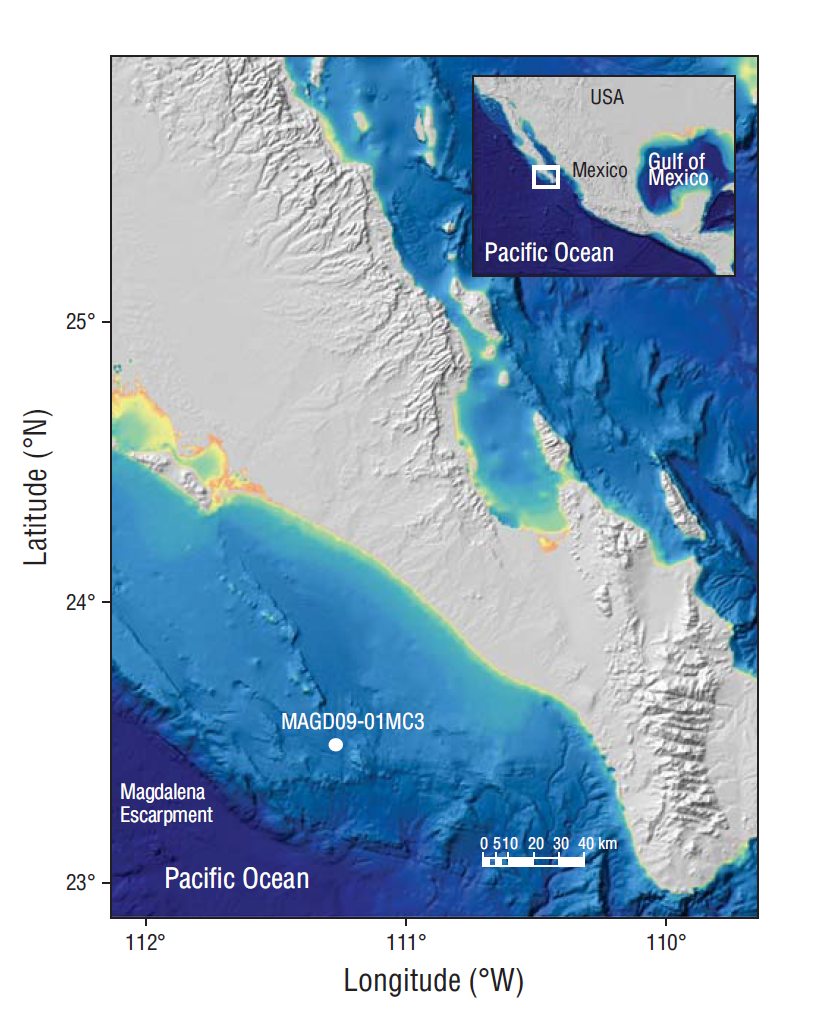
Figure 1 Location of core MAGD09-01MC3 in the study area in the Magdalena Basin, Baja California Sur (Mexico).
For most of the western coasts of the Baja California Peninsula and California, SST and primary productivity show 2 very different seasons. During spring and summer, prevailing upwelling events, with cold, nutrient-rich waters and the presence of species associated with high productivity, promote an increase in regional primary productivity. These conditions intensify during the cold phase of the PDO and/or La Niña events (Mantua et al. 1997, Maturana et al. 2004, Barron et al. 2013). During the winter, northerly winds weaken and warmer water from the southeast is transported by the Southern California Countercurrent and the California Undercurrent (Bringué et al. 2014). These warm conditions become intense during the warm phase of the PDO and/or during El Niño events, due to thermocline and nutricline shoaling in the northeastern Pacific, which causes water stratification and a concurrent decrease in primary productivity on the western coast of Baja California (Espinosa-Carreón et al. 2004, Espinosa-Carreón and Valdez-Holguín 2007, Ortiz-Ahumada 2015).
In general, primary productivity in the region surrounding the Magdalena Basin tends to be relatively low (e.g., chlorophyll values of 0.32 mg/m3 in water samples from 1999; Lara-Lara and Bazán-Guzmán 2005). The variability in marine biological productivity in the Magdalena Basin has been studied using biogenic opal, organic carbon, and CaCO3 (Sánchez and Carriquiry 2007).
Sampling and analysis
A core of laminated sediments from the Magdalena Basin (MAGD09-01MC3, 35 cm long) was extracted at 520 m depth in October 2009 aboard the RV New Horizon, in an expedition led by Dr. William Berelson from the University of Southern California. The lithology for the sediment core was homogenous and consisted of clays and laminated silts (Fig. 2). The core was sampled onboard at 1-mm intervals to obtain a record of the upper 10 cm with 100 samples. The rest of the segment (25 cm) was transported to the Paleoceanography Laboratory in the Ciencias Marinas Faculty at the Autonomous University of Baja California, were it was sampled at 1-cm intervals and X-rayed to check for changes in density. This study presents the results of the palynological analysis of the upper 5-10 cm interval (1811-1907 CE) at a millimetric scale. The 51 earlier samples (1909-2009 CE) were previously analyzed by Castañeda-Quezada (2016).

Radiochronology
For dating of core MAGD09-01MC3, 5 samples were analyzed by the 210Pbexc method at the Academic Service for Dating of the Marine Sciences and Limnology Institute of the Autonomous University of Mexico (Mazatlán, Sinaloa). Though the number of samples was limiting, the analyzed samples exhibited the typical pattern of higher 210Pbexc concentrations at the surface (Table 1), so the constant flux/constant sedimentation model (Sanchez-Cabeza and Ruiz-Fernández 2012) was used to calculate ages corresponding to the Gregorian calendar (CE).
Table 1 210Pbexc concentrations and age calibrations with calendar years (common era, CE) for the analyzed samples taken from core MAGD09-01MC3.
| Depth (cm) | 210Pbexc concentration (Bq/kg) | Calendar year (CE) |
| 0.15 | 1,762 ± 132 | 2007 |
| 0.30 | 1,953 ± 127 | 2005 |
| 2.55 | 521 ± 35 | 1969 |
| 5.25 | 254 ± 19 | 1925 |
| 7.65 | 185 ± 15 | 1887 |
A linear regression between the 210Pbexc age profile and depth was used to calculate sedimentation velocities. We found a very low sedimentation velocity of 0.62 ± 0.01 mm/year with high statistical significance (P = 7.04 × 10-06) (Castañeda-Quezada 2016). Assuming a constant sedimentation rate, the 49 samples analyzed in the present study represent an approximate sedimentary record from 1811 to 1907 CE, with a 2-year resolution per sample.
Palynological processing
All 49 samples were processed using the method for extracting palynomorphs described by Wood et al. (1996), with no oxidation to preserve all dinoflagellate taxa (Rochon et al. 1999) and with modifications made in the Laboratory of Palynology of the Center for Scientific Research and Higher Education at Ensenada (CICESE, for its Spanish acronym). Samples were lyophilized for 24 h at -58 ºC, and, on average, 0.97 g of dry sediment were weighed per sample. Each sample was ground in a mortar and placed in a beaker. A tablet of Lycopodium clavatum spores of batch 483216 (1 tablet = 18,583 spores, s: ±1,708) was then added as an exotic marker to calculate the absolute abundance of palynomorphs.
We added cold hydrochloric acid (37.4%) to remove carbonates and hydrofluoric acid (48%) to eliminate silicates. Heavy metals were separated using sodium polytungstate (2.0 density) to isolate the palynological fractions. For microscopic observations, 49 thin laminae were prepared and analyzed qualitatively and quantitatively. Dinoflagellate cysts were identified using the descriptions for the holotypes of the species and the online databases of Zonneveld and Pospelova (2015) (Marum, https://www.marum.de/Karin-Zonneveld/dinocystkey.htm) and Williams et al. (2017) (Dinoflaj3, http://dinoflaj.smu.ca/dinoflaj3/index.php/Main_Page). These catalogues contain all living dinoflagellate species known to date that produce organic cysts.
Concentrations were calculated using the following equation (Benninghoff 1962):
where C is the concentration of palynomorphs per gram of dried sediment, d c is the number of counted palynomorphs, L t is the number of Lycopodium spores per tablet, t is the number of tablets added to the sample, L c is the number of counted Lycopodium spores, and w is the weight of dried sediments in grams. These concentrations are expressed as palynomorphs (pal), dinoflagellate cysts (cys), or copepod eggs (cop) per gram of sediment (pal/g sed, cys/g sed, and cop/g sed, respectively).
Climate indices
To identify the paleoceanographic and paleoclimatic changes that occurred in the Magdalena Basin during the 19th century, the regional SST, ENSO, and PDO indices for the 1811-1907 CE time frame were used, and their correlation with palynological data was analyzed. Data for SSTI were obtained from Jones and Mann (2004), who used time series with proxies and various global models. Data for the ENSO index were obtained from the North American Drought Atlas tree ring records for the central part of the eastern tropical Pacific (Li et al. 2011). The PDO index (Biondi et al. 2001) was constructed using dendrochronological reconstructions from 5 sites, from the Transverse Ranges in southern California (USA) to Sierra San Pedro Mártir in northern Baja California (Mexico); in addition, 2 other sites were chosen, one close to San Gorgonio Mountain in California and one in Whirlpool Point in Alberta (Canada) (MacDonald and Case 2005). All indices were averaged biennially to equal the sampling resolution and are expressed in degrees Celsius.
Multivariate analysis
Pearson correlation (r) matrices were computed to evaluate relationships between variables. The following statistical hypotheses were formulated to prove statistically significant correlations at a confidence level of 95% (P < 0.05):
H0: palynological variables are not correlated with climate indices (P = 0);
H1: palynological variables are correlated with climate indices (P ≠ 0).
According to the results obtained from the correlations, and to determine if correlation values were high or low and significant, a limit for r higher than ±0.50 was established to indicate high correlations and a value of P < 0.05 was established to indicate statistical significance. A canonical correlation analysis was performed considering the low correlations (r < 0.50) between SSTI and palynological variables (Table 2).
Table 2 Statistical values for palynomorph concentrations per gram of sediment (pal/g sed) in the analyzed samples taken from core MAGD09-01MC3. The values for the 1907-2009 interval were taken from Castañeda-Quezada (2016). Concentrations are indicated in 4 different time intervals. The correlations (r) between concentrations of copepods and heterotrophic and autotrophic dinoflagellates, and the sea surface temperature index (SSTI) is shown with their respective significance values (P). The significant correlation is indicated in bold.
| Concentration (pal/g sed) | Average concentration per time interval | SSTI | ||||||||||
| Variable | Mean | Min | Max | σ | 1811-1907 | 1811-1879 | 1881-1907 | 1907-2009 | r | P | ||
| Copepods | 2,407 | 181 | 12,788 | 2,708 | 2,407 | 1,228 | 5,500 | 868 | 0.46 | 0.001 | ||
| Heterotrophs | 138 | 4 | 1,053 | 152 | 138 | 108 | 211 | 824 | 0.11 | 0.450 | ||
| Autotrophs | 49 | 0 | 363 | 66 | 49 | 56 | 31 | 19 | -0.30 | 0.038 | ||
| Total dinoflagellates | 187 | 23 | 1,053 | 169 | ||||||||
| Total palynomorphs | 2,635 | 204 | 13,200 | 2,733 | ||||||||
Canonical correlation analysis can determine the linear relationship between a group of dependent variables and a group of independent variables, which are grouped into a predicted value to produce the maximum correlation coefficient between the 2 sets of variables (Davis 2002, StatSoft 2013). This test was done to determine the canonical correlation between the palynological data (dependent variables) and the SSTI and ENSO (independent variables).
Results
Palynological analysis indicated the presence of 9 dinoflagellate genera, of which 6 belonged to the order Peridiniales, with 12 species, and 3 belonged to the order Gonyaulacales, with 4 species (Fig. 3). Basic statistics of palynological data (Table 2) revealed that in the 19th century the average of total concentrations of heterotrophic dinoflagellates was 138 cys/g sed and the average of autotrophic dinoflagellates was 49 cys/g sed.
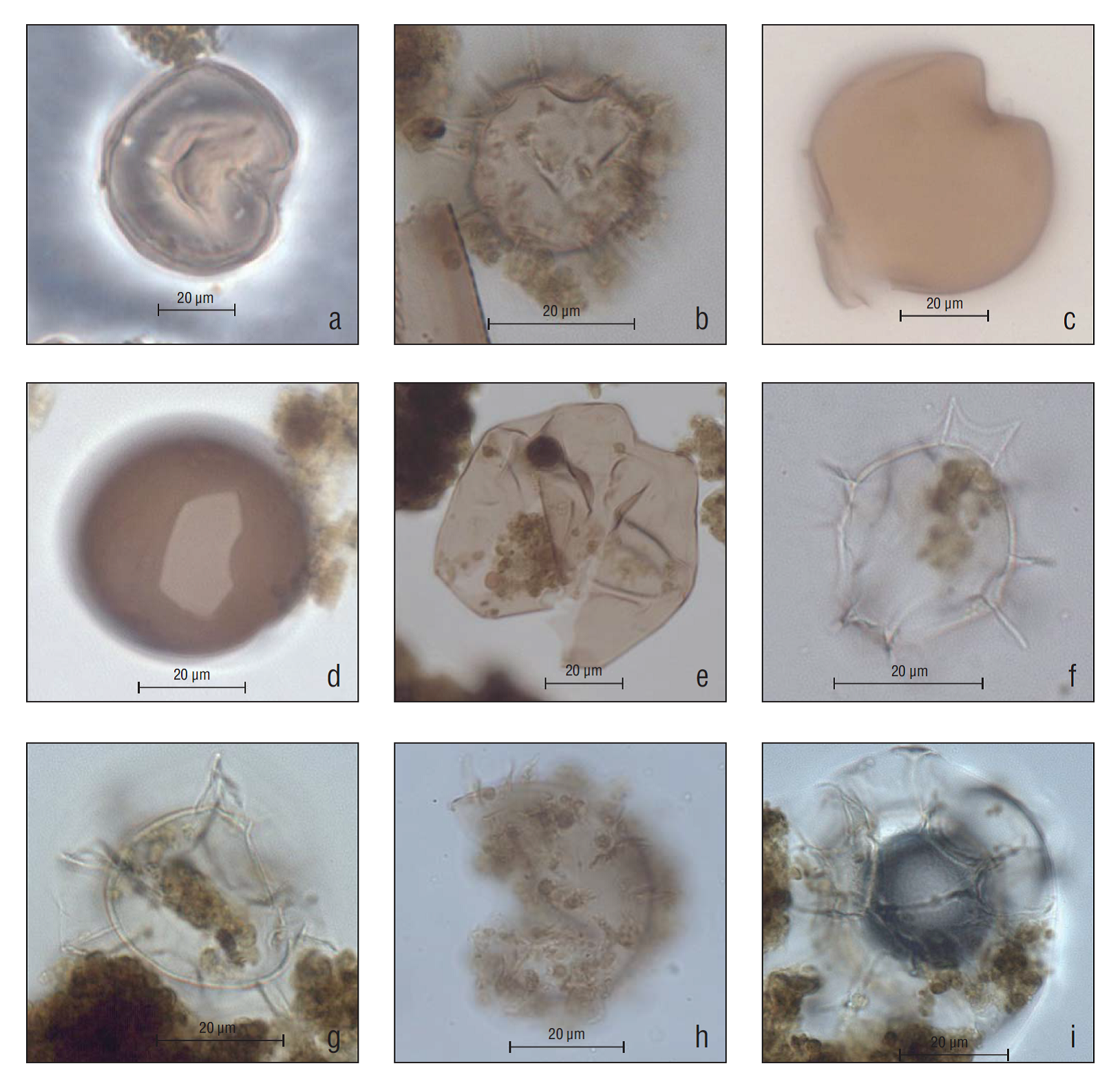
Figure 3 Photomicrographs of the most representative dinoflagellate species in the Magdalena Basin, Baja California Sur (Mexico). (a) Selenopemphix nephroides, lamina 59-60a, M26, high focus; (b) Echinidinium granulatum, lamina 60-61a, X33-1, high focus; (c) Quinquecuspis concreta, lamina 67-68a, F5-4, high focus; (d) Brigantedinium cariacoense, lamina 64-65a, U23-1, high focus; (e) Votadinium calvum, lamina 58-59a, X13, high focus; (f) Impagidinium aculeatum, lamina 65-66a, Z22-1, medium focus; (g) Impagidinium strialatum, lamina 88-89a, H58-3, medium focus; (h) Operculodinium israelianum, lamina 58-59a, W7-3, high focus; (i) Spiniferites sp., lamina 97-98a, C31, medium focus.
Absolute abundance
The concentrations of marine palynomorphs (dinoflagellate cysts and copepod eggs) fluctuated between 204 and 3,904 pal/g sed in the 1811-1907 CE time frame. An increase of 5,485 pal/g sed was detected in 1889 CE, and the highest concentrations were observed in 1901 CE (13,200 pal/g sed) and in 1905 CE (10,569 pal/g sed). Palynomorph concentrations significantly correlated (r = 0.45, P = 0.001) with SSTI values (Table 2), which concurs with the general increase in marine palynomorphs and SSTI in the 19th century. This correlation contrasts the temperature declines in 1837, 1857, and 1881 CE, when marine palynomorph concentrations increased. Conversely, when SSTI increased in 1853 and 1879 CE, marine palynomorph concentrations decreased (Fig. 4).
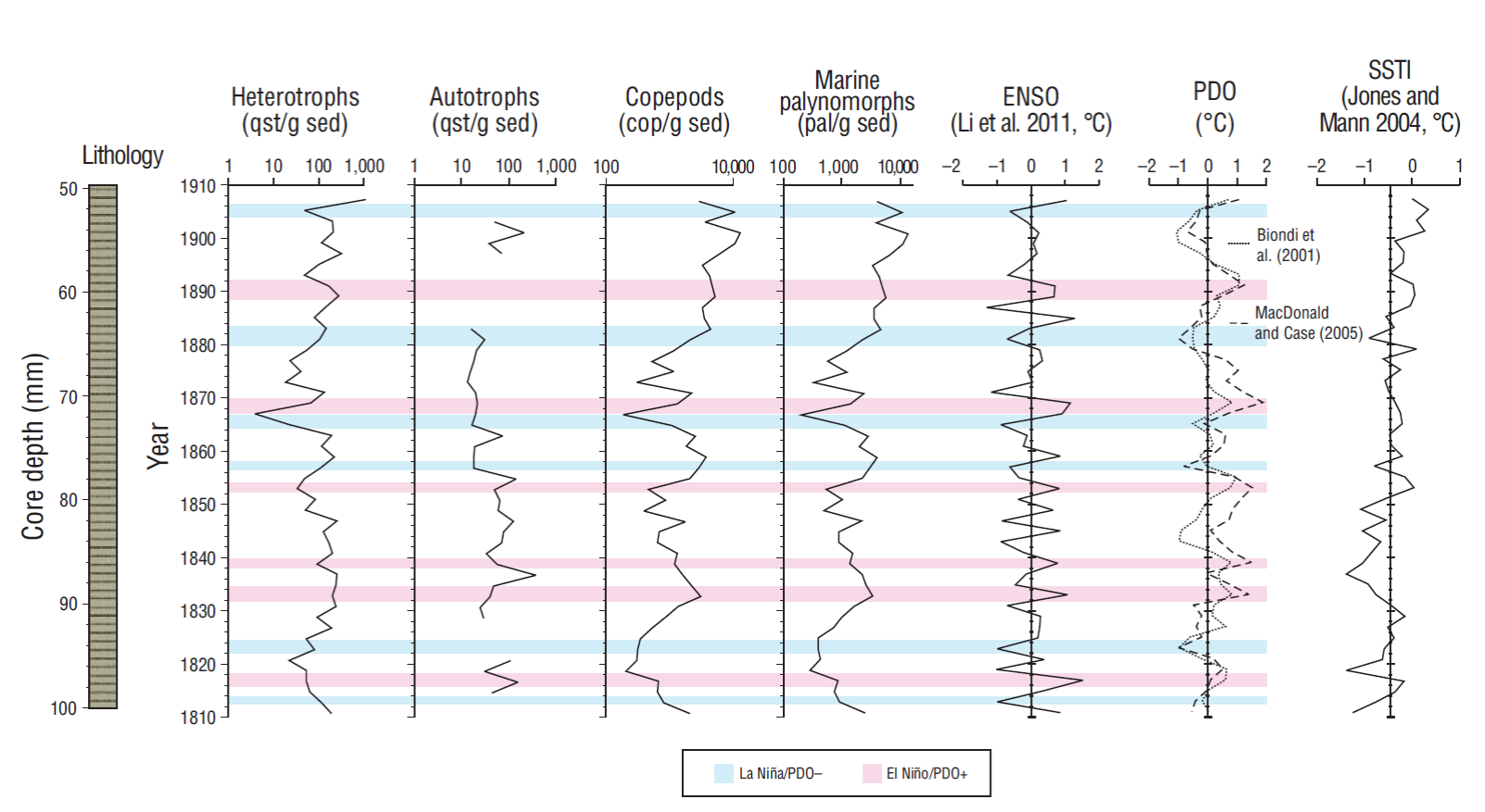
Figure 4 Comparison of concentrations of heterotrophic and autotrophic dinoflagellates (cys/g sed), copepods (cop/g sed), and total marine palynomorphs (pal /g sed) with the El Niño Southern Oscillation (ENSO) index reported by Li et al. (2011), the Pacific Decadal Oscillation Index (PDO) reported by Biondi et al. (2001) and MacDonald and Case (2005), and the sea surface temperature index (SSTI) reported by Jones and Mann (2004) in the period from 1811 to 1907 in Magdalena Basin, Baja California Sur (Mexico). The scale of the first 4 graphs is expressed as log10. The intervals where La Niña events overlapped the ODP-, and El Niño events overlapped the PDO+ are indicated with blue and pink shadings, respectively.
Dinoflagellate assemblages were divided by trophic strategies into heterotrophs and autotrophs. Heterotrophs were found in all samples and were the most important component in total dinoflagellate concentrations. Heterotroph concentrations varied from 17 to 308 cys/g sed in all samples. Heterotroph concentrations decreased during years with positive SSTI anomalies; however, these decreases were less marked than in 1867 CE. Heterotroph concentrations were generally high in the years with negative anomalies (e.g., 1837 CE) (Fig. 4). The correlation between heterotrophs and SSTI was not significant because it showed a value of r = 0.11 (P = 0.45) (Table 2).
In general, the concentrations of autotrophic dinoflagellates oscillated between 12 and 206 cys/g sed. No autotrophs were found for the years 1813, 1823-1827, 1885-1887, 1891, 1895, and 1905-1907 CE. Concentrations did not increase during years with positive SSTI, except in 1901 CE (Fig. 4). The correlation between autotrophic dinoflagellates and SSTI was significant and negative, with a value of r = -0.30 (P = 0.038) (Table 2).
Copepod concentrations were observed within a range of 202-5,960 cop/g sed, which coincided with negative SSTI values. Concentrations increased in 1899 CE (10,722 cop/g sed) and 1905 CE (10,521 cop/g sed), and these increases coincided with positive values of SSTI. Maximum concentrations occurred in 1901 CE with 12,787 cop/g sed (Fig. 4). The correlation with SSTI was significant and positive (r = 0.46, P = 0.001) (Table 2).
El Niño Southern Oscillation and the Pacific Decadal Oscillation
The intervals in which the polarities of ENSO and PDO values coincided are highlighted in Figure 4, that is, the cold zone of La Niña with the PDO- and the warm zone of El Niño with the PDO+. Taxa concentrations increased during 1881-1883 and in 1905 CE (La Niña, ODP-), whereas taxa concentrations decreased in 1817, 1853, and 1867 CE (El Niño, OPD+).
Multivariate analysis
The correlation coefficient (r i ) obtained with the canonical correlation analysis was statistically significant. A linear combination (r) of climate variables and palynological variables was inferred from these results, with a value of r i = 0.67 and P = 0.02. The canonical variate with r = 67% showed that copepods, the single heterotrophic dinoflagellate species, and the 2 autotrophic dinoflagellate species correlated significantly with the SSTI and ENSO.
Discussion
Primary productivity and sea surface temperature
In this study, copepods were identified only at the order level and were used as proxies for primary productivity because they are associated with phytoplankton. During the 19th century copepod concentrations were high in the Magdalena Basin, whereas dinoflagellate concentrations were low (Table 2). SSTI tended to be cooler during the 19th century, particularly from 1811 to 1879 CE, although it gradually increased during the period from 1881 to 1907 CE. Palynomorph concentrations mirrored climatic variability, since copepods and heterotrophic dinoflagellates decreased during 1811-1879 CE and increased during 1881-1907 CE. By contrast, the concentrations of autotrophic dinoflagellates increased during 1811-1879 CE and decreased during 1881-1907 CE. Cold conditions during the 19th century explain the absence of subtropical or tropical autotrophic species in the samples analyzed in the present study. Dinoflagellates that are typically tropical, such as Stelladinium stellatum (heterotroph), Bitectatodinium spongium (autotroph), or Polysphaeridium zoharyi (autotroph, Pyrodinium bahamense cyst), which were observed in this basin during 1907-2009 CE (Castañeda-Quezada 2016), were not observed during the present study.
Regarding productivity, high dinoflagellate productivity was to be expected because SSTI anomalies were predominantly negative from 1800 to 1900 CE. Nevertheless, productivity showed a strong increasing trend over the century as SST increased. This high productivity was directly correlated with heterotrophic dinoflagellates and copepods, and inversely correlated with autotrophic dinoflagellates.
The low productivity in the region in the 19th century has also been detected as low percentage values of biogenic opal (Juárez et al. 2014). This decline in diatom productivity implies that upwelling weakened and/or nutrient input decreased (Barron et al. 2013, Bringué et al. 2014, Juárez et al. 2014). The dominance of planktonic groups in the northeastern Pacific has been shown to alternate between calcareous and opaline forms, the latter being dominant during upwelling periods, when carbonate production is low (Juárez et al. 2015). Contrary to the apparent association between cold waters and high productivity, the changes in productivity observed here indicate that copepods and heterotrophic dinoflagellates proliferated during warm periods, when coastal winds intensified and coastal upwelling, as a result, increased (Rivas et al. 2016). Global warming can intensify coastal upwelling as land surface temperatures increase more than sea surface temperatures, strengthening the land-sea thermal contrast that modulates winds along the coast (Bakun 1990).
The higher productivity in the Magdalena Basin during the warm periods could be associated with the strengthening of oceanographic events on the southwestern coast of the Baja California Peninsula and to the concurrent effect on coastal upwelling, due to the influence of the North Equatorial Current, which mitigates seasonal changes in the coastal zone. This increasing strength coincides with the increase in the coastal upwelling index that was detected at 24ºN and 113ºW during 1900-2009 by the the National Oceanic and Atmospheric Administration Pacific Fisheries Environmental Laboratories (NOAA 2016). It also coincides with current global warming and with the higher productivity in the 2002-2014 CE period recorded for the region (Ortiz-Ahumada 2015).
Heterotrophic species such as those in the genus Brigantedinium are indicative of the continuous influence of coastal upwelling, and the presence of Echinidinium spp. is a strong sign of warmer winter SSTs and increasing primary productivity (Pospelova et al. 2015). Autotrophs, by contrast, proliferated during periods with less intense upwelling events, when stratification increased and competition with diatoms was more favorable.
The comparison of dinoflagellate concentrations in the Magdalena Basin between the 19th and 20th centuries (Table 2) shows changes in primary productivity, with a significant increase in productivity toward the 20th century. The average of total heterotrophic dinoflagellate concentrations was 824 cys/g sed in the 20th century, whereas the average of autotrophic dinoflagellates was 19 cys/g sed (Castañeda-Quezada 2016). This indicates that dinoflagellates behaved similarly for the past 2 centuries in relation to climate indices and current warming.
The correlation between dinoflagellate abundance and SSTI was low, probably because temperature effects are not linear given the influence of stratification (Bringué et al. 2014). Other influencing factors could have been the lack of correlation of the subsampling, the uncertainties of the chronological extrapolation, and the lack of a SST time series from a region closer to the study area.
Primary productivity, El Niño Southern Oscillation, and the Pacific Decadal Oscillation
No significant correlation was observed when palynomorph concentrations during 1811-1907 CE in the Magdalena Basin were compared with the ENSO and PDO indices. However, changes in some taxa concentrations are associated with interactions between these climatic forcings. In 1881-1883 and 1905 CE, variations in concentrations of copepods and heterotrophic dinoflagellates highlighted the signal of cold cycles (La Niña and PDO-). By contrast, changes in concentrations of the autotrophic genus Impagidinium coincided with the signal of warm cycles (El Niño and PDO+) in 1817, 1853, and 1867 CE. This discrepancy could have been caused by (i) uncertainties in the chronological extrapolation, (ii) geographic location of dendrochronology sampling sites, and (iii) a weaker signal of both reconstructions throughout the 19th century and the beginning of the 20th century (Bringué et al. 2014). In other words, no intense events were observed and the Magdalena Basin is, too, a transition zone of subarctic and subtropical waters.
These observations support the paradigm that most heterotrophic planktonic organisms considerably depend on diatom populations and respond to diatom blooms under turbulence conditions (ODP-) (Bringué et al. 2014). On the other hand, autotrophs tend to be more abundant during periods of moderate stratification (ODP+), but their concentrations decrease when the thermocline deeply shoals during strong El Niño events (Flores-Trujillo 2009).
In general, the richness and concentrations of dinoflagellates (

