











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Many tropical fish have adapted their reproductive strategies to the low environmental variability in their habitats and have opted for longer reproductive periods with multiple spawning events (Lowe-McConnell 1987). Tropical Pleuronectiformes species have shown long or nearly year-round reproductive periods (Gibson 2008).
The Achiridae family, also known as American soles, is one of the most abundant and least studied taxa of the order Pleuronectiformes. It includes 7 genera and 29 species (Robertson and Allen 2015). The sole Achirus mazatlanus (Steindachner, 1869), or Mexican sole, is a pelagic spawner that uses estuarine systems as habitats for nursing and development, and it is distributed from Baja California to Peru (Krupp 1995).
Achirus mazatlanus is the most abundant sole in the Barra de Navidad coastal lagoon (Jalisco, Mexico), where it reaches 22 cm in length (González-Sansón et al. 2014). The fact that this species is abundant, easy to catch, and continuously present in the system has arisen interest in the use of this species as a bioindicator of pollution in the lagoon (Aguilar-Betancourt et al. 2016). Despite its abundance and wide distribution range, there are few studies about this species, and it is only mentioned in systematic lists (e.g., Aguilar-Palomino et al. 2001, Lucano-Ramírez et al. 2001), in a study reporting its feeding habits and some information about its reproduction (Amezcua-Linares et al. 1992), and in studies reporting its taxonomic characteristics (Ortíz-Galindo et al. 1990, Gracian-Negrete and Del Moral-Flores 2013).
In view of all the above, and to partially fill in the glaring gaps in the information available for species, this study describes the reproductive characteristics of A. mazatlanus in the Barra de Navidad coastal lagoon, Jalisco, Mexico.
Materials and methods
Samples were obtained from the Barra de Navidad coastal lagoon, on the southern coast of Jalisco, Mexico (19º11′25″N, 104º39′53″W). This lagoon is permanently connected to Navidad Bay, allowing for port activities, tourism, and commerce. Moreover, the lagoon is part of the Laguna Barra de Navidad wetland, which consists of 382 ha of surface water, with seasonal inputs of fresh water from the Arroyo Seco and Marabasco rivers, and 620 ha of mangrove forest, the second largest mangrove forest in Jalisco (CONANP 2008).
Specimens were collected during weekly samplings from February 2011 to March 2012 and monthly samplings from April 2012 to August 2016. They were caught by one of 3 types of fishing gear: a beach seine net with a mesh size of 1 cm (5 casts at 2 sites, north beach and south beach, from 16:00 to 19:30), a cast net 3 m in diameter with a mesh size of 2.5 cm (40-50 casts at random sites from 16:00 to 19:30), and a trawl net measuring 5 m at the mouth with a mesh size of 2.5 cm (2 tows at 8 sites from 03:00 to 07:00). Each specimen was measured for total length (TL, ±0.1 cm) and total weight (TW, ±0.1 g). The gonads were extracted and weighed (±0.01 g), and sex was determined. Gonads were assigned a maturity stage following Everson et al. (1989). All gonads were fixed in neutral formalin.
For histological analyses, we processed a sample from each ovary (n = 96) but processed whole testes (n = 121). Samples were dehydrated, embedded in paraffin (Paraplast, Leica; Richmond, USA), sliced (6 µm-thick sections), stained with haematoxylin-eosin, and preserved in Canada balsam.
Ten sections per maturity stage were chosen for every month. We measured 10 oocytes (with visible nucleus) from every phase present in these sections; in addition, we measured ovarian wall thickness at 3 random points using a digital camera (AxioCam ERc5s, Zeiss; Oberkochen, Germany) mounted on a microscope (Axiostar, Zeiss; Oberkochen, Germany). Oocytes were classified according to Yamamoto and Yamazaki (1961) and Brown-Peterson et al. (2011), and testes were classified according to García-López et al. (2005).
A typical year was determined. We determined sex ratio by month and by length class using the total number of specimens. The chi squared (χ2) test was used to determine significant differences from the expected 1:1 ratio. We determined the gonadosomatic index (GSI; GSI = [gonadal weight/TW] × 100), relative condition factor (CF; CF = [TW/TL3.2] × 100), and average length at sexual maturity (L50), which was estimated by performing a nonlinear fit to the PTL = 1/[1+e (bTL + a) ] × 100 logistic model, where PTL is the percentage of mature individuals at TL, and a and b are the parameters of the fitted curve.
Student’s t test was used to determine significant differences between lengths of females and males. The one-way analysis of variance (ANOVA) was used to examine differences between monthly values. When significant values were obtained, a post hoc Student-Newman-Keuls (SNK) test was run to determine homogenous groups. Spearman’s correlation (r s ) was carried out. All statistical analyses were performed using a significance level of 0.05 and STATISTICA v.7.1 (StatSoft 2006).
Results
Size composition and sex ratio
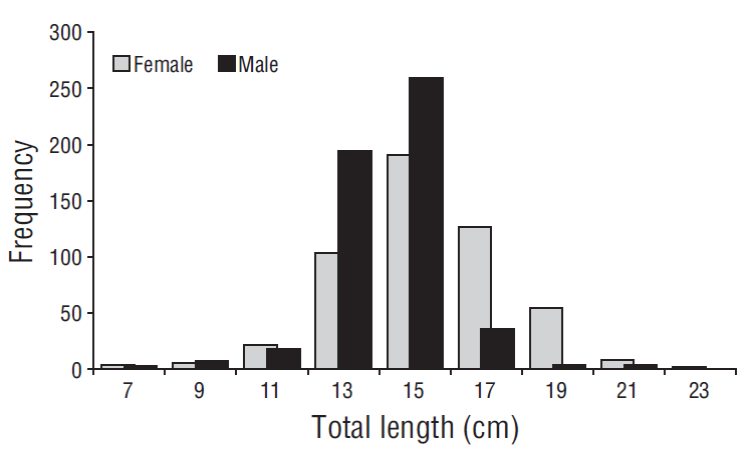
A total of 1,321 specimens of A. mazatlanus were analyzed. Average TL was 14.30 cm (±0.08), with a minimum of 1.4 cm and a maximum of 23 cm. We recorded 518 females (average TL = 15.39 cm, ±0.10), 531 males (average TL = 14.14 cm, ±0.07), and 272 specimens with unidentified sex. Females were larger than males (t 1, 1,048 = 10.04, P = 0.031). The smallest female and male measured 6.1 and 6.0 cm, respectively. Males showed higher frequencies in 2 length classes (13 and 15 cm), whereas females showed higher frequencies in 4 length classes (11, 17, 19, and 21 cm) (Fig. 1). The sex ratio (F:M) showed no differences for the total sample (1.00:1.03) (χ2 = 0.16, P = 0.68); however, the ratio was significantly biased toward males in February (1.0:1.7), July (1.0:1.4), and August (1.0:2.0) and significantly biased toward females in September (1.0:0.2). In addition, the ratio was significantly different among size classes; males were dominant in the 13 cm (1.0:2.2) and 15 cm (1.0:1.3) length classes, whereas females were dominant in the 17 cm (1.0:0.3), 19 cm (1.0:0.1), and 21 cm (1.0:0.3) length classes.
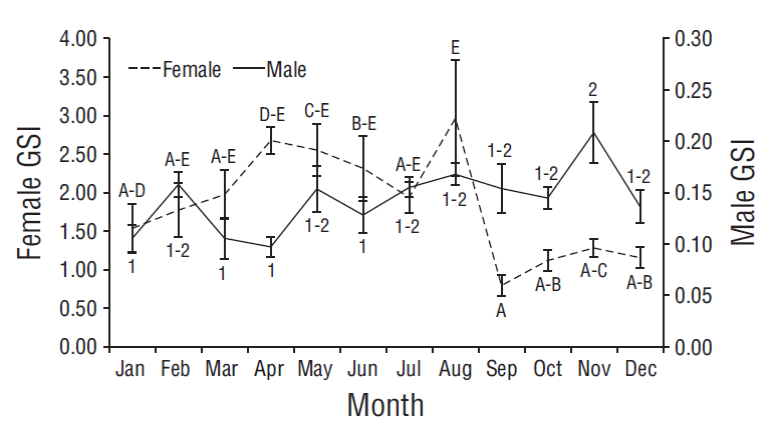
Monthly variation in the gonadosomatic index
Significant differences were observed in the monthly average values of female GSI (F 11, 380 = 3.82, P = 0.021) and male GSI (F 11, 380 = 3.82, P = 0.021) (Fig. 2). In the case of females, paired comparisons showed 5 overlapping groups. August showed the maximum value (2.96 ± 0.75, letter E in Fig. 2) and September the minimum value (0.79 ± 0.13, letter A in Fig. 2); the highest GSI values occurred between April (2.67 ± 0.18) and August. In the case of males, paired comparisons showed 2 overlapping groups. January (0.11 ± 0.01), March (0.11 ± 0.02), April (0.10 ± 0.01), and June (0.13 ± 0.02) showed minimum values (number 1 in Fig. 2), and November showed the maximum value (0.21 ± 0.02, number 2 in Fig. 2).
Monthly variation in the condition factor
Significant differences were observed in the monthly average CF values for females (F 11, 501 = 9.17, P = 0.001) and males (F 11, 504 = 55.7, P = 0.005) (Fig. 3). Paired comparisons showed 5 groups for both females and males, with an overlap in both cases. The maximum average values occurred in May for females (1.14 ± 0.03, letter E in Fig. 3) and males (1.09 ± 0.04, number 5 in Fig. 3); the minimum average values occurred in August (0.98 ± 0.01, letter A in Fig. 3) for females and in January (0.98 ± 0.01, number 1 in Fig. 3) and March (0.98 ± 0.02, number 1 in Fig. 3) for males. The CF of males and females showed a significant correlation (r s = 0.75, P = 0.005, n = 12). No correlation was observed between CF and GSI for males (r s = 0.27, P = 0.396, n = 12) or for females (r s = -0.18, P = 0.575, n = 12).

Gonad description
Gonads were found in the ventral cavity. Ovaries, which were located between the muscle and anal fin bones, formed long paired structures with smooth to granular surfaces. They extended to the middle of the anal fin in the more advanced maturity stages. Immature ovaries were pale in color, small, and flaccid. As they matured, they became larger and firmer, and changed from a yellow to a reddish color. Testes were small, round in shape, and reddish to yellowish-white in color. Turgidity increased with maturity.
Oocyte differentiation in the ovaries occurred in all developing phases. At first, the ovary showed only primary growth oocytes (Fig. 4a). These oocytes matured at different times; that is, oocyte development in the ovaries was asynchronous (Fig. 4b) until reaching advanced maturation phases, which included oocytes with germinal vesicle migration, oocytes with germinal vesicle breakdown, or hydrated oocytes (Fig. 4c, d). Testes showed spermatogenic development throughout the tissue (Fig. 5a). Spermatogonia originated in the cortex and continued to develop in the lobules as testes matured (Fig. 5b), which is typical in unrestricted lobular testes development (Fig. 5c). Nearly 50% of the 96 ovaries and 121 testes analyzed showed melanomacrophage centers (Fig. 5d).
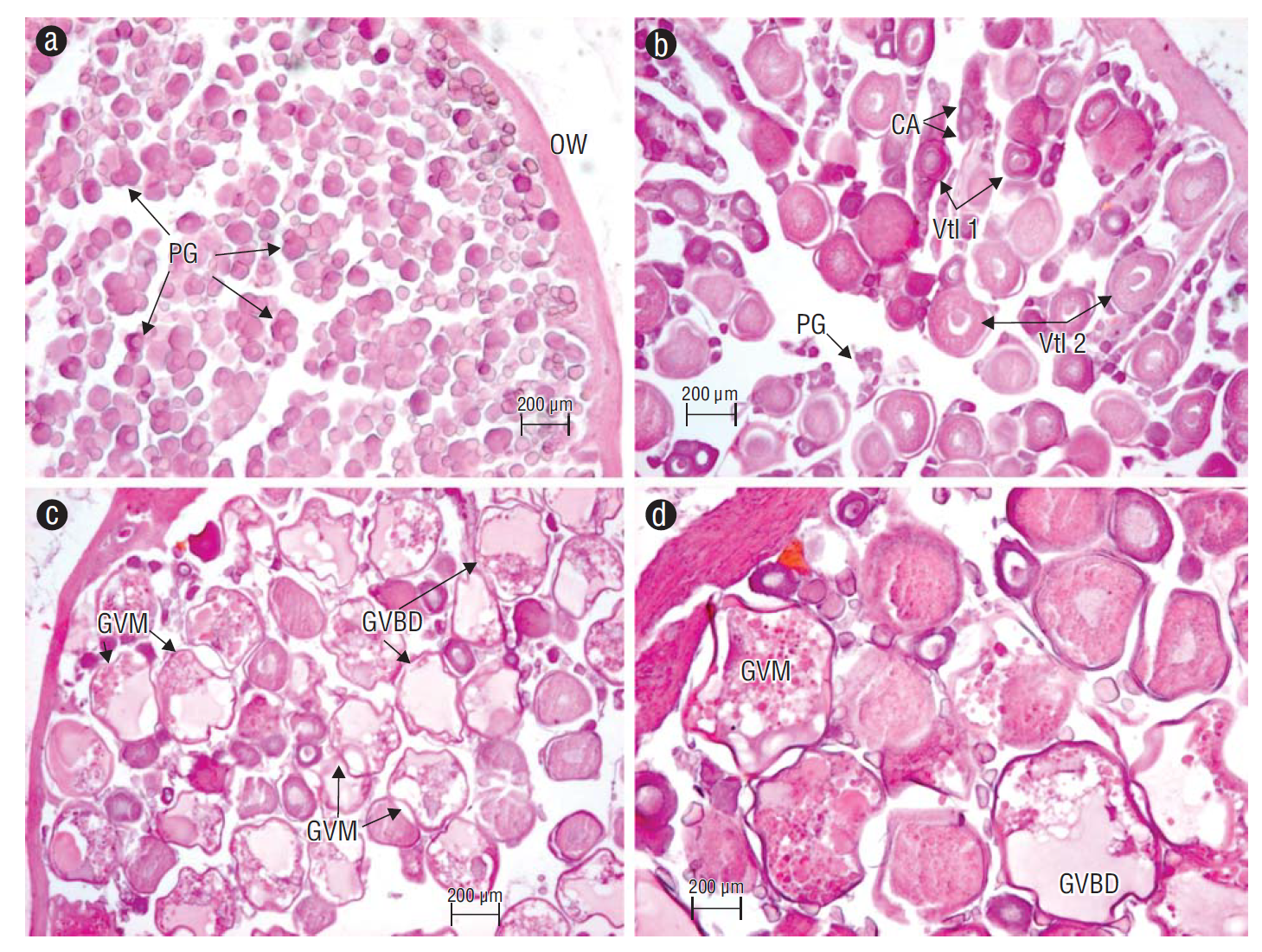
Figure 4 Achirus mazatlanus ovary with asynchronous oocyte development. (a) Oocytes in the primary growth (PG) phase and the ovarian wall (OW). (b) Oocytes during different developmental stages: oocytes in PG, cortical alveolus (CA), primary vitellogenesis (Vtl 1), and secondary vitellogenesis (Vtl 2). (c, d) Oocytes undergoing late germinal vesicle migration (GVM) and germinal vesicle breakdown (GVBD).

Gonad maturity stages
Females showed all 4 maturity stages throughout the year (except for immature females in December) (Fig. 6a). The highest percentages of mature females were found in March and April. In general, reproductive activity was highest from January to April and from June to August. Males also showed all 4 maturity stages, with the highest percentages of mature males occurring in February, March, and June-August (Fig. 6b).
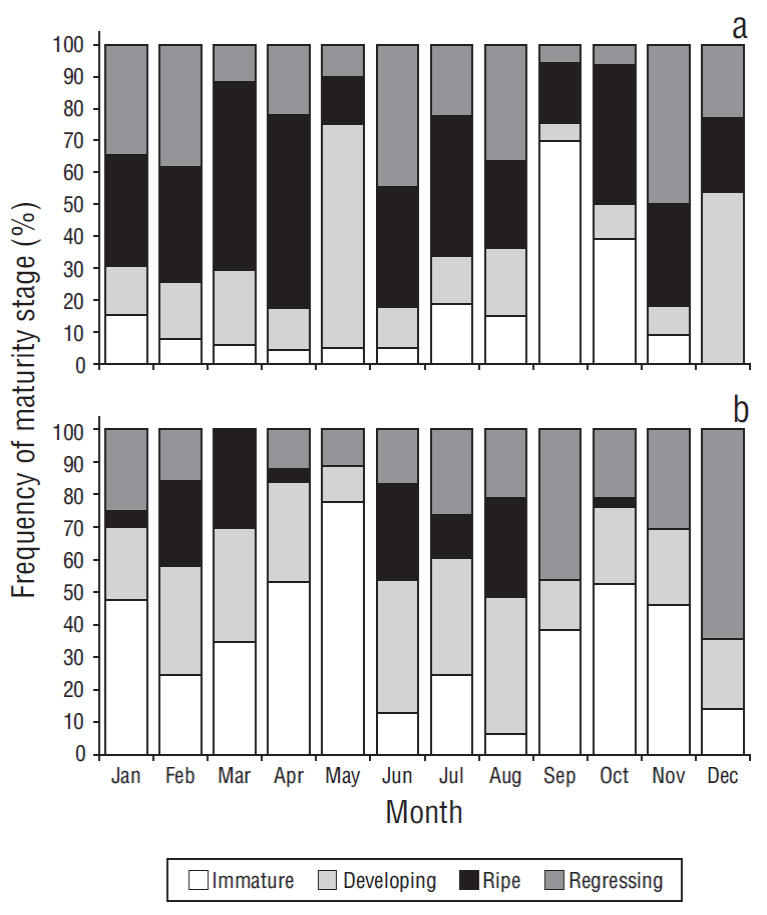
Figure 6 Frequency of the 4 stages of gonad development in female (a) and male (b) Achirus mazatlanus in the Barra de Navidad coastal lagoon, Jalisco, Mexico.
Oocyte diameter and ovarian wall thickness
ANOVA showed significant differences in monthly oocyte diameters (F 11, 941 = 6.43, P < 0.001). Paired comparisons showed 2 groups. The smallest diameters were recorded in February and December, and the highest averages were recorded in April, June, and August-October; all the other months showed an overlap between both groups (Fig. 7). No correlation was observed between oocyte diameter and GSI values (r s = 0.09, P = 0.762, n = 12).
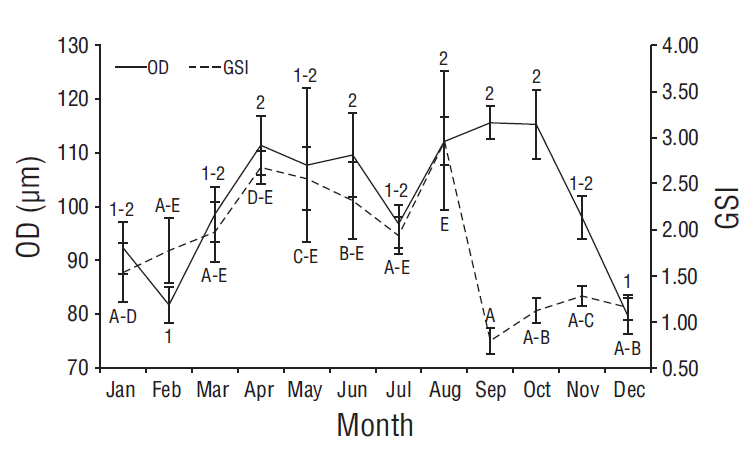
Figure 7 Monthly variation of oocyte diameter (OD) and the gonadosomatic index (GSI) for Achirus mazatlanus in the Barra de Navidad coastal lagoon, Jalisco, Mexico.
Ovarian wall thickness also exhibited differences throughout the year (F 11, 462 = 7.67, P < 0.001). The SNK test showed 3 thickness groups. Maximum values with no overlap were observed in November and December, and minimum values were observed in January, April-June, August, and September; all the other months showed an overlap between groups. A negative correlation was observed between ovarian wall thickness and GSI values (r s = -0.615, P = 0.033, n = 12).
Length at sexual maturity
The smallest mature female was 7.1 cm long, and the smallest mature male was 9.4 cm long. According to the nonlinear regression model, L50 was 11.26 and 10.23 cm for females and males, respectively (Fig. 8). More than 90% of females and males measuring 17 cm in length had already reproduced at least once in their lifetime.

Discussion
Gibson (2008) reported that sexual dimorphism is common in Pleuronectiformes. In this study, A. mazatlanus females were larger than males, a finding also reported by Gibson (2008) and Aguilar-Betancourt et al. (2016). Sexual dimorphism in this species could be the result of males spending more energy on early development and maturation, which is frequent in soles; the larger sizes in females, on the other hand, is related to high fecundity (Roff 1982). Recent studies on the life history of Pleuronectiformes suggest that males can invest little energy in testes maturation, and as a result testes gain less biomass (e.g., Rijnsdorp and Witthames 2005).
The occurrence of sex ratio differences in 4 months of the year could be due to changes in habitat use during the reproductive and resting periods. This behavior was reported by Amezcua-Linares et al. (1992) for A. mazatlanus and is apparently common in Pleuronectiformes (Rijnsdorp and Witthames 2005). However, sex ratio deviated from the 1:1 ratio during periods with less reproductive activity, and since specimens were caught throughout the lagoon, we cannot fully determine if differences in the monthly sex ratios were caused by reproductive factors, physiological factors, or changes in habitat use (Gibson 2008).
The period with the highest reproductive activity occurred during the dry season, in the spring-summer months, a finding that concurs with other Pleuronectiformes studies (Amezcua-Linares et al. 1992, De Oliveira and Fávaro 2010, De Oliveira and Favaro 2011). Kamler (2002) reported that rain and changes in salinity can disrupt egg and larvae development, triggering resting periods or reproductive arrest. In the present study, male A. mazatlanus showed low GSI values because testes biomass was low. This was an expected result because males from many Pleuronectiformes species exhibit this characteristic (García-López et al. 2005, Rijnsdorp and Witthames 2005, García-López et al. 2006). Since male GSI showed little variability, we determined the reproductive period for A. mazatlanus from our observations on female characteristics. This approach is similar to that used by Juntti and Fernald (2016), who used ovarian characteristics to determine the reproductive period.
Some sole species do not significantly change their body condition to improve gonad or, by extension, egg quality (Roff 1982). This behavior could explain the lack of correlation between CF and GSI in the present study. Furthermore, the body condition of soles in estuarine systems depends on changes in benthic communities (Girardin et al. 2016), and though the rainy season in the area might not influence the condition of A. mazatlanus, it does have an effect on its reproductive period (Amezcua-Linares et al. 1992, De Oliveira and Fávaro 2010).
Mature ovaries and hydrated oocytes were observed in months with low GSI values, indicating that females are at times allocating little energy to gonad development (McBride et al. 2013). Mature and spawned/ejaculated gonads were observed throughout the year. This finding is consistent with the bet-hedging theory and, in turn, explains the dominance of soles in this habitat, given the adaptive capacity conferred by this strategy (Simons 2011). This finding could also suggest that, although the species is categorized as a pelagic spawner, it is actually spawning inside the lagoon. Spawning in lagoons has already been recorded in other estuarine systems (Ortíz-Galindo et al. 1990, Silva-Falcão et al. 2012) but would have to be confirmed for the Barra de Navidad Lagoon.
Though high GSI values and periods with maximum oocyte diameters were observed throughout the typical year, paired comparisons did not indicate a defined reproductive period. Nonetheless, in the present study we used other indicators of reproductive activity, such as ovarian wall thickness; this indicator is directly associated with the space occupied by the gonad, since ovarian wall thickness decreases as the muscle stretches with oocyte growth (Behera et al. 2015). Ovarian wall thickness clearly decreased during the months when mature gonads were observed and increased during the resting and spawning seasons. There was a negative correlation between ovarian wall thickness and GSI.
The high GSI values, oocyte diameter, presence of mature gonads, and reduced ovarian wall thickness due to oocyte maturation could all together indicate that, although A. mazatlanus can reproduce throughout most of the year, it exhibits a period with high reproductive activity from April to August.
This study reports L50 values for A. mazatlanus for the first time. In a previous study, Amezcua-Linares et al. (1992) reported that maturity of A. mazatlanus in the Agua Brava Lagoon, north of Nayarit, Mexico, occurs when individuals reach 14.0-15.9 cm in length (no specific value was provided). The differences between the values reported by both studies could be attributed to the sampling site, sampling years, sample size, and different fishing gears used in the present study, which allowed sampling for a wider length range. Though size at sexual maturity in Pleuronectiformes tends to be similar for both sexes, males tend to mature at slightly shorter lengths than females (Rijnsdorp and Witthames 2005, De Oliveira and Favaro 2011), and this is in agreement with the results reported in the present study.
We did not find any studies describing the macroscopic and microscopic characteristics of A. mazatlanus gonads. Nonetheless, the ovarian characteristics observed for this species during this study coincide with those described for females of other Pleuronectiformes species (Rijnsdorp and Witthames 2005). Achirus mazatlanus gonads exhibited melanomacrophage centers but these centers showed no correlation with the length, time, sex, and gonad maturity variables. Some authors have associated melanomacrophage centers with pathological elements and environmental pollution (Agius and Roberts 2003). Aguilar-Betancourt et al. (2016) analyzed concentrations of stable carbon and nitrogen isotopes in A. mazatlanus tissues and found moderate levels of pollution. It is not clear if these pollution levels are the culprits in the presence of melanomacrophage centers in gonads; we therefore recommend carrying out studies that specifically address this aspect for this species.
The paucity of information on A. mazatlanus stems from the fact that this species has no commercial value. However, because of its generalist habits, abundance, wide distribution, and presence in estuaries and bays, this species is ecologically important. We should not wait for commercial species to collapse to begin studying non-commercial species because habitat degradation is ongoing and can be especially harmful on species whose basic biology and habitat preferences are unknown (Damalas et al. 2010).

